
Magnetic Treatment Improves the Seedling Growth, Nitrogen Metabolism, and Mineral Nutrient Contents in Populus × euramericana ‘Neva’ under Cadmium Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Materials
2.3. Experimental Design
- (1)
- (–MF, –Cd(NO3)2)—plants grown with half-strength modified Hoagland solution that were not subjected to magnetic treatment (NM0).
- (2)
- (+MF, –Cd(NO3)2)—plants grown with half-strength modified Hoagland solution that were subjected to magnetic treatment (M0).
- (3)
- (–MF, +Cd(NO3)2)—plants grown with half-strength modified Hoagland solution containing 100 μmol·L−1 Cd(NO3)2 that were not subjected to magnetic treatment (NM100).
- (4)
- (+MF, –Cd(NO3)2)—plants grown with half-strength modified Hoagland solution containing 100 μmol·L−1 Cd(NO3)2 that were subjected to magnetic treatment (M100).
2.4. Determination of Biochemical and Physiological Characteristics
2.4.1. Growth Character Analysis
2.4.2. Tissue Nitrogen Content Measurement
2.4.3. Tissue Ion Content Measurement
2.4.4. Enzyme Activity Determination
2.4.5. Amino Acid Content Estimation
2.4.6. Photosynthetic Pigment Observation
2.5. Statistical Analysis
3. Results
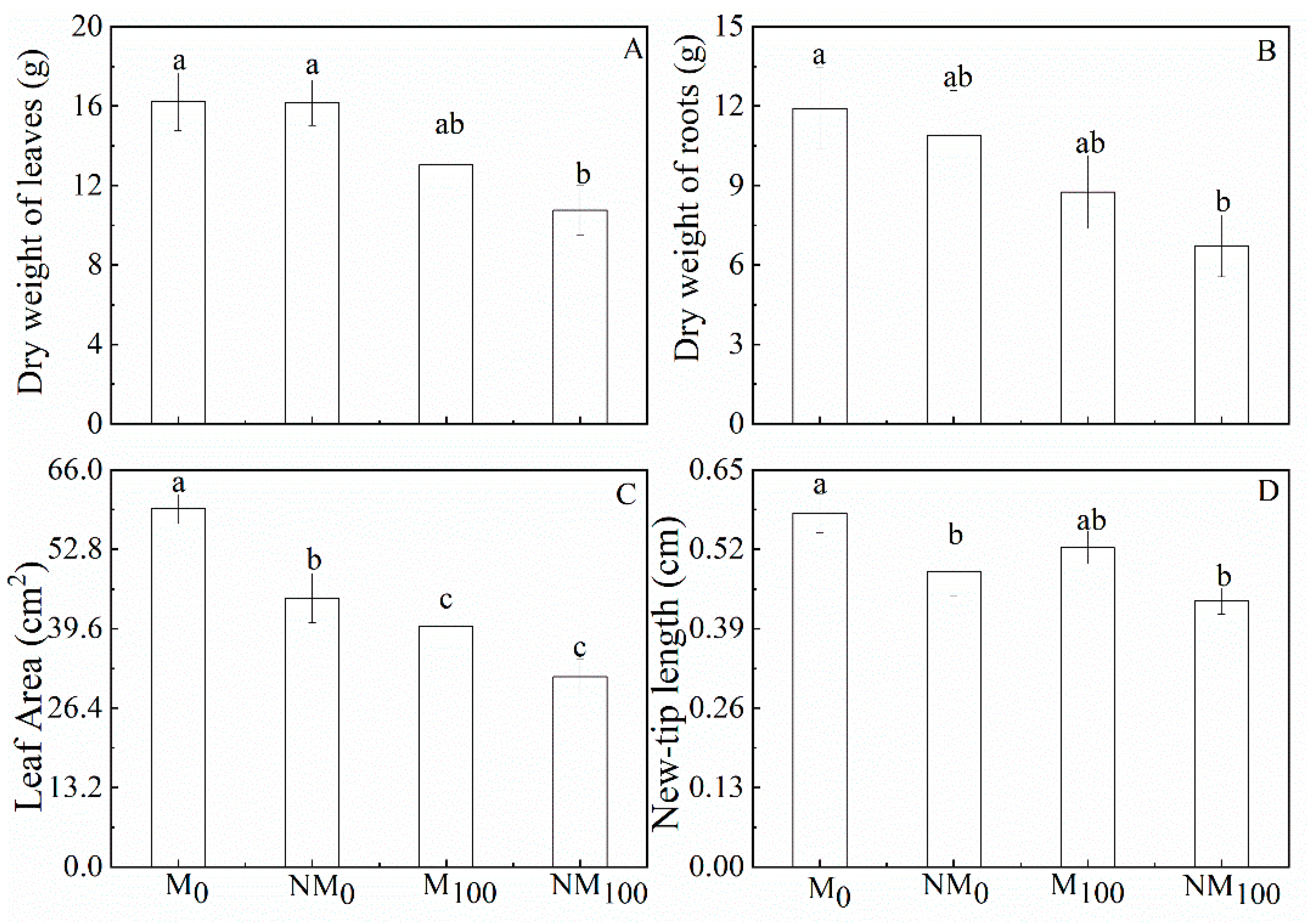
3.1. Plant Growth
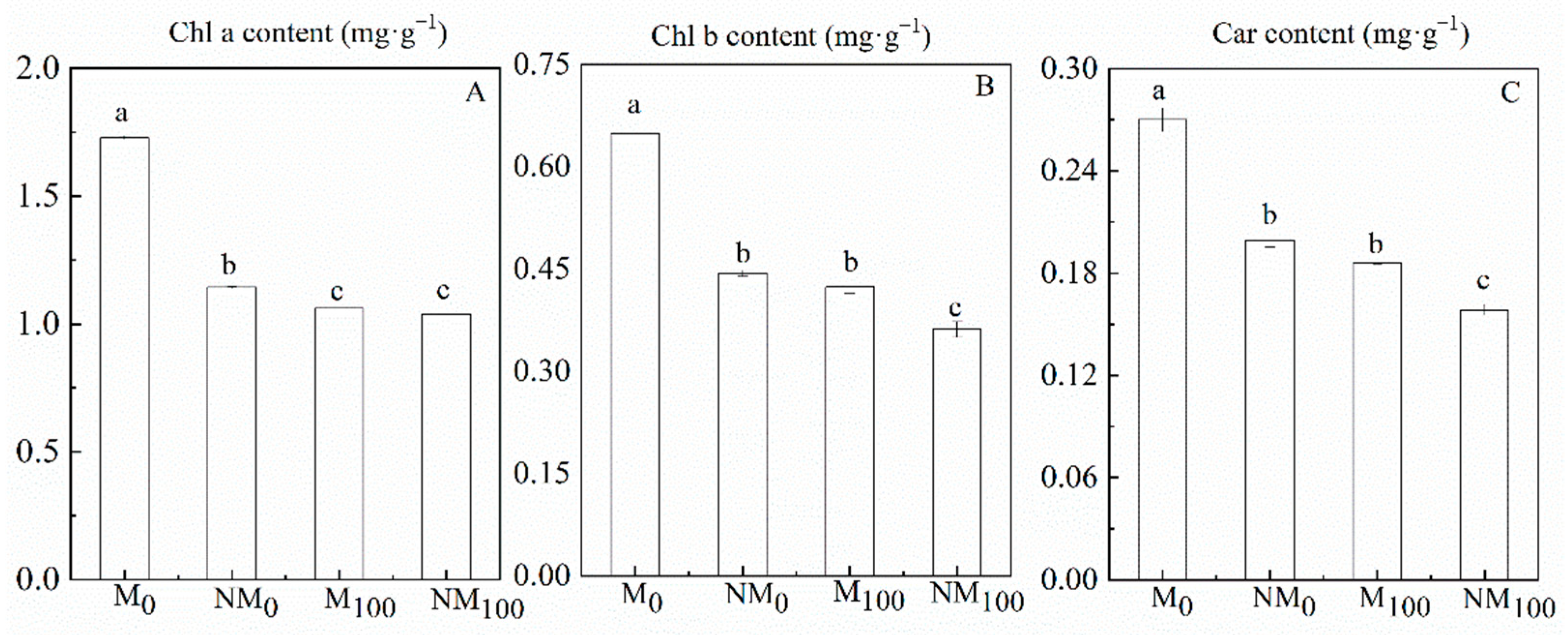
3.2. Photosynthetic Pigment Contents
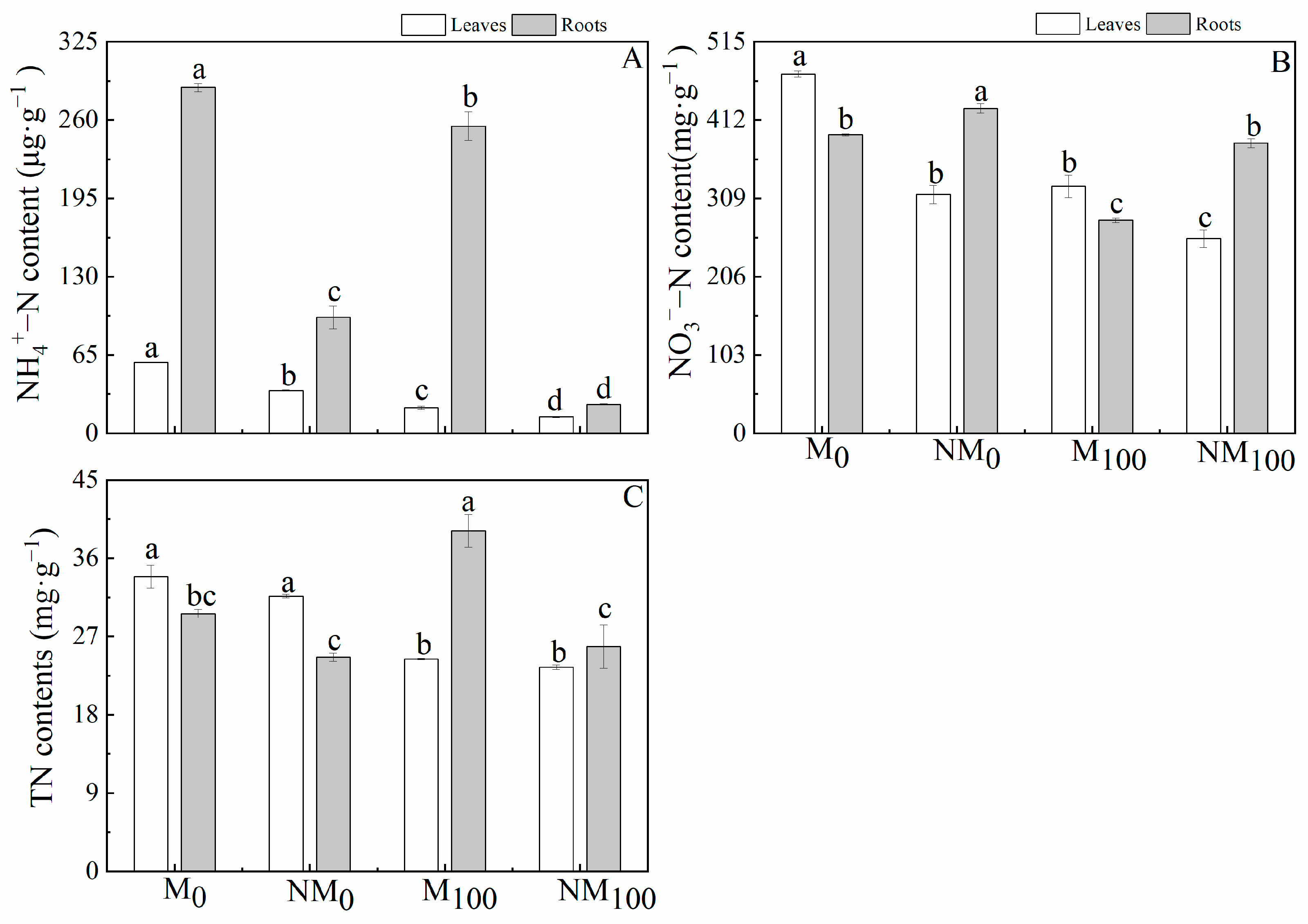
3.3. NH4+–N, NO3−–N, and TN Contents
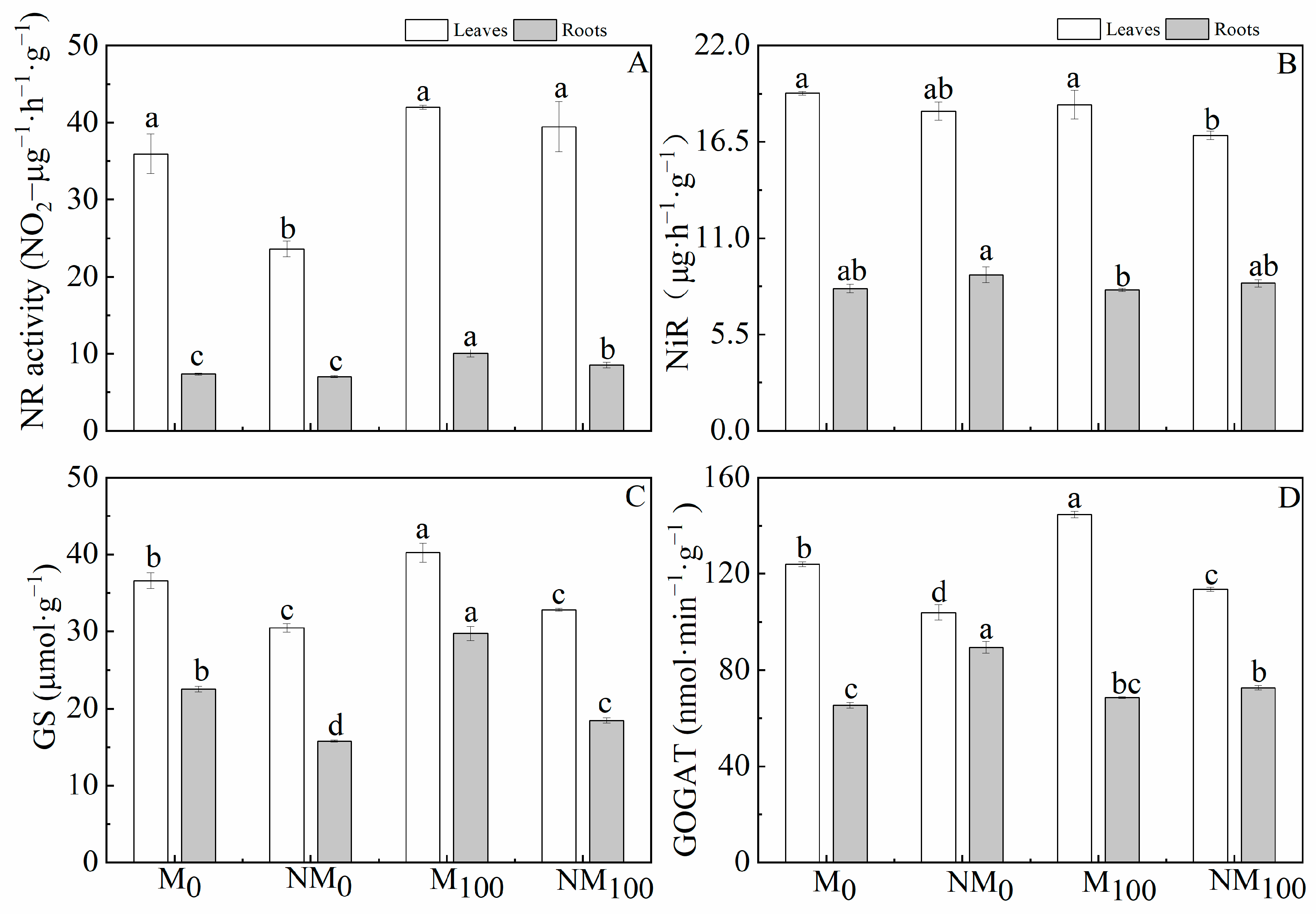
3.4. Activities of Key Enzymes in Nitrogen Metabolism
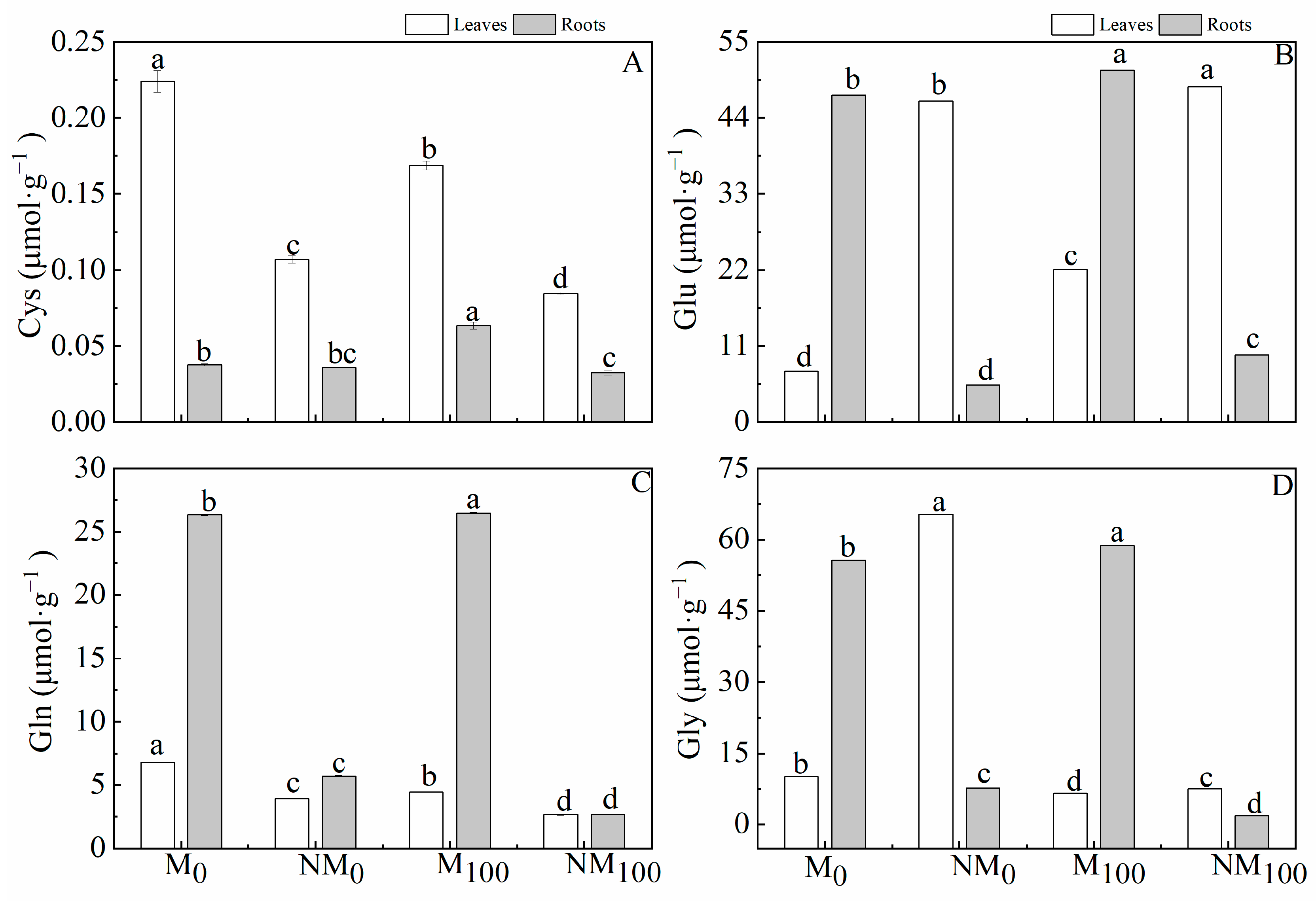
3.5. Free Amino Acid Contents
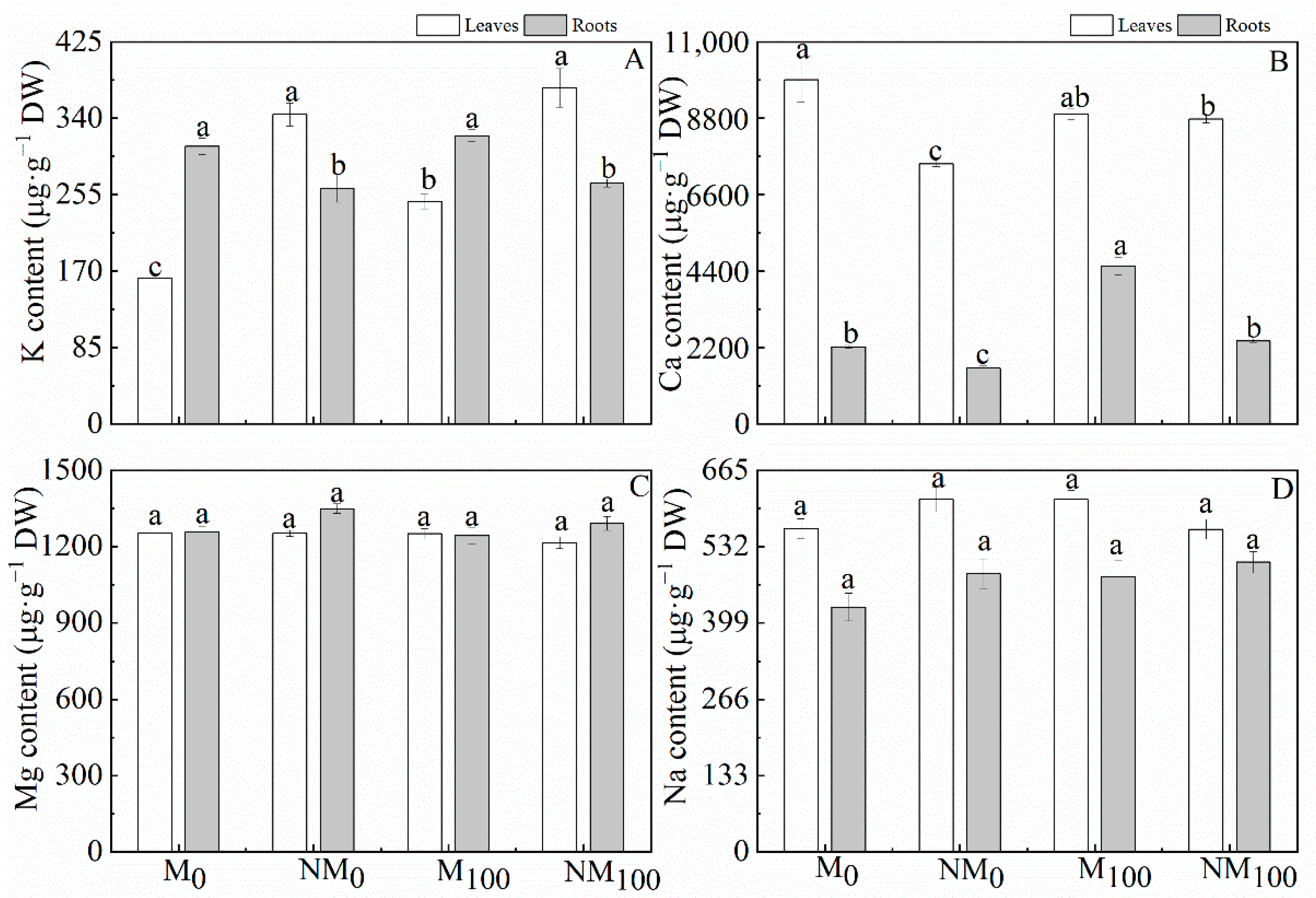
3.6. Contents of the Elements K, Ca, Na and Mg
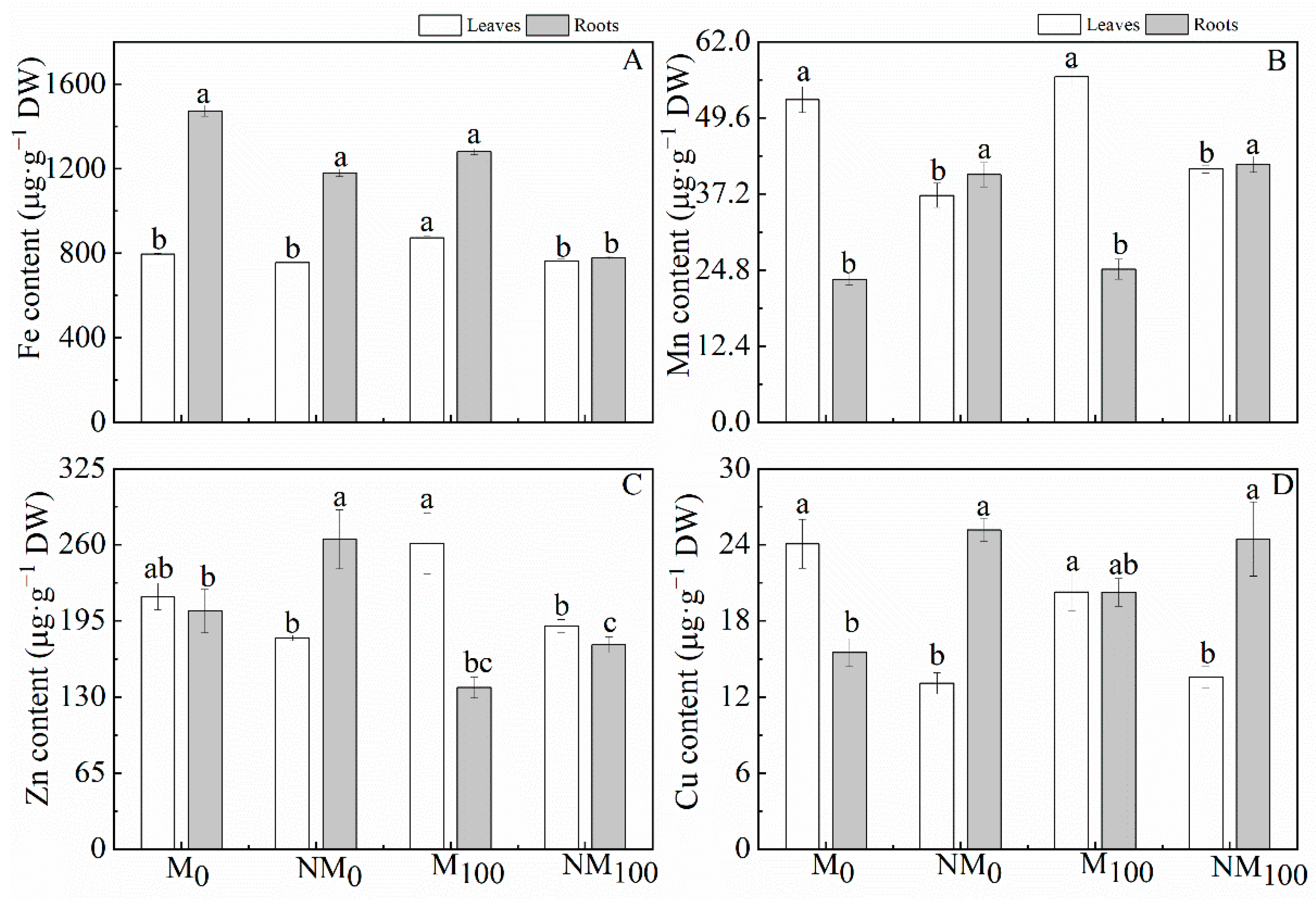
3.7. Trace Element Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fattahi, B.; Kazem, A.; Souri, M.K.; Barzegar, M. Morphophysiological and phytochemical responses to cadmium and lead stress in coriander (Coriandrum sativum L.). Ind. Crops Prod. 2021, 171, 113979. [Google Scholar] [CrossRef]
