
Effects of Different Soils on the Biomass and Photosynthesis of Rumex nepalensis in Subalpine Region of Southwestern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Seed Collection
2.2. Greenhouse Experiment
2.3. Photosynthesis and Chlorophyll Measurements
2.4. Plant and Soil Sampling
2.5. Soil Property Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Properties
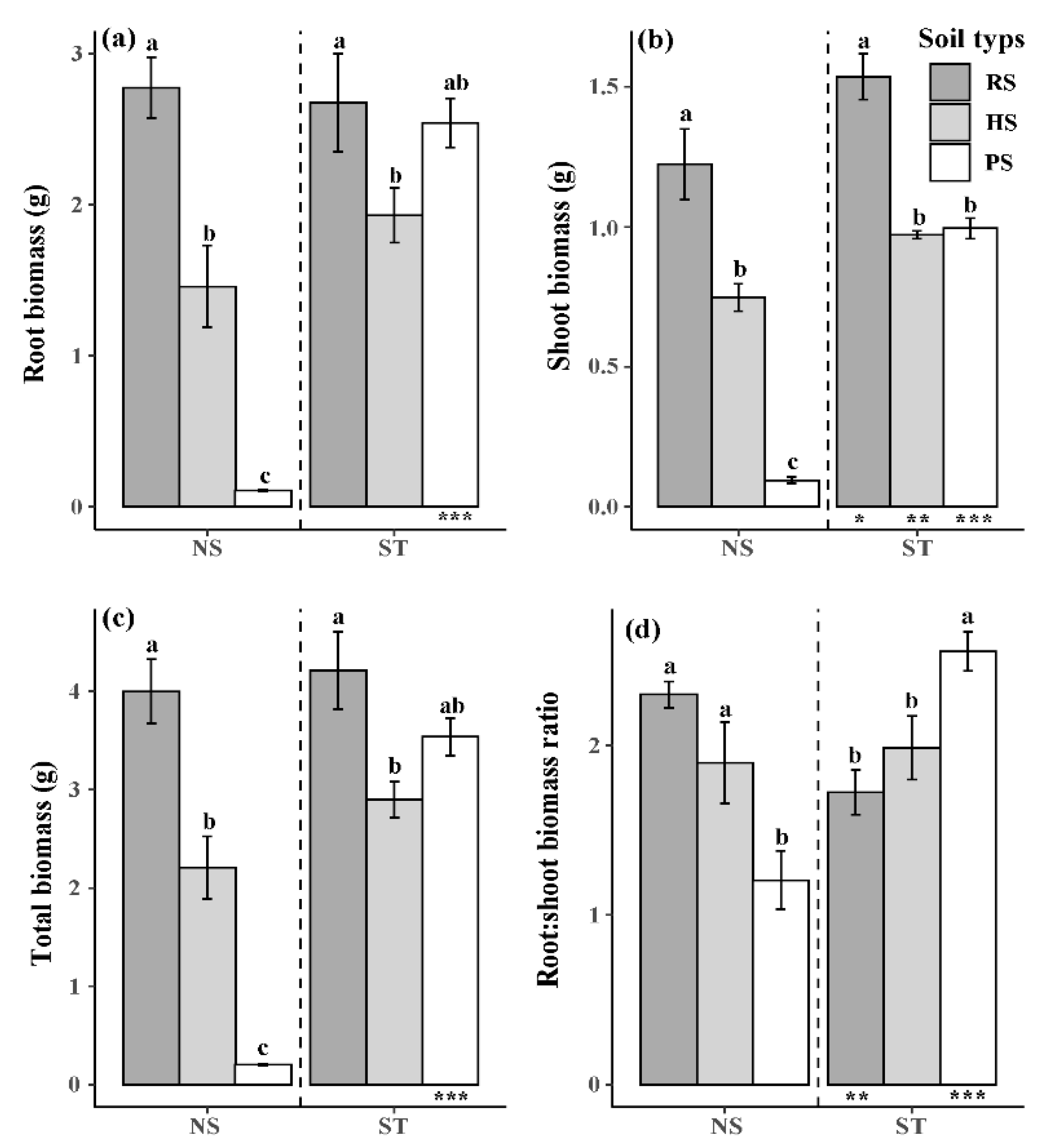
3.2. Biomass and Biomass Allocation
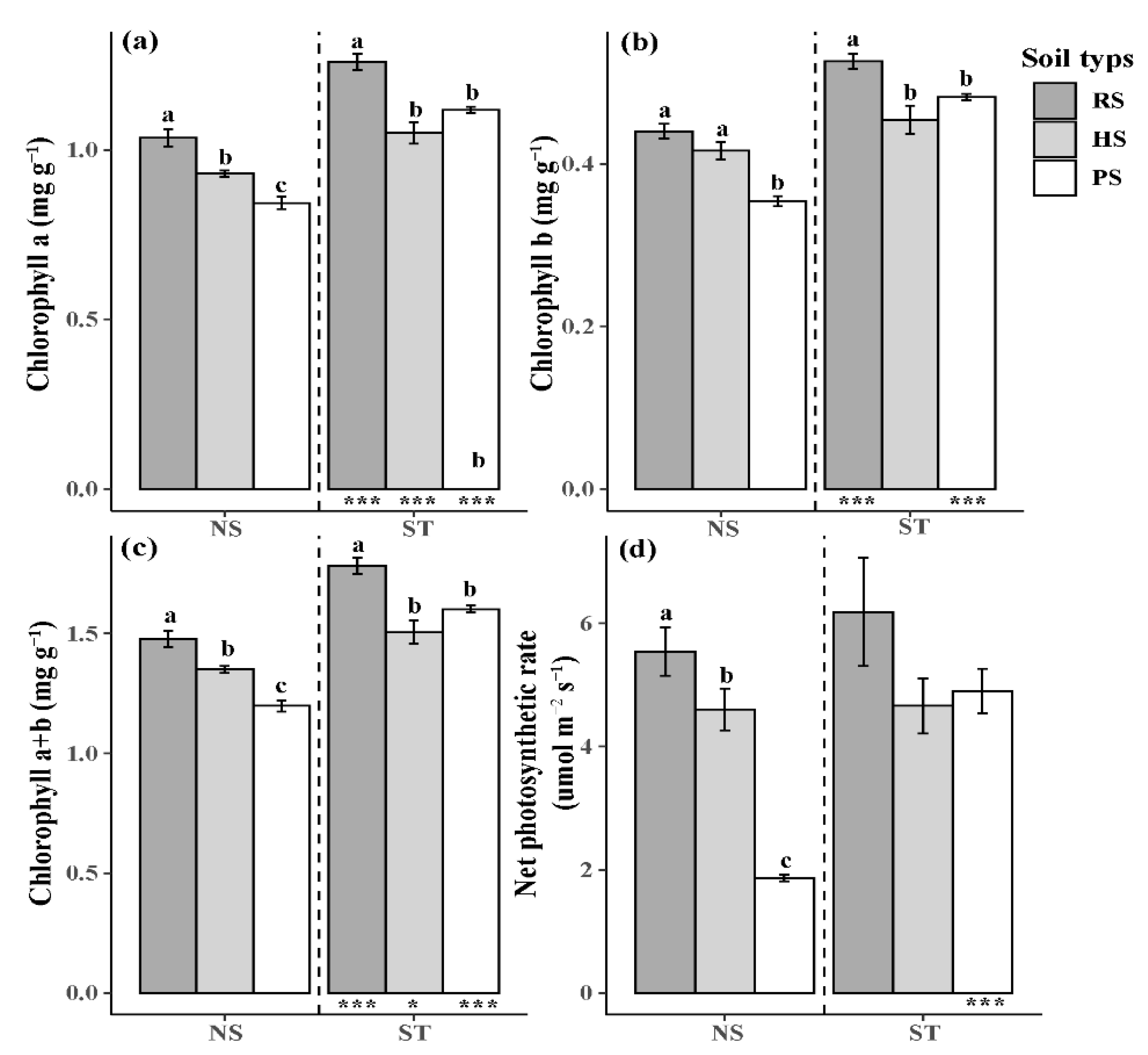
3.3. Photosynthesis and Chlorophyll
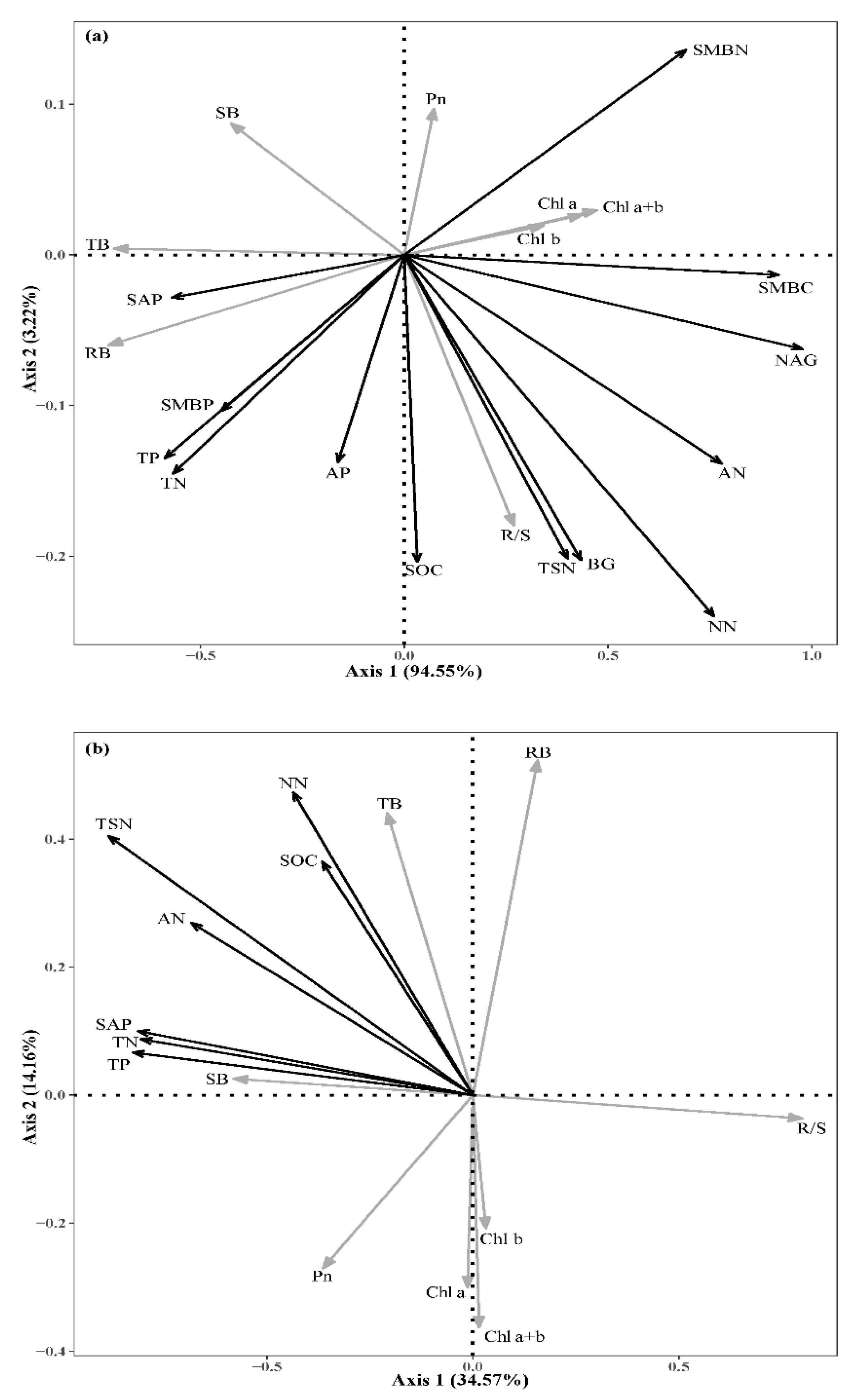
3.4. Effects of Soil Factors on Plant Performance
4. Discussion
4.1. Feedback Effects of Different Soils on Plant Biomass and Physiology
4.2. Important Soil Parameters Affecting Plant Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bever, J.D. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant–soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Yan, Y.; Dai, Q.; Hu, G.; Jiao, Q.; Mei, L.; Fu, W. Effects of vegetation type on the microbial characteristics of the fissure soil-plant systems in karst rocky desertification regions of SW China. Sci. Total Environ. 2020, 712, 136543. [Google Scholar] [CrossRef]
- Carteron, A.; Parasquive, V.; Blanchard, F.; Guilbeault-Mayers, X.; Turner, B.L.; Vellend, M.; Laliberte, E. Soil abiotic and biotic properties constrain the establishment of a dominant temperate tree into boreal forests. J. Ecol. 2020, 108, 931–944. [Google Scholar] [CrossRef]
- Zhao, J.; Li, S.; He, X.; Liu, L.; Wang, K. The soil biota composition along a progressive succession of secondary vegetation in a karst area. PLoS ONE 2014, 9, 0112436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, P.M.S.; Schaefer, C.E.G.A.R.; Silva, J.D.O.; Ferreira Junior, W.G.; Dos Santos, R.M.; Neri, A.V. The influence of soil on vegetation structure and plant diversity in different tropical savannic and forest habitats. J. Plant. Ecol. 2016, 11, 226–236. [Google Scholar] [CrossRef]
