
Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting

National Engineering Laboratory for Tree Breeding, Beijing Forestry University, Beijing 100083, China
*
Author to whom correspondence should be addressed.
Forests 2022, 13(2), 354; https://0-doi-org.brum.beds.ac.uk/10.3390/f13020354
Submission received: 20 January 2022
/
Revised: 15 February 2022
/
Accepted: 16 February 2022
/
Published: 19 February 2022
(This article belongs to the Section Genetics and Molecular Biology)
Abstract
:Messenger RNAs exchanged between scions and rootstocks of grafted plants seriously affect their traits performance. The study goals were to identify the long-distance mRNA transmission events in grafted willows using a transcriptome analysis and to reveal the possible effects on rooting traits. The results showed that the Salix matsudana variety 9901 has better rooting ability than YJ, which reasonably improved the rooting performance of the heterologous grafts 9901 (scion)/YJ (rootstock). A transcriptome analysis showed that 2948 differentially expressed genes (DEGs) were present in the rootstock of 9901/YJ grafted plants in comparison with YJ/YJ. Among them, 692 were identified as mRNAs moved from 9901 scion based on SNP analysis of two parents. They were mostly 1001–1500 bp, had 40–45% GC contents, or had expression abundance values less than 10. However, mRNAs over 4001 bp, having 50–55% GC contents, or having expression abundance values of 10–20 were preferentially transferred. Eight mRNAs subjected to long-distance trafficking were involved in the plant hormone pathways and may significantly promote the root growth of grafted plants. In summary, heterologous grafts of Salix matsudana could efficiently influence plant rooting of the mRNAs transport from scion to rootstock.
1. Introduction
Grafting is used for the asexual propagation of woody plants, allowing the rapid proliferation and preservation of improved lines [1], as well as the shortening of flowering plants’ breeding periods, without altering genotypes [2]. Occasionally, grafting results in changes in plant traits. For example, Siberian pine (scion) grafted onto Scots pine (rootstock) (presented as scion/rootstock) grew faster and taller than Siberian/Siberian pines [1]. A wild-type (WT) tomato plant grafted on top of the mouse ear leaf mutant resulted in the scion leaves being phenotypically similar to the mutant leaves [3]. The tomato grafts displayed leaf phenotype changes from the original multi-serrated angustifoliate leaves of the scion to the ovate leaves of the transgenic rootstock containing the Cmgaip gene (Cucurbita maxima) [4]. And, grafting a watermelon scion onto a Cucurbit rootstock effectively improved the plant’s resistance to wilt disease [5], drought [6], cold [7], and heavy metal pollution [8].
Several studies had explored the variations mechanisms of the grafted plants and revealed that the transcripts could travel long distances between scions and rootstocks. For example, the CmGAI transcript has been readily detected in the WT scion grafted onto a CmGAI transgenic tomato rootstock [4]. In potato grafts, BEL5 transcripts from the BEL5-overexpressing scion have been found in the WT rootstocks [9]. The transcriptome was analyzed to determine the transcript compositions in heterologous grafts using single nucleotide polymorphisms (SNPs). Large numbers of mRNAs were identified as having undergone long-distance movement. In grape [10], 1963 mRNAs were transferred from scions to rootstocks, while 2210 mRNAs were transferred from rootstocks to scions. Further studies revealed that highly expressed genes might generate great quantities of mobile mRNAs and that ~10% of the mobile mRNAs showed large differences in their transmission ratio owing to different field conditions. Genotypes, graft combinations and growth environments impacted the direction of the mRNA movement, as well as the numbers and species of mRNAs being exchanged. They suggested the existence of passive and selective mechanisms of RNA movement. More evidence was provided in other grafting assays such as Arabidopsis thaliana (2006 mobile mRNAs) [11], cucumber (3546 mobile mRNAs) [12], and tobacco (1163 mobile mRNAs) [13].
Salix matsudana is a widely planted willow species that has rich germplasm resources and plays an important role in greening, ecological protection, and conifer utilization. Here, the study goals were to identify the long-distance mRNA transmission events in grafted willows using transcriptome analysis and to reveal the possible effects on rooting traits. Characterization of the transferred mRNAs will provide insights into long-distance mobile mRNAs mechanisms and help improve grafting systems.
2. Materials and Methods
2.1. Plant Materials and Grafting
Two-month-old branches (with length of 15 cm) of the Salix mandshurica variety 9901 and Yanjiang (YJ) were used as sources to construct the homologous and heterologous grafts 9901/9901, YJ/YJ, and 9901/YJ by V-shaped grafting method. The grafted segments were grown in hydroponics condition (changed every other day) in a growth chamber of a 16/8 h light/dark cycle (2000 lux) and relative humidity of 75–80% at 25 °C. The total root length was measured for each plant starting from the 10th day of grafting. Permission for collection, planting and grafting of the willow plants was obtained from Beijing Forestry University. The study was in accordance with relevant institutional, national, and international guidelines and legislation.
2.2. Extraction of Total RNA
On 22th day of grafting, the bark tissue (containing phloem for organic substances transporting) of 9901/9901, YJ/YJ and 9901/YJ rootstocks were individually sampled (10–12 cm below the grafting joint) for total RNA extraction by using polysaccharide polyphenol kit (TIANGEN Company, Beijing, China). The isolated RNA was treated with RNase-free DNase I (Takara, Kusatsu-shi, Japan) for 30 min at 37 °C to remove the residual DNA. RNA quality was tested by using NanoDrop2000 spectrophotometer (NanoDrop, Thermo Scientific, Waltham, MA, USA). RNA integrity was assessed using the RNA Nano 6000 Assay Kit of the Agilent Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA).
2.3. RNA Library Construction and Sequencing
Five plants of each type of grafting (9901/9901 or 9901/YJ or YJ/YJ) were selected and bulked for RNA sequencing library construction using NEBNext UltraTM RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA) following the manufacturer’s instruction. The library was sequenced on an Illumina platform (Biomaker Company, Beijing, China) and the paired-end reads were generated. The 10 Gb raw data was gained for each sample and the clean data were obtained by removing adapter and low-quality reads from the raw data. Q20, Q30, GC-content and sequence duplication level of the clean data were calculated. All the analyses were based on high-quality clean data.
2.4. Identification of the Differentially Expressed Genes (DEGs) and Function Analysis
The reads per kilobase per million mapped reads (RPKM) were generated to represent the expression abundance of each mRNA. Differential expression analysis of two data sets from the rootstocks of (YJ/YJ and 9901/YJ was performed using the edgeR. The FDR < 0.01 and Fold Change ≥ 2 was as the threshold for significantly differential expression. Gene Ontology (GO) enrichment analysis of the DEGs was implemented by the GOseq R packages based Wallenius non-central hyper-geometric distribution, which can adjust for gene length bias in DEGs [14]. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were performed using Blast2GO software (BioBam Bioinformatics SL, Valencia, Spain) [15].