- Wani, A.S.; Tahir, I.; Ahmad, S.S.; Dar, R.A.; Nisar, S. Efficacy of 24-epibrassinolide in improving the nitrogen metabolism and antioxidant system in chickpea cultivars under cadmium and/or NaCl stress. Sci. Hortic. 2017, 225, 48–55. [Google Scholar] [CrossRef]
- Huybrechts, M.; Hendrix, S.; Kyndt, T.; Demeestere, K.; Vandamme, D.; Cuypers, A. Short-term effects of cadmium on leaf growth and nutrient transport in rice plants. Plant Sci. 2021, 313, 111054. [Google Scholar] [CrossRef] [PubMed]
- Sardar, R.; Ahmed, S.; Akbar, M.; Yasin, N.A.; Li, G. Alleviation of cadmium phytotoxicity in triacontanol treated Coriandrum sativum L. by modulation of physiochemical attributes, oxidative stress biomarkers and antioxidative system. Chemosphere 2022, 295, 133924. [Google Scholar] [CrossRef] [PubMed]
- Đukić-Ćosić, D.; Baralić, K.; Javorac, D.; Djordjevic, A.B.; Bulat, Z. An overview of molecular mechanisms in cadmium toxicity. Curr. Opin. Toxicol. 2020, 19, 56–62. [Google Scholar] [CrossRef]
- Hatamian, M.; Nejad, A.R.; Kafi, M.; Souri, M.K.; Shahbazi, K. Nitrate improves hackberry seedling growth under cadmium application. Heliyon 2020, 6, e03247. [Google Scholar] [CrossRef] [Green Version]
- Hatamian, M.; Nejad, A.R.; Kafi, M.; Souri, M.K.; Shahbazi, K. Interaction of lead and cadmium on growth and leaf morphophysiological characteristics of European hackberry (Celtis australis) seedlings. Chem. Biol. Technol. Agric. 2020, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Labidi, O.; Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M.; Sleimi, N. Assessing of growth, antioxidant enzymes, and phytohormones regulation in Cucurbita pepo under cadmium stress. Food Sci. Nutr. 2021, 9, 2021–2031. [Google Scholar] [CrossRef]