- Kutakova, E.; Herben, T.; Munzbergova, Z. Heterospecific plant–soil feedback and its relationship to plant traits, species relatedness, and co-occurrence in natural communities. Oecologia 2018, 187, 679–688. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Brooker, R.; Atwater, D.Z.; Maron, J.L.; Callaway, R.M. The mechanisms and consequences of interspecific competition among plants. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 263–281. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; DeLucia, E.H.; Moore, D.; Nowak, R.; Schlesinger, W.H. Competition and facilitation: Contrasting effects of Artemisia tridentata on desert vs montane pines. Ecology 1996, 77, 2130–2141. [Google Scholar] [CrossRef] [Green Version]
- Balicka, A.; Szadkowska, D. XRF analysis of heavy metals contents in oak wood (Quercur robur L.). Ann. Warsaw Univ. Life Sci. SGGW For. Wood Technol. 2020, 109, 76–80. [Google Scholar] [CrossRef]
- Niedbała, M. Selected techniques of soil contaminated phytoremediation with the use of hyperaccumulative plants and trees. Ann. Warsaw Univ. Life Sci.--SGGW For. Wood Technol. 2021, 115, 77–84. [Google Scholar] [CrossRef]
- Zielenkiewicz, T.; Szadkowski, J.; Drożdżek, M.; Zielenkiewicz, A.; Kłosińska, T.; Antczak, A.; Zawadzki, J.; Gawron, J. Application of X-ray fluorescence technique for determination of heavy metals uptake by different species of poplar. Drewno 2016, 59, 113–126. [Google Scholar]
- de la Pena, E.; Bonte, D. Soil biota effects on clonal growth and flowering in the forest herb Stachys sylvatica. Acta Oecol. 2011, 37, 110–116. [Google Scholar] [CrossRef]
- Bennett, J.A.; Klironomos, J. Mechanisms of plant–soil feedback: Interactions among biotic and abiotic drivers. New Phytol. 2019, 222, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Ramesh, L.M.; Reynolds, H.L.; Michalet, R. The next frontier of plant–soil feedback research: Unraveling context dependence across biotic and abiotic gradients. J. Veg. Sci. 2017, 28, 484–494. [Google Scholar] [CrossRef]
- Gross, N.; Kunstler, G.; Liancourt, P.; de Bello, F.; Suding, K.N.; Lavorel, S. Linking individual response to biotic interactions with community structure: A trait-based framework. Funct. Ecol. 2009, 23, 1167–1178. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Raaijmakers, C.E.; Van der Putten, W.H. Plant community development is affected by nutrients and soil biota. J. Ecol. 2004, 92, 824–834. [Google Scholar] [CrossRef]