2.5. Determination of the Transfer RNAs
All the clean reads of 9901/9901, YJ/YJ and 9901/YJ were compared by using HISAT2 software [16]. SNPs between two parents were identified with GATK software [17]. The reads (transcripts) in rootstocks of 9901/YJ having SNPs with YJ/YJ but homozygous with 9901/9901 were determined as the mobile mRNAs from 9901 scion to YJ rootstock in 9901/YJ grafts.
2.6. Estimation of RNA Transfer Ratio
Based on the expression abundance of the mobile mRNAs in 9901/YJ, the RNA transfer ratio from 9901 to YJ in 9901/YJ was estimated (the normalized number of the 9901-sourced mobile RNAs in the rootstock of 9901/YJ divided by the normalized number of the total RNAs in the rootstock of 9901/YJ). When SNPs were present in several mobile reads, the average value was used as the transfer ratio of the mobile RNA. The mobile RNA with less than 30 mapped reads in the graft was not considered for transfer ratio estimation to reduce the bias.
3. Results
3.1. 9901 Scions Enhanced the Rooting of YJ Rootstocks in 9901/YJ Grafts
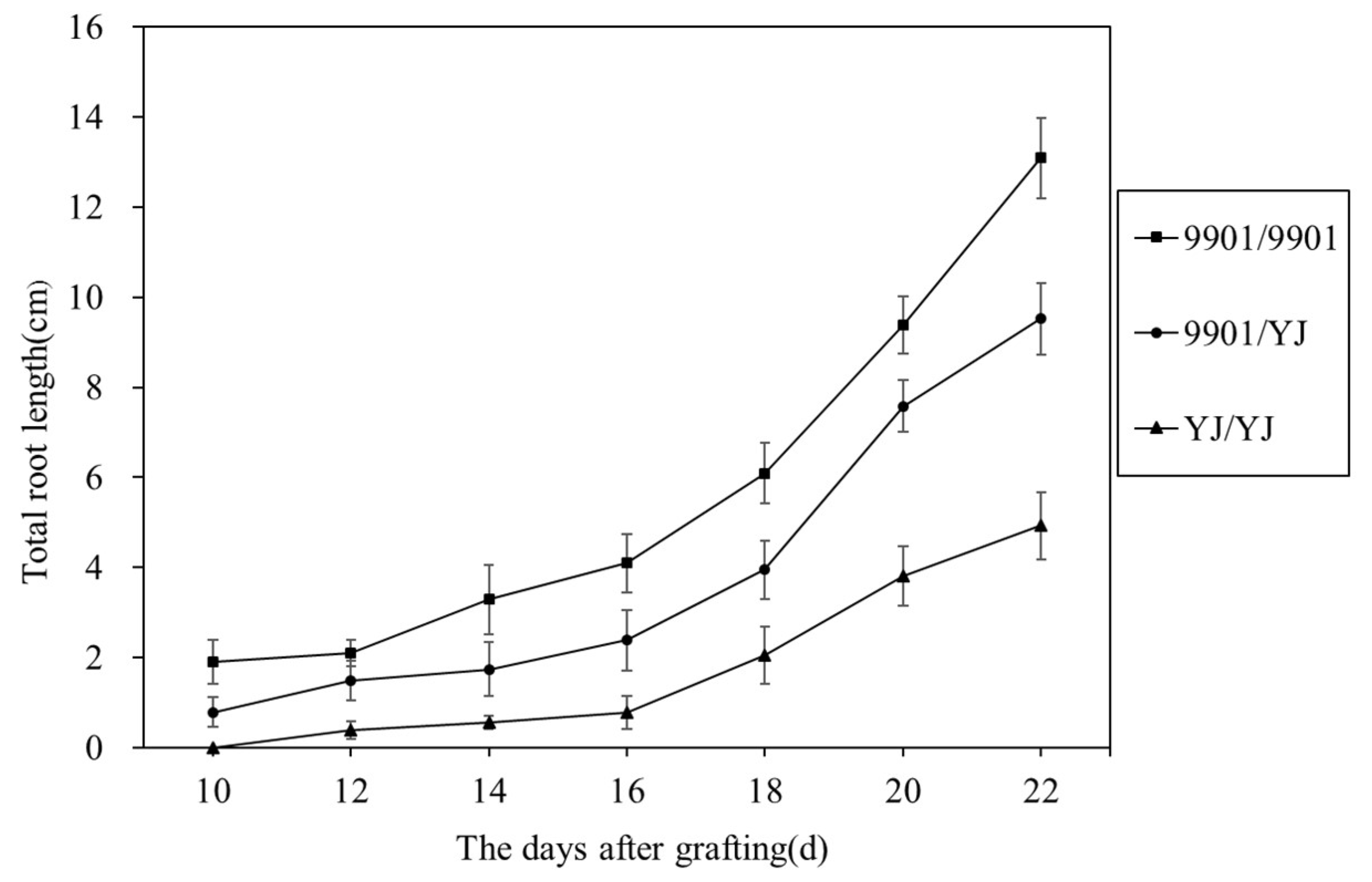
During hydroponic culturing, the grafted willows started rooting at the bases of the rootstocks. Obviously, the 9901/9901 grafts rooted faster than the YJ/YJ plants (Figure 1). The total root length of the 9901/9901 grafts was 2.66 times that of the YJ/YJ at 22nd after grafting, indicating that 9901 has a strong rooting potential than YJ (Figure 2). Meanwhile, the 9901/YJ plants rooted faster than YJ/YJ plants, and the total root length of the 9901/YJ grafts was 1.93 times that of the YJ/YJ grafts. This revealed that the 9901 scions had a certain promotive effect on the rooting of the YJ rootstocks.