- Fidalgo, F.; Freitas, R.; Ferreira, R.; Pessoa, A.M.; Teixeira, J. Solanum nigrum L. antioxidant defence system isozymes are regulated transcriptionally and posttranslationally in Cd-induced stress. Environ. Exp. Bot. 2011, 72, 312–319. [Google Scholar] [CrossRef]
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cadmium: Toxicity and tolerance in plants. J. Environ. Biol. 2009, 30, 165–174. [Google Scholar]
- Hakeem, K.R.; Alharby, H.F.; Pirzadah, T.B. Exogenously applied calcium regulates antioxidative system and reduces cadmium-uptake in Fagopyrum esculentum. Plant Physiol. Bioch. 2022, 18, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.R.; Fang, Z.J.; Gao, J.; Wang, J.W.; Li, Y.B.; Sun, L.J.; Wang, Y.L.; Liao, J.M.; Gooneratne, R. Protective role of l-threonine against cadmium toxicity in Saccharomyces cerevisiae. J. Basic Microbiol. 2021, 61, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Javad, S.; Shah, A.A.; Ramzan, M.; Sardar, R.; Javed, T.; Al-Huqail, A.A.; Ali, H.M.; Chaudhry, O.; Yasin, N.A.; Ahmed, S.; et al. Hydrogen sulphide alleviates cadmium stress in Trigonella foenum-graecum by modulating antioxidant enzymes and polyamine content. Plant Biol. 2022, 24, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Nejad, A.R.; Hatamian, M.; Kafi, M.; Souri, M.K.; Shahbazi, K. Interaction of lead and nitrate on growth characteristics of ornamental judas tree (Cercis siliquastrum L.). Open Agric. 2018, 3, 670–677. [Google Scholar] [CrossRef]
- Souri, M.K.; Alipanahi, N.; Hatamian, M.; Ahmadi, M.; Tesfamariam, T. Elemental profile of heavy metals in garden cress, coriander, lettuce and spinach, commonly cultivated in Kahrizak, South of Tehran-Iran. Open Agric. 2018, 3, 3237. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.T.; Kao, C.H. Changes in protein and amino acid contents in two cultivars of rice seedlings with different apparent tolerance to cadmium. Plant Growth Regul. 2003, 40, 147–155. [Google Scholar] [CrossRef]