- Heinze, J.; Gensch, S.; Weber, E.; Joshi, J. Soil temperature modifies effects of soil biota on plant growth. J. Plant. Ecol. 2017, 10, 808–821. [Google Scholar] [CrossRef] [Green Version]
- Bever, J.D.; Westover, K.M.; Antonovics, J. Incorporating the soil community into plant population dynamics: The utility of the feedback approach. J. Ecol. 1997, 85, 561–573. [Google Scholar] [CrossRef]
- Kutakova, E.; Meszarosova, L.; Baldrian, P.; Munzbergova, Z. Evaluating the role of biotic and chemical components of plant–soil feedback of primary successional plants. Biol. Fert. Soils 2020, 56, 345–358. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X.L. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, F.R.; Fernandez, I.J.; Simon, K.S.; Dail, D.B. Nitrogen and phosphorus regulation of soil enzyme activities in acid forest soils. Soil Biol. Biochem. 2016, 98, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Raiesi, F.; Salek-Gilani, S. The potential activity of soil extracellular enzymes as an indicator for ecological restoration of rangeland soils after agricultural abandonment. Appl. Soil Ecol. 2018, 126, 140–147. [Google Scholar] [CrossRef]
- Zhang, C.; Song, Z.L.; Zhuang, D.H.; Wang, J.; Xie, S.S.; Liu, G.B. Urea fertilization decreases soil bacterial diversity, but improves microbial biomass, respiration, and N-cycling potential in a semiarid grassland. Biol. Fert. Soils 2019, 55, 229–242. [Google Scholar] [CrossRef]
- Castle, S.C.; Lekberg, Y.; Affleck, D.; Cleveland, C.C. Soil abiotic and biotic controls on plant performance during primary succession in a glacial landscape. J. Ecol. 2016, 104, 1555–1565. [Google Scholar] [CrossRef]
- Shipley, B. Net assimilation rate, specific leaf area and leaf mass ratio: Which is most closely correlated with relative growth rate? A meta-analysis. Funct. Ecol. 2006, 20, 565–574. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Jahan, M.A.; Kovinich, N. Acidity stress for the systemic elicitation of glyceollin phytoalexins in soybean plants. Plant Signal Behav. 2019, 14, 1604018. [Google Scholar] [CrossRef]
