
Distribution Pattern of Woody Plants in a Mountain Forest Ecosystem Influenced by Topography and Monsoons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling Design and Data Collection
2.3. Data Analysis
3. Results
3.1. Species Composition of Two Habitats
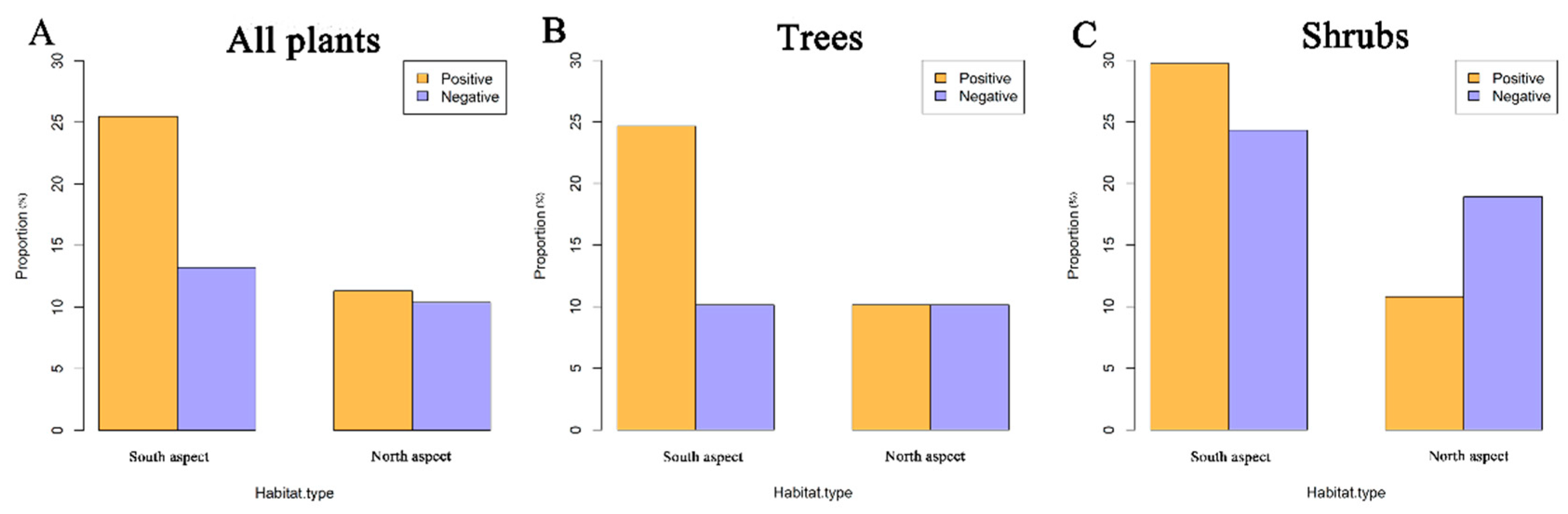
3.2. Associations between Species and Habitats
3.3. Co-Occurrence Network Analysis of Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Haugen, J.E.; Xu, C.Y. Precipitation pattern in the Western Himalayas revealed by four datasets. Hydrol. Earth Syst. Sci. 2018, 22, 5097–5110. [Google Scholar] [CrossRef] [Green Version]
- Chian, Y.S.; Yeh, C.L.; Wang, C.C. Variation in mountain vegetation composition between the east and the west sides of southern Taiwan. Forests 2016, 7, 179. [Google Scholar] [CrossRef] [Green Version]
- Bradstock, R.A.; Hammill, K.A.; Collins, L.; Price, O. Effects of weather, fuel and terrain on fire severity in topographically diverse landscapes of south-eastern Australia. Landsc. Ecol. 2010, 25, 607–619. [Google Scholar] [CrossRef]
- Erfanzadeh, R.; Omidipour, R.; Faramarzi, M. Variation of plant diversity components in different scales in relation to grazing and climatic conditions. Plant Ecol. Divers. 2015, 8, 537–545. [Google Scholar] [CrossRef]
- Sporbert, M.; Welk, E.; Seidler, G.; Jandt, U.; Aćić, S.; Biurrun, I.; Campos, J.A.; Čarni, A.; Cerabolini, B.E.L.; Chytrý, M.; et al. Different sets of traits explain abundance and distribution patterns of European plants at different spatial scales. J. Veg. Sci. 2021, 32, e13016. [Google Scholar] [CrossRef]
- Kowarik, I.; Lipp, M.; Cierjacks, A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales. Preslia 2013, 85, 113–132. [Google Scholar]
- Huang, E.; Chen, Y.; Fang, M.; Zheng, Y.; Yu, S. Environmental drivers of plant distributions at global and regional scales. Glob. Ecol. Biogeogr. 2021, 30, 697–709. [Google Scholar] [CrossRef]
- Opedal, Ø.H.; Armbruster, W.S.; Graae, B.J. Linking small-scale topography with microclimate, plant species diversity and intra-specific trait variation in an alpine landscape. Plant Ecol. Divers. 2015, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hu, G.; Ni, J. Effects of topographical and edaphic factors on the distribution of plant communities in two subtropical karst forests, southwestern China. J. Mt. Sci. 2013, 10, 95–104. [Google Scholar] [CrossRef]