3.2. DEGs in Homologous and Heterologous Willow Grafts
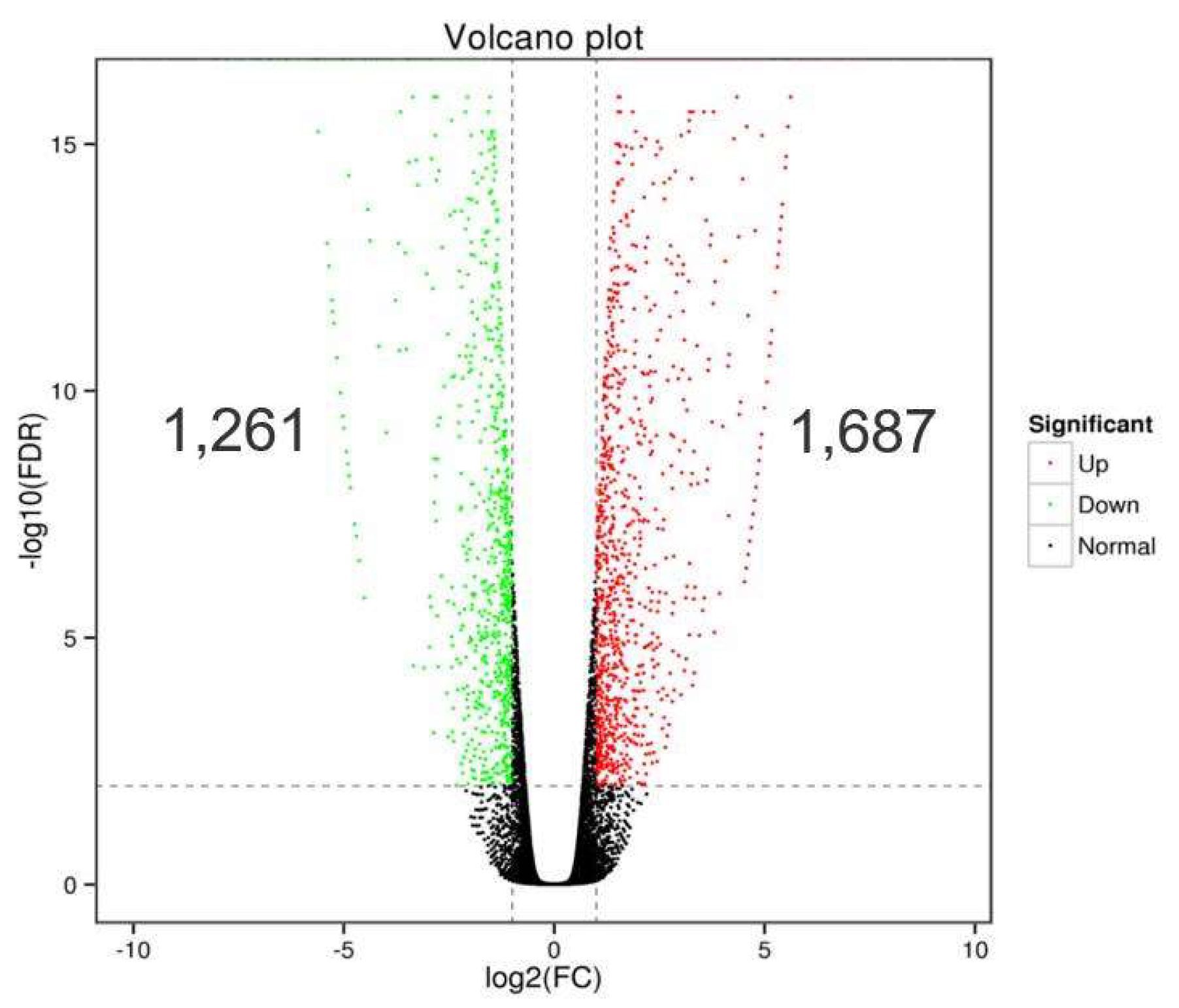
A total of 49,425, 48,176 and 48,713 genes were identified in the rootstock tissues of 9901/9901, YJ/YJ, and 9901/YJ grafted plants, respectively (Figure 3), based on transcriptome analysis. In total, 2,948 DEGs were identified between YJ/YJ and 9901/YJ. Among them, 1687 and 1261 genes were significantly up- and down-regulated in 9901/YJ compared with YJ/YJ.
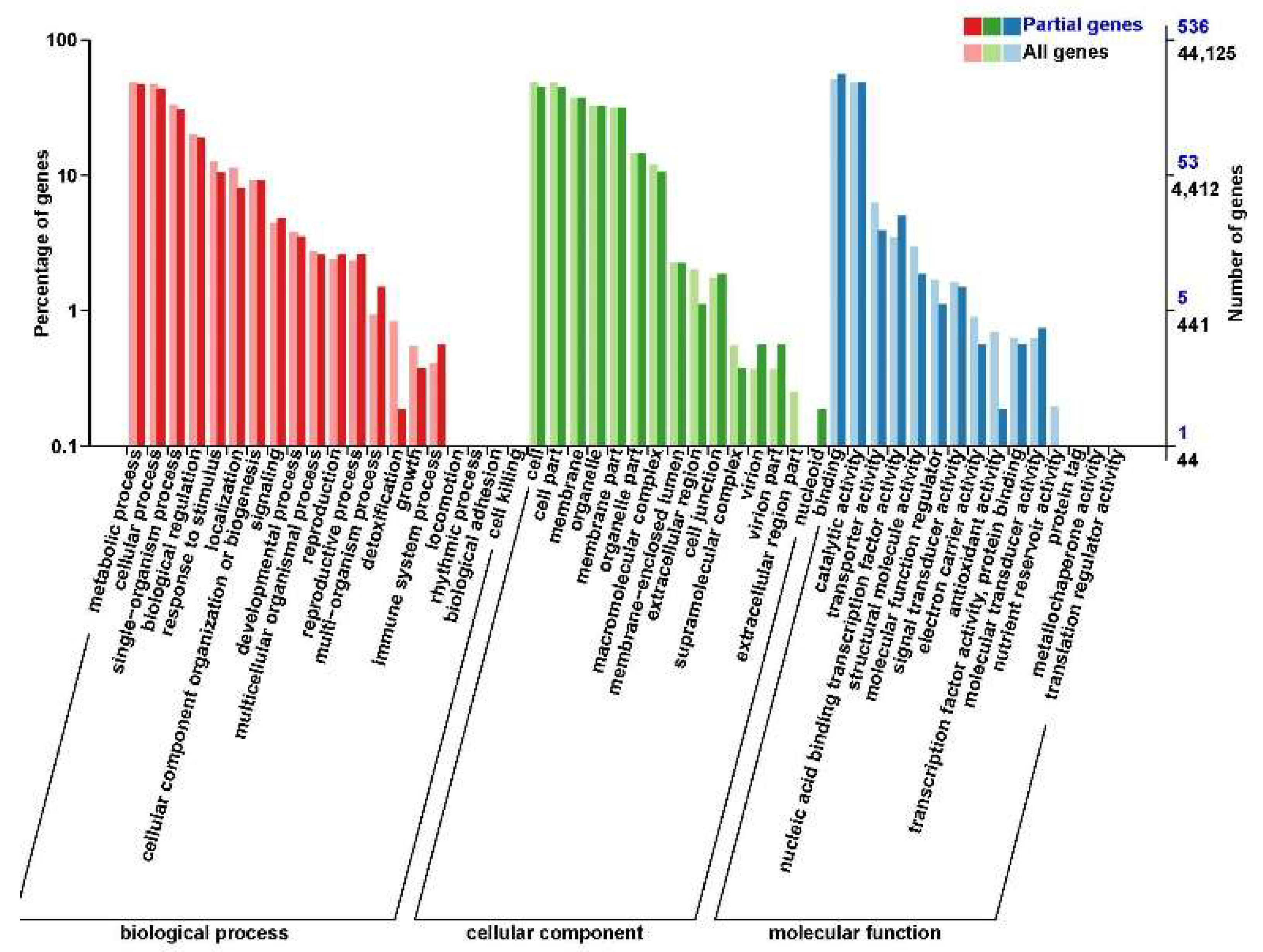
A total of 2309 DEGs (Figure S1) were annotated in the GO database. The DEGs related to biological processes were grouped in 21 terms, which were mainly involved in metabolic (1100 genes), cellular (983 genes), and single-organism (916 genes) processes. The DEGs related to cellular components were grouped in 15 terms, which were mainly involved in cell (999 genes), cell part (975 genes), membrane (931 genes), and organelle (571 genes). The DEGs related to molecular function were grouped in 15 terms, which were mainly involved in catalytic activity (1226 genes), binding (1170 genes), and transporter activity (216 genes).