- Xu, J.; Wang, W.; Yin, H.; Liu, X.; Hong, S.; Mi, Q. Exogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots of Medicago truncatula seedlings under cadmium stress. Plant Soil 2010, 326, 321–330. [Google Scholar] [CrossRef]
- Silva, J.A.T.D.; Dobránszki, J. Impact of magnetic water on plant growth. Environ. Exp. Biol. 2014, 12, 137–142. [Google Scholar]
- Aguilar, C.H.; Dominguezpacheco, A.; Carballo, A.C.; Cruzorea, A.; Ivanov, R.; Bonilla, J.; Montanez, J. Alternating magnetic field irradiation effects on three genotype maize seed field performance. Acta Agrophysica 2009, 14, 7–17. [Google Scholar]
- Morejón, L.P.; Palacio, J.C.C.; Abad, L.V.; Govea, A.P. Stimulation of Pinus tropicalis M. seeds by magnetically treated water. Int. Agrophysica 2007, 21, 173–177. [Google Scholar]
- Afzal, I.; Saleem, S.; Skalicky, M.; Javed, T.; Muhammad, A.B.; Haq, Z.; Kamran, M.; Shahid, M.; Sohail, S.M.; Afzal, A.; et al. Magnetic field treatments improves sunflower yield by inducing physiological and biochemical modulations in seeds. Molecules 2021, 26, 2022. [Google Scholar] [CrossRef] [PubMed]
- Grewal, H.S.; Maheshwari, B.L. Magnetic treatment of irrigation water and snow pea and chickpea seeds enhances early growth and nutrient contents of seedlings. Bioelectromagnetics 2010, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.N.; Li, Y.; Yang, Y.Q.; Li, C.Y. Effect of seed pretreatment by magnetic field on the sensitivity of cucumber (Cucumis sativus) seedlings to ultraviolet-B radiation. Environ. Exp. Bot. 2004, 54, 286–294. [Google Scholar] [CrossRef]
- Kandeler, E.; Eder, G.; Sobotik, M. Microbial biomass, N mineralization, and the activities of various enzymes in relation to nitrate leaching and root distribution in a slurry-amended grassland. Biol. Fert. Soils 1994, 18, 7–12. [Google Scholar] [CrossRef]