- Liu, Q.; Bi, L.; Song, G.; Wang, Q.; Jin, G. Species–habitat associations in an old-growth temperate forest in northeastern China. BMC Ecol. 2018, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Ozinga, W.A.; Schaminée, J.H.J.; Bekker, R.M.; Bonn, S.; Poschlod, P.; Tackenberg, O.; Bakker, J.; Van Groenendael, J.M. Predictability of plant species composition from environmental conditions is constrained by dispersal limitation. Oikos 2005, 108, 555–561. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, Y.; Jia, H.; Sun, W.; Ming, A.; Pang, S.; An, N.; Zhang, J.; Tang, C.; Dong, S. Influence of slope direction on the soil seed bank and seedling regeneration of Castanopsis hystrix seed rain. Forests 2021, 12, 500. [Google Scholar] [CrossRef]
- Han, T.D.; Ding, Y.J.; Ye, B.S.; Xie, C.W. Precipitation Variations on the Southern and Northern Slopesof the Tianger Range in Tianshan Mountains. J. Glaciol. Geocryol. 2004, 26, 761–766. [Google Scholar]
- Frazier, A.G.; Giambelluca, T.W. Spatial trend analysis of Hawaiian rainfall from 1920 to 2012. Int. J. Climatol. 2017, 37, 2522–2531. [Google Scholar] [CrossRef]
- Ren, W.; Yao, T.; Xie, S. Key drivers controlling the stable isotopes in precipitation on the leeward side of the central Himalayas. Atmos. Res. 2017, 189, 134–140. [Google Scholar] [CrossRef]
- Bunce, J.A.; Chabot, B.F.; Miller, L.N. Role of annual leaf carbon balance in the distribution of plant species along an elevational gradient. Bot. Gaz. 1979, 140, 288–294. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.C.G.; Thomaz, S.M.; Bini, L.M. Experiments reveal that environmental heterogeneity increases species richness, but they are rarely designed to detect the underlying mechanisms. Oecologia 2018, 188, 11–22. [Google Scholar] [CrossRef]
- Zhirnova, D.F.; Belokopytova, L.V.; Barabantsova, A.E.; Babushkina, E.A.; Vaganov, E.A. What prevails in climatic response of Pinus sylvestris in-between its range limits in mountains: Slope aspect or elevation? Int. J. Biometeorol. 2020, 64, 333–344. [Google Scholar] [CrossRef]
- Suggitt, A.J.; Wilson, R.J.; Isaac, N.J.B.; Beale, C.M.; Auffret, A.G.; August, T.; Bennie, J.J.; Crick, H.Q.P.; Duffield, S.; Fox, R.; et al. Extinction risk from climate change is reduced by microclimatic buffering. Nat. Clim. Chang. 2018, 8, 713–717. [Google Scholar] [CrossRef]
- Li, X.; Zhang, B.; Ren, R.; Li, L.; Simonovic, S.P. Spatio-Temporal Heterogeneity of Climate Warming in the Chinese Tianshan Mountainous Region. Water 2022, 14, 199. [Google Scholar] [CrossRef]
- Sigua, G.C.; Coleman, S.W. Spatial distribution of soil carbon in pastures with cow-calf operation: Effects of slope aspect and slope position. J. Soils Sediments 2010, 10, 240–247. [Google Scholar] [CrossRef]
- Zhang, X.; Adamowski, J.F.; Liu, C.; Zhou, J.; Zhu, G.; Dong, X.; Cao, J.; Feng, Q. Which slope aspect and gradient provides the best afforestation-driven soil carbon sequestration on the China’s Loess Plateau? Ecol. Eng. 2020, 147, 105782. [Google Scholar] [CrossRef]
- Bangroo, S.A.; Najar, G.R.; Rasool, A. Effect of altitude and aspect on soil organic carbon and nitrogen stocks in the Himalayan Mawer Forest Range. Catena 2017, 158, 63–68. [Google Scholar] [CrossRef]