3.3. SNP Analysis Revealed 692 Shoot-to-Root Mobile mRNAs in 9901/YJ Grafts
SNP analysis was conducted between the homologous genes of two grafts (YJ/YJ, 9901/YJ), which showed 692 of 48,713 mRNAs in rootstocks of 9901/YJ came from the scions. Among them, 536 mRNAs were annotated in the GO database (Figure 4). In total, 16 annotated terms were related to biological processes, including metabolic processes (253 mRNAs), cellular processes (234 mRNAs), and single-organism processes (163 mRNAs). There were 14 annotated terms related to cellular component, including cell (240 mRNAs), cell part (239 mRNAs), and membrane (201 mRNAs), and there were 11 annotated terms related to molecular function, including binding (301 mRNAs) and catalytic activity (259 mRNAs).
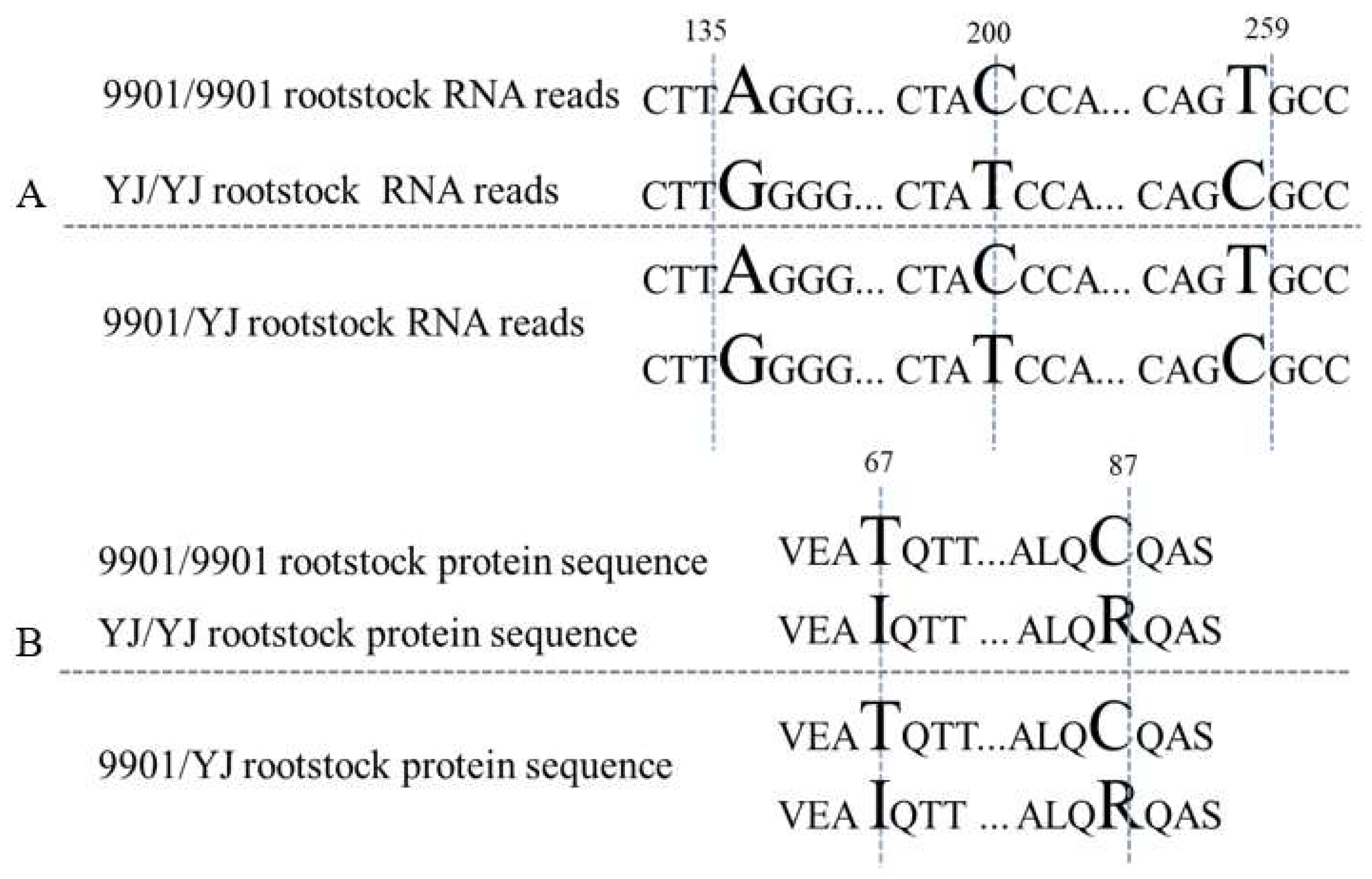
An example was given in detail. The transcriptome analysis showed that ABF2 (ABA-responsive element binding factor) (K14432) gene was 966 bp in both parents but displayed three SNPs at site of 135, 200, and 259 bp. The SNPs resulted in coding sequence changes in two amino acids. The gene fragments were amplified and cloned from the rootstock samples of 9901/9901, YJ/YJ, and 9901/YJ. The sequencing results showed that 9901/9901 or YJ/YJ individually had one specific fragment with the SNPs existing the same as the transcriptome data, while the heterografting rootstock had both fragments (Figure 5).