- Khoshravesh-Miangoleh, M.; Kiani, A.R. Effect of magnetized water on infiltration capacity of different soil textures. Soil Use Manag. 2015, 30, 588–594. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, T.T.; Huang, X.Y.; Hou, Y.S.; Huang, H.Z. Magnetic water improve the tolerance of corn to Cd ion. J. Xiamen Univ. (Nat. Sci.) 2008, 47, 278–281. [Google Scholar]
- Chen, Y.P.; Li, R.; He, J.M. Magnetic field can alleviate toxicological effect induced by cadmium in mungbean seedlings. Ecotoxicology 2011, 20, 760–769. [Google Scholar] [CrossRef]
- Zhao, S.J.; Cang, J. Experimental Guidance for Plant Physiology; China Agriculture Press: Beijing, China, 2016. [Google Scholar]
- Xu, W.A. Comprehensive Experimental Guidance of Biology; Soochow University Press: Suzhou, China, 2010. [Google Scholar]
- Bergareche, C.; Ayuso, R.; Masgrau, C.; Simon, E. Nitrate reductase in cotyledons of cucumber seedlings as affected by nitrate, phytochrome and calcium. Physiol. Plant. 2010, 91, 257–262. [Google Scholar] [CrossRef]
- Barbosa, J.M.; Singh, N.K.; Cherry, J.H.; Locy, R.D. Nitrate uptake and utilization is modulated by exogenous gamma-aminobutyric acid in Arabidopsis thaliana seedlings. Plant Physiol. Biochem. 2010, 48, 443–450. [Google Scholar] [CrossRef]
- He, Y.; Chen, Y.F.; Zhu, Y.P.; Lu, J.; Lin, R.C. HPLC determination of the main free amino acids in root of isatis indigotica fort. Chin. J. Pharm. Anal. 2005, 25, 274–277. [Google Scholar]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen modifies NaCl toxicity in eggplant seedlings: Assessment of chlorophyll a fluorescence, antioxidative response and proline metabolism. Biocatal. Agric. Biotechnol. 2016, 7, 76–86. [Google Scholar] [CrossRef]
- Ahmad, P.; Latef, A.A.A.; Hashem, A.; Abd_Allah, E.F.; Tran, L.S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataria, S.; Baghel, L.; Guruprasad, K. Alleviation of adverse effects of ambient UV stress on growth and some potential physiological attributes in soybean (Glycine max) by seed pre-treatment with static magnetic field. J. Plant Growth Regul. 2017, 36, 550–565. [Google Scholar] [CrossRef]