- Lindroth, A.; Grelle, A.; Moren, A. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity. Glob. Chang. Biol. 1998, 4, 443–450. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Zhu, L.; Guo, L. Study on change of northern subtropical border in mountainous regions in western Henan Province. J. Henan Univ. Nat. Sci. 2016, 46, 40–49. [Google Scholar] [CrossRef]
- Hao, C.; Zhang, H.; Zhou, J. Comparative Analysis on Soil Basic Properties Between Southern and Northern Slopes in Funiu Mountain, China. Open Electr. Electron. Eng. J. 2014, 8, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, Z.; Chen, M.; Zhang, Q.; Yang, F. Research on the population structure and dynamics of the natural Taxus chinensis in Funiushan Mountain area of Henan Province. Res. Soil Water Conserv. 2016, 23, 262–268. [Google Scholar] [CrossRef]
- Chen, S.; Hai, X.; Ye, Y.; Yuan, Z.; Yao, S.; Wang, T. Contrastive analysis of soil nutrients in coniferous and broad-leaved mixed forest on southern and northern slope of Funiu mountain. J. Cent. South Univ. For. Technol. 2018, 38, 65–71. [Google Scholar] [CrossRef]
- Shao-hui, W.; Meng-na, L.; Xu, W.; Ya-chen, Z.; Yi-wen, H.; Ce, Z.; Tong-xiang, L. Protection and Quality Control of Wild Chinese Herbal Medicine Resources in Nature Reserves by Taking Longyuwan Scenic Spot Within Funiu Mountain Nature Reserve as an Example. J. Northeast Agric. Univ. 2016, 23, 90–96. [Google Scholar] [CrossRef]
- Peng, J.; Li, J.; Wang, T.; Huo, J.; Yang, L. Effect of altitude on climate–growth relationships of Chinese white pine (Pinus armandii) in the northern Funiu Mountain, central China. Clim. Chang. 2019, 154, 273–288. [Google Scholar] [CrossRef]
- Zhu, W.; Li, S. The dynamic response of forest vegetation to hydrothermal conditions in the Funiu Mountains of western Henan Province. J. Geogr. Sci. 2017, 27, 565–578. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Liu, H.; Hu, N.; Ding, S. Photosynthetic characteristics of plant functional groups in forest ecosystem at the national natural reserve of FuNiu Mountain. Acta Ecol. Sin. 2016, 36, 4609–4616. [Google Scholar]
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Chen, Y.; Shao, Y.; Xi, J.; Yuan, Z.; Ye, Y.; Wang, T. Community preferences of woody plant species in a heterogeneous temperate forest, China. Front. Ecol. Evol. 2020, 8, 165. [Google Scholar] [CrossRef]
- Xi, J.; Shao, Y.; Li, Z.; Zhao, P.; Ye, Y.; Li, W.; Chen, Y.; Yuan, Z. Distribution of woody plant species among different disturbance regimes of forests in a temperate deciduous broad-leaved forest. Front. Plant Sci. 2021, 12, 618524. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 1 March 2021).