3.4. Characterization of the Mobile mRNAs in Grafted Willows
The transfer ratio of 455 mobile mRNAs was estimated (the others, having less than 30 reads, were not considered), and they ranged from 7.14–68.75% with an average of 27.30%. Among them, 29.89%, 23.74%, and 20.88% of the mobile mRNAs had a transfer ratio of 10–20%, 20–30%, and 30–40%, respectively.
The maximum size of a mobile mRNA was 10,710 bp and the minimum was 186 bp, with an average of 1657 bp, which was 19.64% greater than the average 1385 bp length of all willow transcripts. Based on mRNA length, the 455 mobile mRNAs were classified into several groups. The 1000–1500 bp group contained the largest proportion, at 26.59%, of all the mobile mRNAs, followed by the 501–1000 bp and 1501–2000 bp groups. However, the mRNAs > 4001 bp were preferred for transfer, accounting for 2.40% of the mobile mRNAs in the group, followed by the 3501–4000 and 2,501–3,000-bp mRNAs (Table 1).
GC content of all the transcripts in willow was calculated in a range of 28.70–67.84%, on average 44.37%. Comparably, GC content of the 455 mobile mRNAs ranged from 36.40% to 57.89%, with an average of 44.18%. The 455 mobile mRNAs were grouped by their GC content (Table 2). Among them, the mRNAs with GC content of less than 35% were not transferred. More mobile mRNAs (64.18%) had GC content of 40–45%, followed by mRNAs with GC content of 45–50%. The transfer ratio calculation showed that the mRNAs with GC content of 50–55% had the largest value at 1.20%, followed by the mRNAs with GC content of 40–45%.
The expression abundances (presented as FPKM values) of the mobile mRNAs (Table 3) revealed minimum and maximum expression levels of 0.42 and 817.09, respectively, with an average of 19.81. Comparably, the average expression level of all the mRNAs (16.83) was 17.74% lower than that of the mobile mRNAs. The mobile mRNAs were classified into several groups based on the expression abundance. Among them, the mRNAs with expression value less than 10 formed the largest group of the mobile mRNAs (63.30%), followed by the mRNAs with expression value of 10–20 of the mobile mRNAs (21.10%). The transfer efficiency evaluation showed that mRNAs with expression value of 10–20 accounted for 1.12% of the transcripts, followed by the mRNAs with expression values of 40–50 (1.11%).
3.5. The Mobile mRNAs and Pathways Involved in Roots Formation
A total of 142 (20.52%) mobile mRNAs were annotated to 85 pathways in the KEGG database. The main pathways included spliceosome, biosynthesis of amino acids, plant-pathogen interaction, and protein processing in endoplasmic reticulum (Table 4), which enriched with 11, 11, 10, and 10 mobile mRNAs, respectively.
Notably, the plant hormone signal transduction was enriched with eight mobile mRNAs (Figure S2), which participated in auxin (AUX), cytokinin (CK), gibberellin (GA), abscisic acid (ABA), jasmonates (JAs), and salicylic acid (SA) metabolism, with transfer ratio of 14.72–68.29% (Table 5). They may potentially participate in root formation as shown in the previous reports.
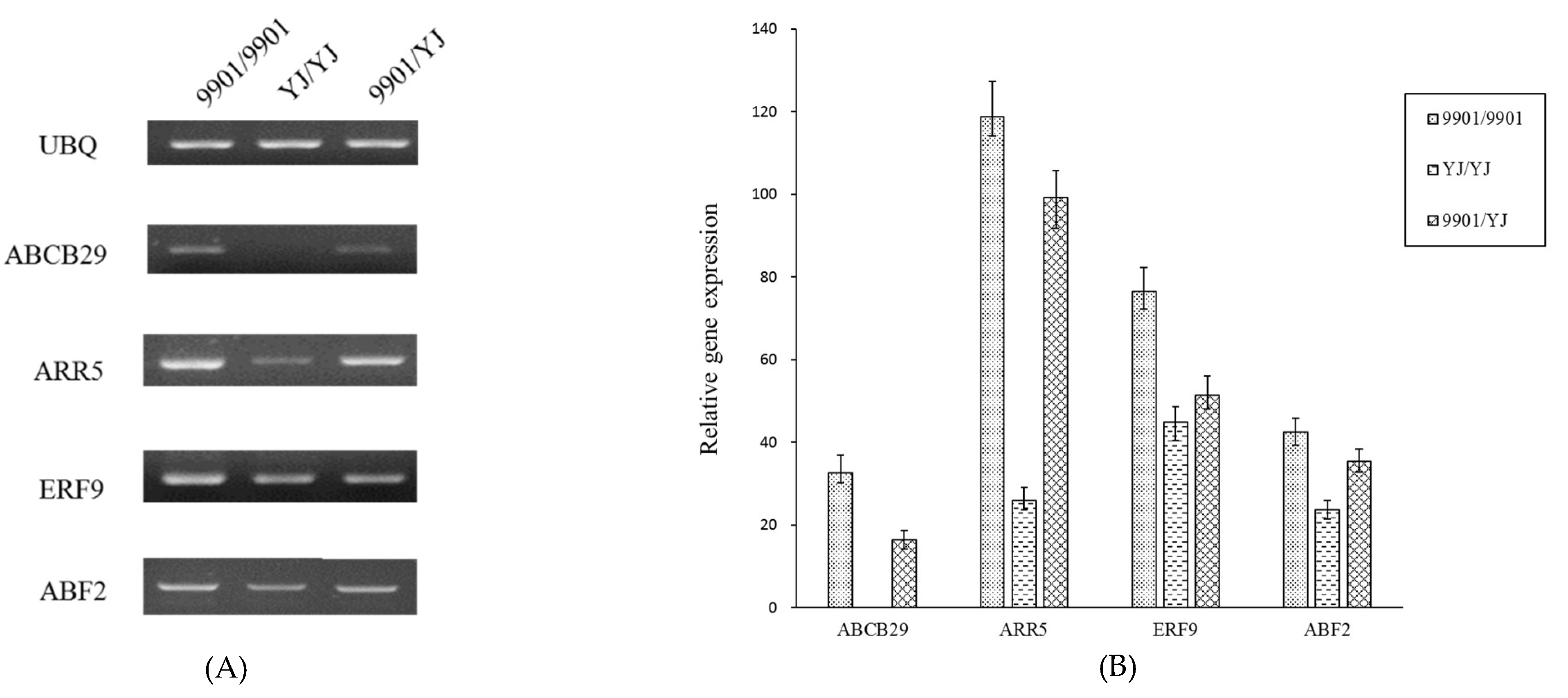
Several mobile mRNAs were identified in association with rooting as the previous reports such as ABCB29 [18], ARR5 [19], ERF9 [20], and ABF [21]. PCR tests showed them well expressed in 9901 rootstock of 9901/9901 plants, but less in YJ rootstock of YJ/YJ plants (Figure 6). These genes also had more transcripts in YJ rootstock of 9901/YJ plants than in YJ rootstock of YJ/YJ plants, potentially gained from its 9901scion, and improved rooting of 9901/YJ plants as to that of YJ/YJ plants.