- Baghel, L.; Kataria, S.; Guruprasad, K.N. Static magnetic field treatment of seeds improves carbon and nitrogen metabolism under salinity stress in soybean. Bioelectromagnetics 2016, 37, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shang, Y.K.; Li, L.; Chen, Y.H.; Qin, Z.Z. Cadmium stress in Dongying wild soybean seedlings: Growth, Cd accumulation, and photosynthesis. Photosynthetica 2018, 56, 1346–1352. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2017, 255, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C.A.; Sioulas, C. Safflower yield, chlorophyll content, photosynthesis, and water use efficiency response to nitrogen fertilization under rainfed conditions. Ind. Crops Prod. 2008, 27, 75–85. [Google Scholar] [CrossRef]
- Namdjoyan, S.; Kermanian, H. Exogenous nitric oxide (as sodium nitroprusside) ameliorates arsenic-induced oxidative stress in watercress (Nasturtium officinale R. Br.) Plants. Sci. Hortic. 2013, 161, 350–356. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Z.; Zhu, C. Endogenous nitric oxide mediates alleviation of cadmium toxicity induced by calcium in rice seedlings. J. Environ. Sci. 2012, 24, 940–948. [Google Scholar] [CrossRef]
- Ci, D.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Cadmium stress in wheat seedlings: Growth, cadmium accumulation and photosynthesis. Acta Physiol. Plant 2010, 32, 365–373. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, G.; Arora, K.; Kohli, R.K. Nitric oxide (as sodium nitroprusside) supplementation ameliorates Cd toxicity in hydroponically grown wheat roots. Environ. Exp. Bot. 2008, 63, 158–167. [Google Scholar] [CrossRef]