- Luo, X.F.; Dai, Y.J.; Zheng, C.; Yang, Y.Z.; Chen, W.; Wang, Q.C.; Chandrasekaran, U.; Du, J.; Liu, W.G.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Phys. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Horn, R.; Grundmann, G.; Paulsen, H. Consecutive binding of chlorophylls a and b during the assembly in vitro of light-harvesting chlorophyll-a/b protein (LHCIIb). J. Mol. Biol. 2007, 366, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, K.J.F.; Biere, A.; Nevo, E.; Damme, J. Differential selection of growth rate-related traits in wild barley, Hordeum spontaneum, in contrasting greenhouse nutrient environments. J. Evol. Biol. 2004, 17, 184–196. [Google Scholar] [CrossRef] [Green Version]
- Bresson, J.; Vasseur, F.; Dauzat, M.; Koch, G.; Granier, C.; Vile, D. Quantifying spatial heterogeneity of chlorophyll fluorescence during plant growth and in response to water stress. Plant Methods 2015, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marbà, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, P.M. Tolerance of Mycorrhiza infected Pistachio (Pistacia vera L.) seedling to drought stress under glasshouse conditions. J. Plant Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Kostenko, O.; Bezemer, T.M. Abiotic and biotic soil legacy effects of plant diversity on plant performance. Front. Ecol. Evol. 2020, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Teste, F.P.; Kardol, P.; Turner, B.L.; Wardle, D.A.; Zemunik, G.; Renton, M.; Laliberte, E. Plant–soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science 2017, 355, 173–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.B.; Duan, B.L.; Xian, J.R.; Korpelainen, H.; Li, C.Y. Links between plant diversity, carbon stocks and environmental factors along a successional gradient in a subalpine coniferous forest in Southwest China. For. Ecol. Manag. 2011, 262, 361–369. [Google Scholar] [CrossRef]
- Shaikh, S.; Shriram, V.; Srivastav, A.; Barve, P.; Kumar, V. A critical review on Nepal Dock (Rumex nepalensis): A tropical herb with immense medicinal importance. Asian Pac. J. Trop. Med. 2018, 11, 405–414. [Google Scholar]
- Endlweber, K.; Scheu, S. Establishing arbuscular mycorrhiza-free soil: A comparison of six methods and their effects on nutrient mobilization. Appl. Soil Ecol. 2006, 34, 276–279. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N,N-dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [Green Version]