4. Discussion
4.1. The Number of Mobile mRNAs Varied Vastly in Different Grafts
Long-distance mRNA movement events occur in grafts frequently, but the number of mobile mRNAs varied greatly by graft parents. For example, 422 bottle-gourd (LAG) and 441 watermelon (WM) mobile transcripts were identified in WM/LAG and LAG/WM heterografts [22], 138 mobile mRNAs were identified in Nicotiana benthamiana/Arabidopsis grafts [23], and 23 mobile mRNAs were identified in Canola/Arabidopsis grafts [13]. In our study here, both grafting willow parents were bred from the same species of Salix matsudana, more than 692 mobile mRNAs were identified in heterologous grafts. Obviously, the number of the mobile mRNAs is correlated with the affinity between the scion and rootstock. Grafts by two closely related varieties of a species could transfer more mRNAs than grafts by more distantly related species.
The numbers of mobile mRNAs are also associated with the sampling distance from the grafting joint. In a previous report, the number of mobile mRNAs from the tissues below the grafting joint of N. benthamiana (scions)/tomato (stock) were decreased from the high stem (723) to the low stem (663) and the root (242) along the movement path [13]. Similarly, grafting of two different Arabidopsis thaliana ecotypes showed a decreasing number of mobile root-produced transcripts in the lower (225) and upper stem (88) tissues [11]. And, 26 of all 739 root-exported mRNAs were exclusively detected in the inflorescence, indicating the tissue-specific accumulation of mobile transcripts. In our experiment, we sampled the tissues near the base of the cuttings. Some mobile mRNAs and pathways were identified and clearly displayed their potential to the rooting activity improvement in heterologous grafts. Therefore, sampling distance from the grafting joint is very important to gain useful mobile RNAs for trait performance.
4.2. The Mobile mRNAs Efficiently Affect Plant Performance
The study here showed that 9901 scions could really transmit their strong rooting ability to the YJ rootstock, thereby improving rooting by as much as 1.93 times that of YJ/YJ. Transcriptome analysis showed that several mobile mRNAs in relation to hormones metabolism were involved in the process. For example, two ABF genes in the ABA metabolic pathway were determined mobile in willow grafts at a ratio of 1.91% and 15.24%, respectively. As a transcription factor, the ABF gene regulates downstream genes expression such as the basic leucine zipper transcription factor of bZIP4. The ZmbZIP4 over-expression lines had longer primary, seminal, and lateral roots in comparison with those of the WT plants [24]. Therefore, the significance of the mobile ABF genes from 9901 line with strong rooting activity is understandable. In others, a mobile mRNA in relation to AUX/IAA was detected in willow grafts, which was also frequently mobile in some other grafts [9,25,26]. The AUX/IAA gene may activate the downstream transcription factor ARF and further regulate the root development [27,28,29]. Some more 9901-sourced mobile genes were supposed to have a functional characterization using CRISPR-Cas9 or related techniques in the future.
4.3. The Mobile mRNAs in Grafts Are of Some Sequence Characteristics
mRNA length had significant effects on the mobile efficiency in grafts. More than 70% of the mobile mRNAs in grafts were less than 2000 bp in the previous study of Arabidopsis [11], N. benthamiana [13], cucumber [12], and grape [10]. Similar results were also found in willow grafts. Especially, the transfer ratio of mRNAs > 4000 bp was 2.4%, which is 3.13 times that of mRNAs < 2000 bp. And in Arabidopsis, the transfer ratio of mRNAs > 4000 bp reached 11.52%, which was 2.55 times that of mRNAs < 2000 bp. Thus, it was common that the larger mRNA fragments were preferentially transferred. the larger mRNA fragments may easily form certain stable spatial structures and are suited for long-distance transfers.
A computational diffusion-based model had shown that mRNA abundance is an important factor in mRNA’s long-distance mobility [30]. In willow grafts, mRNAs with the expression abundances of 10–20 and 40–50 had high transfer efficiencies, being 1.62 and 1.61 times that of mRNAs with expression abundances of 30–40. Unfortunately, the correlation between mRNA expression abundance and the transfer ratio was not perfectly defined in several experiments. In N. benthamiana grafts, only 3 of 183 mobile mRNA were highly ranked among all the transcribed genes based on their expression abundance levels [13]. Even, the over-expressed GUS and GFP genes under control of the 35S promoter [31,32] and AtAMT1 and AtCHL1 genes under control of the strong phloem-specific promoter SUC [13] did not undergo long-distance transfer. Possibly, the effect of gene expression level on their mobile ratio in grafts might be in combination with some other mechanisms.
GC content may also influence mRNA mobile. In willow grafts, 95.16% of mobile mRNAs had GC content between 30% and 50%. However, mRNAs with GC content of 50–55% were preferentially transferred with a transfer ratio of 1.20%, which was 1.60 times that of mRNAs with GC contents greater than 55% and 1.13 times that of mRNAs with GC contents less than 50%. Similarly in Arabidopsis grafts, mRNAs with GC contents of 50–55% exhibited the highest transfer ratio of 8.45%, which is 1.08 times that of mRNAs with GC contents >55% and 1.54 times that of the mRNAs with GC contents <50% [11]. Therefore, sequence characteristics of the mobile mRNAs in grafts should be of comprehensive effect and be much concerned in the future.
5. Conclusions
Heterologous grafts of Salix matsudana could efficiently influence plant rooting since the mRNAs transport from scion to rootstock. These mobile mRNAs are of some sequence characteristics such as length, GC content and expression abundance. And, the number of the mobile mRNAs is in relation to the affinity of the grafting parents, which deserved serious consideration in the grafting program to gain the expected results.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13020354/s1, Figure S1: GO annotation of DEGs in the grafted willows of 9901/YJ comparing with YJ/YJ; Figure S2: The mobile genes (in gray) involved in plant hormone signal transduction pathway.