- Xu, J.; Yin, H.X.; Li, X. Protective effects of proline against cadmium toxicity in micropropagated hyperaccumulator, Solanum nigrum L. Plant Cell Rep. 2009, 28, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Broadley, M.R.; Escobar-Gutiérrez, A.; Burns, A.; Burns, I.G. What are the effects of nitrogen deficiency on growth components of lettuce? New Phytol. 2010, 147, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Kataria, S.; Jain, M.; Rastogi, A.; Brestic, M. Static magnetic field treatment enhanced photosynthetic performance in soybean under supplement ultraviolet-B radiation. Photosynth. Res. 2021, 150, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, M.; Canal, A.D.; Quaggiotti, S.; Pegoraro, P.; Bottacin, A. Differences in nitrate and ammonium uptake between Scots pine and European larch. Plant Soil 2000, 221, 1–3. [Google Scholar] [CrossRef]
- Gajewska, E.; SkOdowska, M. Nickel-induced changes in nitrogen metabolism in wheat shoots. J. Plant Physiol. 2009, 166, 1034–1044. [Google Scholar] [CrossRef]
- Peoples, M.B.; Gifford, R.M. Regulation of the transport of nitrogen and carbonin higher plants. In Plant Metabolism; Dennis, D.T., Turpin, D.H., Lefebvre, D.D., Eds.; Longman Singapore Publishers (Pte) Ltd.: London, UK, 1997; pp. 525–537. [Google Scholar]
- Hernández, L.E.; Gárate, A.; Carpena-Ruiz, R. Effects of cadmium on the uptake, distribution and assimilation of nitrate in Pisum sativum. Plant Soil 1997, 189, 97–106. [Google Scholar] [CrossRef]
- Ghnaya, T.; Nouairi, I.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C.; Ghorbel, M.H. Cadmium effects on growth and mineral nutrition of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef]