- Bing, H.; Wu, Y.; Zhou, J.; Sun, H.; Luo, J.; Wang, J.; Yu, D. Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau. J. Soils Sediments 2016, 16, 405–416. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Hooper, D.; Coughlan, J.; Mullen, M. Structural equation modelling: Guidelines for determining model fit. Electron J. Bus. Res. Methods 2008, 6, 53–60. [Google Scholar]
- Team, R.C. R: A language and Enviroment for Stadistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- van der Putten, W.H.; van Dijk, C.; Troelstra, S.R. Biotic soil factors affecting the growth and development of Ammophila arenaria. Oecologia 1988, 76, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Cardinaux, A.; Hart, S.P.; Alexander, J.M. Do soil biota influence the outcome of novel interactions between plant competitors? J. Ecol. 2018, 106, 1853–1863. [Google Scholar] [CrossRef] [PubMed]
- Glynn, C.; Herms, D.A.; Egawa, M.; Hansen, R.; Mattson, W.J. Effects of nutrient availability on biomass allocation as well as constitutive and rapid induced herbivore resistance in poplar. Oikos 2003, 101, 385–397. [Google Scholar] [CrossRef]
- Johnson, N.C.; Rowland, D.L.; Corkidi, L.; Allen, E.B. Plant winners and losers during grassland N-eutrophication differ in biomass allocation and mycorrhizas. Ecology 2008, 89, 2868–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Ai, Z.M.; Zhang, J.Y.; Liu, H.F.; Xin, Q.; Xue, S.; Liu, G.B. Soil nutrients influence the photosynthesis and biomass in invasive Panicum virgatum on the Loess Plateau in China. Plant Soil 2017, 418, 153–164. [Google Scholar] [CrossRef]
- Qi, Y.; Wei, W.; Chen, C.; Chen, L. Plant root-shoot biomass allocation over diverse biomes: A global synthesis. Glob. Ecol. Conserv. 2019, 18, 00606. [Google Scholar] [CrossRef]
- Franco, A.L.C.; Gherardi, L.A.; de Tomasel, C.M.; Andriuzzi, W.S.; Ankrom, K.E.; Bach, E.M.; Guan, P.T.; Sala, O.E.; Wall, D.H. Root herbivory controls the effects of water availability on the partitioning between above- and below-ground grass biomass. Funct. Ecol. 2020, 34, 2403–2410. [Google Scholar] [CrossRef]
- Erb, M.; Lu, J. Soil abiotic factors influence interactions between belowground herbivores and plant roots. J. Exp. Bot. 2013, 64, 1295–1303. [Google Scholar] [CrossRef]