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. Proc. Int. AAAI Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar]
- Heegaard, E.; Boddy, L.; Diez, J.M.; Halvorsen, R.; Kauserud, H.; Kuyper, T.W.; Bässler, C.; Büntgen, U.; Gange, A.C.; Krisai-Greilhuber, I.; et al. Fine-scale spatiotemporal dynamics of fungal fruiting: Prevalence, amplitude, range and continuity. Ecography 2017, 40, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Torrecillas, E.; del Mar Alguacil, M.; Roldán, A.; Díaz, G.; Montesinos-Navarro, A.; Torres, M.P. Modularity reveals the tendency of arbuscular mycorrhizal fungi to interact differently with generalist and specialist plant species in gypsum soils. Appl. Environ. Microbiol. 2014, 80, 5457–5466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicks, L.V.; Corbet, S.A.; Pywell, R.F. Compartmentalization in plant-insect flower visitor webs. J. Anim. Ecol. 2002, 71, 32–43. [Google Scholar] [CrossRef]
- Wirta, H.K.; Hebert, P.D.N.; Kaartinen, R.; Prosser, S.W.; Várkonyi, G.; Roslin, T. Complementary molecular information changes our perception of food web structure. Proc. Natl. Acad. Sci. USA 2014, 111, 1885–1890. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Zhou, J. Species diversity of south and north slopes of Fu’niu Mountain and its correlation with main ecological factors. J. Plant Resour. Environ. 2013, 2, 38–44. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, X.; Jia, L.M. Establishment of site index table for Quercus variabilis natural secondary forest in Funiushan Mountainous area. J. Northeast. For. Univ. 2017, 45, 32–37. [Google Scholar] [CrossRef]
- Yang, X.F.; Wang, T.X.; Zhang, J.Y.; Li, F.Q. The Pteridophytic Flora of Funiu Mountains in Henan Province. J. Henan Norm. Univ. 1998, 26, 65–67. [Google Scholar] [CrossRef]
- Chen, Z.C.; Hao, C.y.; Liu, C.H.; Ding, X.D. Influences of barrier function on the temporal and spatial characteristics of temperature, precipitation, land cover and net primary productivity in Mt. Funiu. Ecol. Environ. Sci. 2013, 22, 761–766. [Google Scholar] [CrossRef]
- Auslander, M.; Nevo, E.; Inbar, M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. J. Arid Environ. 2003, 55, 405–416. [Google Scholar] [CrossRef]
- Warren, R.J. Mechanisms driving understory evergreen herb distributions across slope aspects: As derived from landscape position. Plant Ecol. 2008, 198, 297–308. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Hédl, R.; Chudomelová, M.; McCulley, R.L.; Nelson, J.A. Variation in vegetation and microbial linkages with slope aspect in a montane temperate hardwood forest. Ecosphere 2014, 5, 1–17. [Google Scholar] [CrossRef]
- Warren, R.J.; Ursell, T.; Keiser, A.D.; Bardford, M.A. Habitat, dispersal and propagule pressure control exotic plant infilling within an invaded range. Ecosphere 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Eilu, G.; Obua, J. Tree condition and natural regeneration in disturbed sites of Bwindi Impenetrable Forest National Park, southwestern Uganda. Trop. Ecol. 2005, 46, 99–112. [Google Scholar]