Author Contributions
Conceptualization, J.X.; Methodology, P.Y.; Investigation, P.Y., B.L.; Data Curation, Y.C., Y.W.; Writing–Original Draft Preparation, P.Y., X.L.; Writing–Review & Editing, J.X.; Supervision, J.X.; Funding Acquisition, J.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 31870648.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequencing data is available in NCBI (accession# PRJNA616160).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Darikova, J.A.; Savva, Y.V.; Vaganov, E.A.; Grachev, A.M.; Kuznetsova, G.V. Grafts of woody plants and the problem of incompatibility between scion and rootstock. J. Sib. Fed. Univ. Biol. 2011, 4, 54–63. [Google Scholar]
- Haroldsen, V.M.; Szczerba, M.W.; Aktas, H.; Lopez-Baltazar, J.; Odias, M.J.; Chi-Ham, C.L.; Labavitch, J.M.; Bennett, A.B.; Powell, A.L.T. Mobility of transgenic nucleic acids and proteins within grafted rootstocks for agricultural improvement. Front. Plant Sci. 2012, 3, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Canio, W.; Kessler, S.; Sinha, N. Developmental changes due to long-distance movement of a homeobox fusion mRNA in tomato. Science 2001, 293, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Haywood, V.; Yu, T.; Huang, N.; Lucas, W.J. Phloem long-distance trafficking of gibberellic acid-insensitive RNA regulates leaf development. Plant J. 2005, 42, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Gaion, L.A.A.; Braz, L.T.; Carvalho, R.F. Grafting in vegetable crops: A great technique for agriculture. Int. J. Veg. Sci. 2018, 24, 85–102. [Google Scholar] [CrossRef]
- Sanchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Blasco, B.; Leyva, R.; Romero, L.; Ruiz, J.M. Antioxidant response resides in the shoot in reciprocal grafts of drought-tolerant and drought-sensitive cultivars in tomato under water stress. Plant Sci. 2012, 188–189, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, J.; Huang, L.; Ding, X.; Shi, K.; Yu, J. Grafting of Cucumis sativus onto Cucurbita ficifolia leads to improved plant growth, increased light utilization and reduced accumulation of reactive oxygen species in chilled plants. J. Plant Res. 2009, 122, 529–540. [Google Scholar] [CrossRef]
- Savvas, D.; Colla, G.; Rouphael, Y.; Dietmar, S. Amelioration of heavy metal and nutrient stress in fruit vegetables by grafting. Sci. Hortic. 2010, 127, 156–161. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell. 2006, 18, 3443–3457. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Mao, L.; Jittayasothorn, Y.; Kang, Y.; Jiao, C.; Fei, Z.; Zhong, G. Messenger RNA exchange between scions and rootstocks in grafted grapevines. BMC Plant Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [Green Version]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Miñambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 15025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, Y.; Ham, B.; Chen, J.; Yoshida, A.; Kochian, L.V.; Fei, Z.; Lucas, W.J. Vascular-mediated signalling involved in early phosphate stress response in plants. Nat. Plants 2016, 2, 16033. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zheng, Y.; Huang, J.; Zhou, X.; Li, R.; Zhao, M.; Wang, S.; Huang, Z.; Lan, H.; Turgeon, R.; et al. Elucidation of the mechanisms of long-distance mRNA movement in a Nicotiana benthamiana/tomato heterograft system. Plant Physiol. 2018, 177, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Gotz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3425. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Sukumar, P.; Maloney, G.S.; Muday, G.K. Localized induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis. Plant Physiol. 2013, 162, 1392–1405. [Google Scholar] [CrossRef]
- Cheng, X.; Jiang, H.; Zhang, J.; Qian, Y.; Zhu, S.; Cheng, B. Overexpression of type-A rice response regulators, OsRR3 and OsRR5, results in lower sensitivity to cytokinins. Genet. Mol. Res. 2010, 9, 348–359. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, S.; Song, Y.; Huang, Y.; Zhou, S.; Liu, X.; Zhou, D. The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling. Plant Cell 2015, 27, 2469–2483. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, J.; Cho, D.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lozano, M.; Dutta, S.K.; Natarajan, P.; Tomason, Y.R.; Lopez, C.; Katam, R.; Levi, A.; Nimmakayala, P.; Reddy, U.K. Transcriptome changes in reciprocal grafts involving watermelon and bottle gourd reveal molecular mechanisms involved in increase of the fruit size, rind toughness and soluble solids. Plant Mol. Biol. 2020, 102, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Notaguchi, M.; Higashiyama, T.; Suzuki, T. Identification of mRNAs that move over long distances using an RNA-seq analysis of Arabidopsis/Nicotiana benthamiana heterografts. Plant Cell Physiol. 2015, 56, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Liu, C.; Li, Z.; Ran, Q.; Xie, G.; Wang, B.; Fang, S.; Chu, J.; Zhang, J. ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol. 2018, 178, 753–770. [Google Scholar] [CrossRef] [Green Version]
- Kanehira, A.; Yamada, K.; Iwaya, T.; Tsuwamoto, R.; Kasai, A.; Mikio, N.; Harada, T. Apple phloem cells contain some mRNAs transported over long distances. Tree Genet. Genom. 2010, 6, 635–642. [Google Scholar] [CrossRef]
- Omid, A.; Keilin, T.; Glass, A.; Leshkowitz, D.; Wolf, S. Characterization of phloem-sap transcription profile in melon plants. J. Exp. Bot. 2007, 58, 3645–3656. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, H.; Estelle, M. Plant development: Regulation by protein degradation. Science 2002, 297, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Maraschin, F.S.; Memelink, J.; Offringa, R. Auxin-induced, SCFTIR1-mediated poly-ubiquitination marks AUX/IAA proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef]
- Li, J.; Dai, X.; Zhao, Y. A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 2006, 140, 899–908. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, A.; Kopriva, S.; Morris, R. Transcript abundance explains mRNA mobility data in Arabidopsis thaliana. Plant Cell 2016, 28, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Thieme, C.; Kollwig, G.; Apelt, F.; Yang, L.; Winter, N.; Andresen, N.; Walther, D.; Kragler, F. tRNA-related sequences trigger systemic mRNA transport in plants. Plant Cell 2016, 28, 1237–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yu, P.; Zhao, J.; Jiang, H.; Wang, H.; Zhu, Y.; Botella, M.A.; Samaj, J.; Li, C.; Lin, J. Expression of tomato prosystemin gene in Arabidopsis reveals systemic translocation of its mRNA and confers necrotrophic fungal resistance. New Phytol. 2018, 217, 799–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Rooting of the willow grafts 9901/9901, YJ/YJ, and 9901/YJ.

Figure 2.
The total root length of willow grafts in hydroponic culture.

Figure 3.
Volcano plot of DEGs in rootstocks of 9901/YJ compared with YJ/YJ.

Figure 4.
GO annotation of the transfer genes from scion to rootstock in 9901/YJ.

Figure 5.