- González, A.; Lynch, S.J.P. Light and excess manganese: Implications for oxidative stress in common bean. Plant Physiol. 1998, 118, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Dučić, T.; Polle, A. Transport and detoxification of manganese and copper in plants. Braz. J. Plant Physiol. 2005, 17, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Sarraf, M.; Deamici, K.M.; Taimourya, H.; Islam, M.; Kataria, S.; Raipuria, R.K.; Abdi, G.; Brestic, M. Effect of magnetopriming on photosynthetic performance of plants. Int. J. Mol. Sci. 2021, 22, 9353. [Google Scholar] [CrossRef] [PubMed]
- Duy, D.; Wanner, G.; Meda, A.R.; Von Wiren, N.; Soll, J.; Philippar, K. PIC1, an ancient permease in Arabidopsis chloroplasts, mediates iron transport. Plant Cell 2007, 19, 986–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellingen, K.; Dominique, V.; Remans, T.; Vangronsveld, J.; Keunen, E.; Cuypers, A. Ethylene signalling is mediating the early cadmium-induced oxidative challenge in Arabidopsis thaliana. Plant Sci. 2015, 239, 137–146. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Anna, A. K+ Nutrition and Na+ Toxicity: The Basis of Cellular K+/Na+ Ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wang, L.; Ma, F.; Guo, J.; Zhu, H.; Meng, S.; Bi, S.; Wang, H. Magnetic Treatment Improves the Seedling Growth, Nitrogen Metabolism, and Mineral Nutrient Contents in Populus × euramericana ‘Neva’ under Cadmium Stress. Forests 2022, 13, 947. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060947
Liu X, Wang L, Ma F, Guo J, Zhu H, Meng S, Bi S, Wang H. Magnetic Treatment Improves the Seedling Growth, Nitrogen Metabolism, and Mineral Nutrient Contents in Populus × euramericana ‘Neva’ under Cadmium Stress. Forests. 2022; 13(6):947. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060947
Chicago/Turabian StyleLiu, Xiumei, Lu Wang, Fengyun Ma, Jianyao Guo, Hong Zhu, Shiyuan Meng, Sisheng Bi, and Huatian Wang. 2022. "Magnetic Treatment Improves the Seedling Growth, Nitrogen Metabolism, and Mineral Nutrient Contents in Populus × euramericana ‘Neva’ under Cadmium Stress" Forests 13, no. 6: 947. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060947