- Punchi-Manage, R.; Getzin, S.; Wiegand, T.; Kanagaraj, R.; Savitri Gunatilleke, C.V.; Nimal Gunatilleke, I.A.U.; Wiegand, K.; Huth, A. Effects of topography on structuring local species assemblages in a S ri L ankan mixed dipterocarp forest. J. Ecol. 2013, 101, 149–160. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, T.; Zang, D.; Dong, Y.; Dai, Y. Analysis on community Composition and Structure of Lindera obtusiloba in Laoshan. J. Agric. 2014, 4, 60–63. [Google Scholar]
- Li, M.; Lv, G.; Niu, Y.; Meng, Z.; Yang, X. Ectomycorrhizal community structure and indiving factors of Betula platyphylla in different climatic zones in Inner Mongolia. Acta Ecol. Sin. 2022, 1–14. [Google Scholar]
- Li, H.D.; Wu, X.W.; Xiao, Z.S. Assembly, ecosystem functions and stability in species interaction networks. Chin. J. Plant Ecol. 2021, 45, 1049–1063. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional molecular ecological networks. MBio 2010, 1, e00169-10. [Google Scholar] [CrossRef] [Green Version]
- Hai, X. Response of Radial Growth of Pinus tabulaeformis to Climate Change and Temperature Reconstruction in Funiu Mountains; Henan Agricultural University: Zhengzhou, China, 2020. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, W.; Zhu, L.; Li, Y. Multi-scale analysis of trade-off/synergy effects of forest ecosystem services in the Funiu Mountain Region. Acta Geogr. Sin. 2020, 75, 05000975. [Google Scholar] [CrossRef]
- Tripathee, L.; Kang, S.; Huang, J.; Sillanpää, M.; Sharma, C.M.; Lüthi, Z.L.; Guo, J.; Paudyal, R. Ionic composition of wet precipitation over the southern slope of central Himalayas, Nepal. Environ. Sci. Pollut. Res. 2014, 21, 2677–2687. [Google Scholar] [CrossRef]
- Zhang, X.; Xing, Y.J.; Yan, G.Y.; Wang, Q.G. Response of fine roots to precipitation change: A meta-analysis. Chin. J. Plant Ecol. 2018, 42, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.L.; Dai, X.Q.; Wang, H.M.; Fu, X.L.; Kou, L. Rhizosphere effects of overstory tree and understory shrub species in central subtropical plantations—A case study at Qianyanzhou, Taihe, Jiangxi, China. Chin. J. Plant Ecol. 2018, 42, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Pan, L.; Lei, J.; Tang, W.; Pang, H.; Cui, H.; Wang, X. Effects of Forest Stand Structure Characteristics on Shrub Species Diversity in Pinus massoniana Forest in Three Gorges Reservoir Area. Ecol. Environ. 2019, 28, 1332. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Wang, Z.; Liu, W.; Fu, Q.; Shao, Y.; Liu, F.; Ye, Y.; Chen, Y.; Yuan, Z. Distribution Pattern of Woody Plants in a Mountain Forest Ecosystem Influenced by Topography and Monsoons. Forests 2022, 13, 957. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060957
Zhou X, Wang Z, Liu W, Fu Q, Shao Y, Liu F, Ye Y, Chen Y, Yuan Z. Distribution Pattern of Woody Plants in a Mountain Forest Ecosystem Influenced by Topography and Monsoons. Forests. 2022; 13(6):957. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060957
Chicago/Turabian StyleZhou, Xiayan, Zhao Wang, Wenxin Liu, Qianjin Fu, Yizhen Shao, Fengqin Liu, Yongzhong Ye, Yun Chen, and Zhiliang Yuan. 2022. "Distribution Pattern of Woody Plants in a Mountain Forest Ecosystem Influenced by Topography and Monsoons" Forests 13, no. 6: 957. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060957