SNP analysis of the ABF2 gene (K14432) in the rootstock of 9901/YJ. (A), nucleotide; (B), amino acids.
Figure 5.
SNP analysis of the ABF2 gene (K14432) in the rootstock of 9901/YJ. (A), nucleotide; (B), amino acids.

Figure 6.
Expression tests of the mobile mRNAs in rootstock of the grafted plants. (A), PCR results of four mobile genes (ABCB29, ARR5, ERF9, ABF2) with UBQ as the reference; (B), The quantitative PCR results.
Figure 6.
Expression tests of the mobile mRNAs in rootstock of the grafted plants. (A), PCR results of four mobile genes (ABCB29, ARR5, ERF9, ABF2) with UBQ as the reference; (B), The quantitative PCR results.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
mRNA length and transfer ratio in the rootstock of 9901/YJ.
| mRNA Length (bp) | Mobile mRNA Amount | Total Transcribed Gene Amount | Mobile mRNA Ratio in Total Transcribed Genes of the Group (%) a | Mobile mRNA Propotion in All Mobile mRNAs (%) b |
|---|---|---|---|---|
| 0–500 | 44 | 4883 | 0.90 | 9.67 |
| 501–1000 | 96 | 12,127 | 0.79 | 21.10 |
| 1001–1500 | 121 | 11,219 | 1.08 | 26.59 |
| 1501–2000 | 65 | 6441 | 1.01 | 14.29 |
| 2001–2500 | 46 | 3444 | 1.34 | 10.11 |
| 2501–3000 | 35 | 1901 | 1.84 | 7.69 |
| 3001–3500 | 17 | 1151 | 1.48 | 3.74 |
| 3501–4000 | 11 | 558 | 1.97 | 2.42 |
| 4001– | 20 | 835 | 2.40 | 4.39 |
a Ratio of the mobile mRNAs amount in the group and the total transcribed gene amount in the group; b Ratio of the mobile mRNAs amount in the group and the total mobile mRNAs (455) in rootstock of 9901/YJ.
Table 2.
GC content and transfer ratio in the rootstock of 9901/YJ.
| GC Content (%) | Mobile mRNA Amount | Total Transcribed Gene Amount | Mobile mRNA Ratio in Total Transcribed Gene of the Group (%) | Mobile mRNA Proportion in All Mobile mRNA (%) |
|---|---|---|---|---|
| 25–30 | 0 | 2 | 0 | 0 |
| 30–35 | 0 | 72 | 0 | 0 |
| 35–40 | 13 | 1353 | 0.96 | 2.86 |
| 40–45 | 292 | 25,776 | 1.13 | 64.18 |
| 45–50 | 128 | 13,471 | 0.95 | 28.13 |
| 50–55 | 21 | 1752 | 1.2 | 4.62 |
| 55– | 1 | 133 | 0.75 | 0.21 |
Table 3.
Gene expression abundance and transfer ratio in the rootstock of 9901/YJ.
| mRNA Expression Abundance (FPKM) | Mobile mRNAs Amount | Total Transcribed Gene Amount | Mobile mRNAs Ratio in Total Transcribed Gene of the Group (%) | Mobile mRNAs Proportion in All Mobile mRNAs (%) |
|---|---|---|---|---|
| 0–10 | 288 | 33,241 | 0.87 | 63.30 |
| 10–20 | 96 | 7113 | 1.12 | 21.10 |
| 20–30 | 39 | 3168 | 0.85 | 8.57 |
| 30–40 | 16 | 1734 | 0.69 | 3.52 |
| 40–50 | 12 | 1077 | 1.11 | 2.63 |
| 50–60 | 4 | 631 | 0.63 | 0.88 |
Table 4.
The main pathways of the mobile mRNAs involved.
| Pathway | ko-id | p-Value | Mobile mRNAs Amount |
|---|---|---|---|
| Spliceosome | ko03040 | 0.0606 | 11 |
| Biosynthesis of amino acids | ko01230 | 0.1549 | 11 |
| Plant-pathogen interaction | ko04626 | 0.0588 | 10 |
| Protein processing in endoplasmic reticulum | ko04141 | 0.1816 | 10 |
| Plant hormone signal transduction | ko04075 | 0.5378 | 8 |
| RNA transport | ko03013 | 0.3316 | 7 |
| Carbon metabolism | ko01200 | 0.7074 | 7 |
| Ribosome | ko03010 | 0.9146 | 7 |
| Fructose and mannose metabolism | ko00051 | 0.1093 | 4 |
| Carbon fixation in photosynthetic organisms | ko00710 | 0.2043 | 4 |
Table 5.
The mobile mRNAs involved in the plant hormone signal transduction pathway.
| Mobile mRNAs | Gene Function | Expression Abundance * | Transfer Ratio | ||
|---|---|---|---|---|---|
| 9901/9901 | YJ/YJ | 9901/YJ | |||
| AUX/IAA (K14484) | activation of the downstream transcription factor ARF | 0.50(0) | 0(0.14) | 0.37(0.26) | 58.73% |
| CRE1 (K14489) | transmembrane histidine kinase cytokinin receptor activity | 0.29(0) | 0(0.03) | 0.12(0.12) | 50.00% |
| B-ARR (K14491) | sequence-specific DNA binding | 0.45(0) | 0(0.11) | 0.04(0.04) | 50.00% |
| TF (K16189) | protein dimerization activity | 0.10(0) | 0(0.09) | 0.04(0.06) | 40.00% |
| ABF2 (K14432) | sequence-specific DNA binding | 4.20(0) | 0(0.84) | 0.08(0.40) | 16.67% |
| ABF5 (K14432) | sequence-specific DNA binding | 0.20(0) | 0(0.31) | 0.03(0.18) | 14.29% |
| JAZ (K13464) | transcription corepressor activity | 0.80(0) | 0(0.31) | 0.28(0.13) | 68.29% |
| NRP1 (K14508) | Regulatory protein | 0.23(0) | 0(0.38) | 0.10(0.37) | 21.28% |
* The data outside the parentheses means the expression abundance of 9901 homologous gene in rootstock of grafts, while the data inside the parentheses means the expression abundance of YJ homologous gene in rootstock of grafts.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yin, P.; Lan, B.; Liu, X.; Cui, Y.; Wang, Y.; Xu, J. Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting. Forests 2022, 13, 354. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020354
AMA Style
Yin P, Lan B, Liu X, Cui Y, Wang Y, Xu J. Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting. Forests. 2022; 13(2):354. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020354
Chicago/Turabian StyleYin, Peng, Baoliang Lan, Xiao Liu, Yu Cui, Yan Wang, and Jichen Xu. 2022. "Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting" Forests 13, no. 2: 354. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020354
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.