- Tsunoda, T.; Kachi, N.; Suzuki, J.-I. Effects of belowground vertical distribution of a herbivore on plant biomass and survival in Lolium perenne. Ecol. Res. 2014, 29, 351–355. [Google Scholar] [CrossRef]
- Bever, J.D.; Mangan, S.A.; Alexander, H.M. Maintenance of Plant Species Diversity by Pathogens. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 305–325. [Google Scholar] [CrossRef]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J. Plant. Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef]
- Bussotti, F.; Pollastrini, M.; Holland, V.; Bruggemann, W. Functional traits and adaptive capacity of European forests to climate change. Environ. Exp. Bot. 2015, 111, 91–113. [Google Scholar] [CrossRef]
- Nowicka, B.; Ciura, J.; Szymanska, R.; Kruk, J. Improving photosynthesis, plant productivity and abiotic stress tolerance—Current trends and future perspectives. J. Plant Physiol. 2018, 231, 415–433. [Google Scholar] [CrossRef]
- Malhi, Y.; Doughty, C.E.; Goldsmith, G.R.; Metcalfe, D.B.; Girardin, C.A.J.; Marthews, T.R.; del Aguila-Pasquel, J.; Aragao, L.E.O.C.; Araujo-Murakami, A.; Brando, P.; et al. The linkages between photosynthesis, productivity, growth and biomass in lowland Amazonian forests. Glob. Chang. Biol. 2015, 21, 2283–2295. [Google Scholar] [CrossRef]
- Onoda, Y.; Hikosaka, K.; Hirose, T. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Funct. Ecol. 2004, 18, 419–425. [Google Scholar] [CrossRef]
- Shangguan, Z.P.; Shao, M.G.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Xu, Z.; Wan, C.; Xiong, P.; Tang, Z.; Hu, R.; Cao, G.; Liu, Q. Initial responses of soil CO2 efflux and C, N pools to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China. Plant Soil 2010, 336, 183–195. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Q.; Huang, Y.; Deng, H.; Shen, X.; Tang, S.; Zhang, J.; Liu, Y. Response of soil respiration and microbial biomass carbon and nitrogen to nitrogen application in subalpine forests of western Sichuan. Acta Ecol. Sin. 2019, 39, 7197–7207. [Google Scholar]
- Nasholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Persson, J.; Hogberg, P.; Ekblad, A.; Hogberg, M.N.; Nordgren, A.; Nasholm, T. Nitrogen acquisition from inorganic and organic sources by boreal forest plants in the field. Oecologia 2003, 137, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.E.; Bowman, W.D.; Suding, K.N. Plant uptake of inorganic and organic nitrogen: Neighbor identity matters. Ecology 2007, 88, 1832–1840. [Google Scholar] [CrossRef]
- Stark, S.; Mannisto, M.K.; Eskelinen, A. Nutrient availability and pH jointly constrain microbial extracellular enzyme activities in nutrient-poor tundra soils. Plant Soil 2014, 383, 373–385. [Google Scholar] [CrossRef]
- Zhu, X.M.; Liu, M.; Kou, Y.P.; Liu, D.Y.; Liu, Q.; Zhang, Z.L.; Jiang, Z.; Yin, H.J. Differential effects of N addition on the stoichiometry of microbes and extracellular enzymes in the rhizosphere and bulk soils of an alpine shrubland. Plant Soil 2020, 449, 285–301. [Google Scholar] [CrossRef]
- Kaye, J.P.; Hart, S.C. Competition for nitrogen between plants and soil microorganisms. Trends Ecol. Evol. 1997, 12, 139–143. [Google Scholar] [CrossRef]
- Hodge, A.; Robinson, D.; Fitter, A. Are microorganisms more effective than plants at competing for nitrogen? Trends Plant Sci. 2000, 5, 304–308. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Streeter, T.C.; Bol, R. Soil microbes compete effectively with plants for organic-nitrogen inputs to temperate grasslands. Ecology 2003, 84, 1277–1287. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Unsterilized Soil | Sterilized Soil | ||||
|---|---|---|---|---|---|---|
| RS | HS | PS | RS | HS | PS | |
| SOC (g kg−1) | 38.30 ± 0.19 a | 25.67 ± 0.60 c | 35.23 ± 0.05 b | 37.94 ± 0.34 a | 24.38 ± 0.22 c | 34.95 ± 0.06 b |
| TN (g kg−1) | 3.70 ± 0.04 a | 2.80 ± 0.05 b | 2.59 ± 0.06 b | 3.76 ± 0.02 a | 2.83 ± 0.05 b | 2.62 ± 0.03 c |
| TP (g kg−1) | 1.018 ± 0.006 aA | 0.660 ± 0.007 b | 0.650± 0.005 bA | 0.988 ± 0.007 aB | 0.676 ± 0.007 b | 0.668 ± 0.004 bB |
| SAP (mg kg−1) | 14.35 ± 0.14 aA | 5.23 ± 0.14 bA | 3.02 ± 0.05 cA | 15.34 ± 0.41 aB | 5.66 ± 0.06 bB | 3.41 ± 0.09 cB |
| Soil Properties | Unsterilized Soil | Sterilized Soil | ||||
|---|---|---|---|---|---|---|
| RS | HS | PS | RS | HS | PS | |
| SOC (g kg−1) | 39.13 ± 0.45 aA | 26.8 ± 0.21 cA | 35.4 ± 0.02 bA | 36.94 ± 0.17 aB | 24.09 ± 0.07 cB | 31.96 ± 0.39 bB |
| TN (g kg−1) | 3.96 ± 0.04 aA | 3.01 ± 0.02 bA | 3.11 ± 0.03b A | 3.81 ± 0.04 aB | 2.94 ± 0.03 bB | 2.72 ± 0.02 cB |
| TP (g kg−1) | 0.840 ± 0.007 a | 0.530 ± 0.004 cA | 0.556 ± 0.005 bA | 0.814 ± 0.013 a | 0.572 ± 0.011 bB | 0.518 ± 0.004 cB |
| DTN (mg kg−1) | 17.23 ± 0.36 bA | 10.32 ± 0.33 cA | 21.14 ± 0.69 aA | 10.65 ± 0.44 aB | 6.10 ± 0.13 bB | 6.44 ± 0.23 bB |
| NN (mg kg−1) | 10.55 ± 0.48 bA | 7.74 ± 0.48 cA | 15.94 ± 1.13 aA | 2.74 ± 0.27 aB | 1.50 ± 0.16 bB | 2.26 ± 0.20 aB |
| AN (mg kg−1) | 3.32 ± 0.14 bA | 2.65 ± 0.06 cA | 4.25 ± 0.11 aA | 1.82 ± 0.11 aB | 1.47 ± 0.02 bB | 1.48 ± 0.04 bB |
| SAP (mg kg−1) | 10.03 ± 0.26 aA | 5.06 ± 0.07 bA | 2.63 ± 0.07 cA | 9.06 ± 0.15 aB | 3.59 ± 0.04 bB | 1.79 ± 0.09 cB |
| SMBC (mg kg−1) | 230.71 ± 10.30 c | 303.18 ± 11.05 b | 423.16 ± 8.79 a | ND | ND | ND |
| SMBN (mg kg−1) | 53.92 ± 2.95 b | 72.31 ± 3.08 a | 80.51 ± 3.92 a | ND | ND | ND |
| SMBP (mg kg−1) | 24.68 ± 0.51 a | 9.23 ± 0.43 c | 13.19 ± 0.86 b | ND | ND | ND |
| BG (nmol h−1 g−1) | 186.15 ± 7.11 a | 122.07 ± 6.33 b | 199.80 ± 4.95 a | ND | ND | ND |
| NAG (nmol h−1 g−1) | 37.79 ± 2.80 c | 66.87 ± 3.87 b | 255.20 ± 4.82 a | ND | ND | ND |
| AP (nmol h−1 g−1) | 151.75 ± 6.27 a | 97.55 ± 2.95 c | 124.96 ± 3.58 b | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Yu, L.; Yang, X.; Luo, L.; Liu, J.; Chen, J.; Kou, Y.; Zhao, W.; Liu, Q. Effects of Different Soils on the Biomass and Photosynthesis of Rumex nepalensis in Subalpine Region of Southwestern China. Forests 2022, 13, 73. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010073
He H, Yu L, Yang X, Luo L, Liu J, Chen J, Kou Y, Zhao W, Liu Q. Effects of Different Soils on the Biomass and Photosynthesis of Rumex nepalensis in Subalpine Region of Southwestern China. Forests. 2022; 13(1):73. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010073
Chicago/Turabian StyleHe, Heliang, Lan Yu, Xiaocheng Yang, Lin Luo, Jia Liu, Jing Chen, Yongping Kou, Wenqiang Zhao, and Qing Liu. 2022. "Effects of Different Soils on the Biomass and Photosynthesis of Rumex nepalensis in Subalpine Region of Southwestern China" Forests 13, no. 1: 73. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010073