
Changes in the Abundance of Monoterpenes from Breathable Air of a Mediterranean Conifer Forest: When Is the Best Time for a Human Healthy Leisure Activity?

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
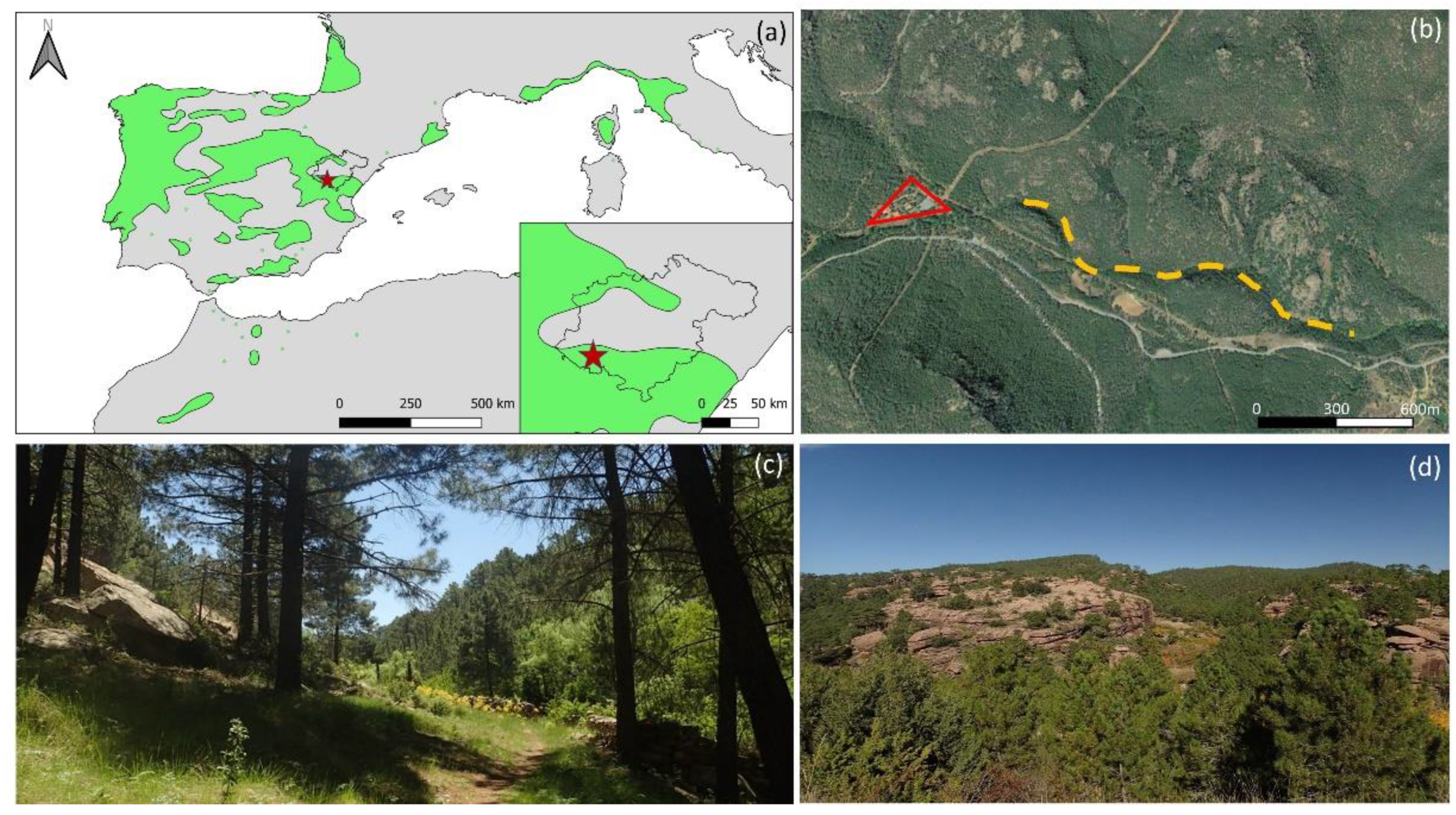
2.1. Experimental Site
2.2. Environmental Factors and Phenology
2.3. Monoterpenes Sampling
2.4. Statistical Analysis
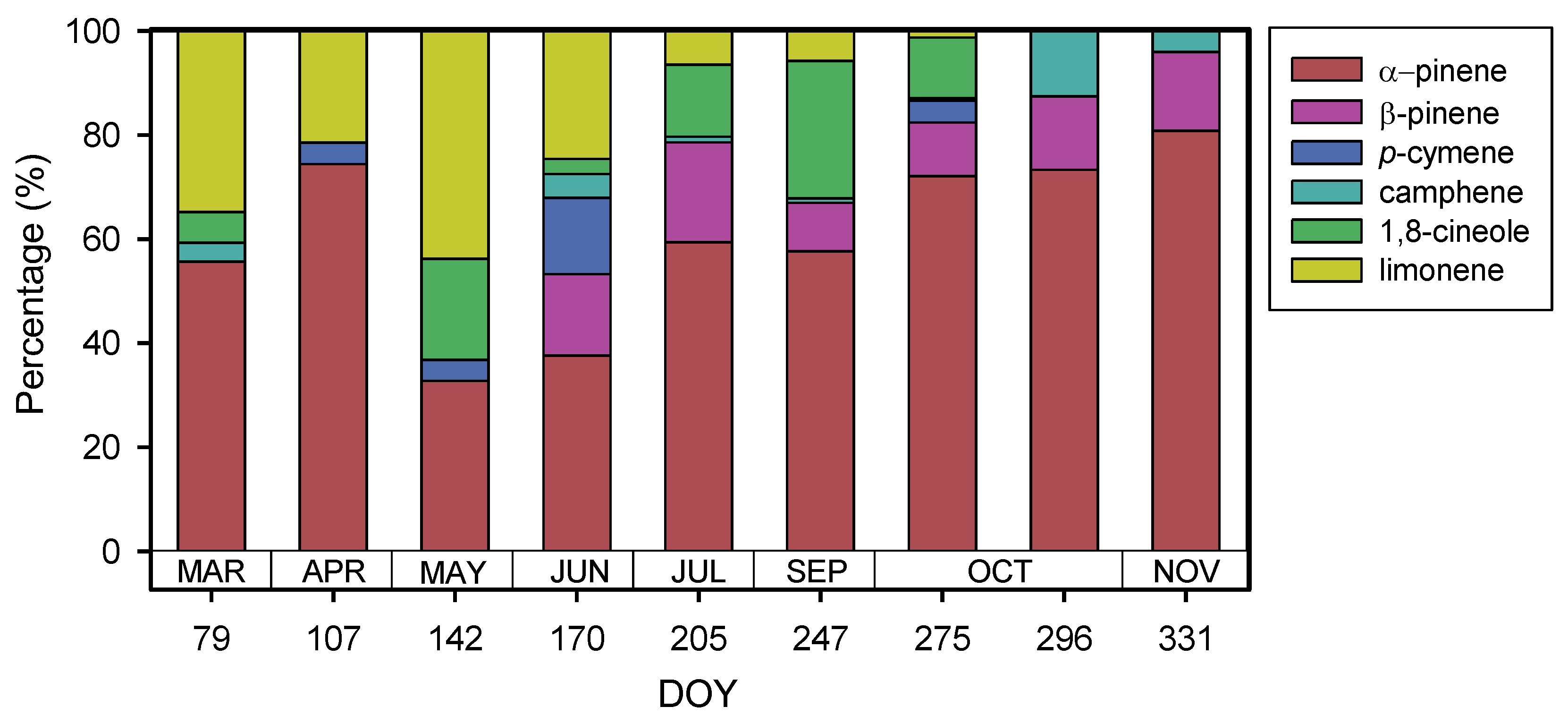
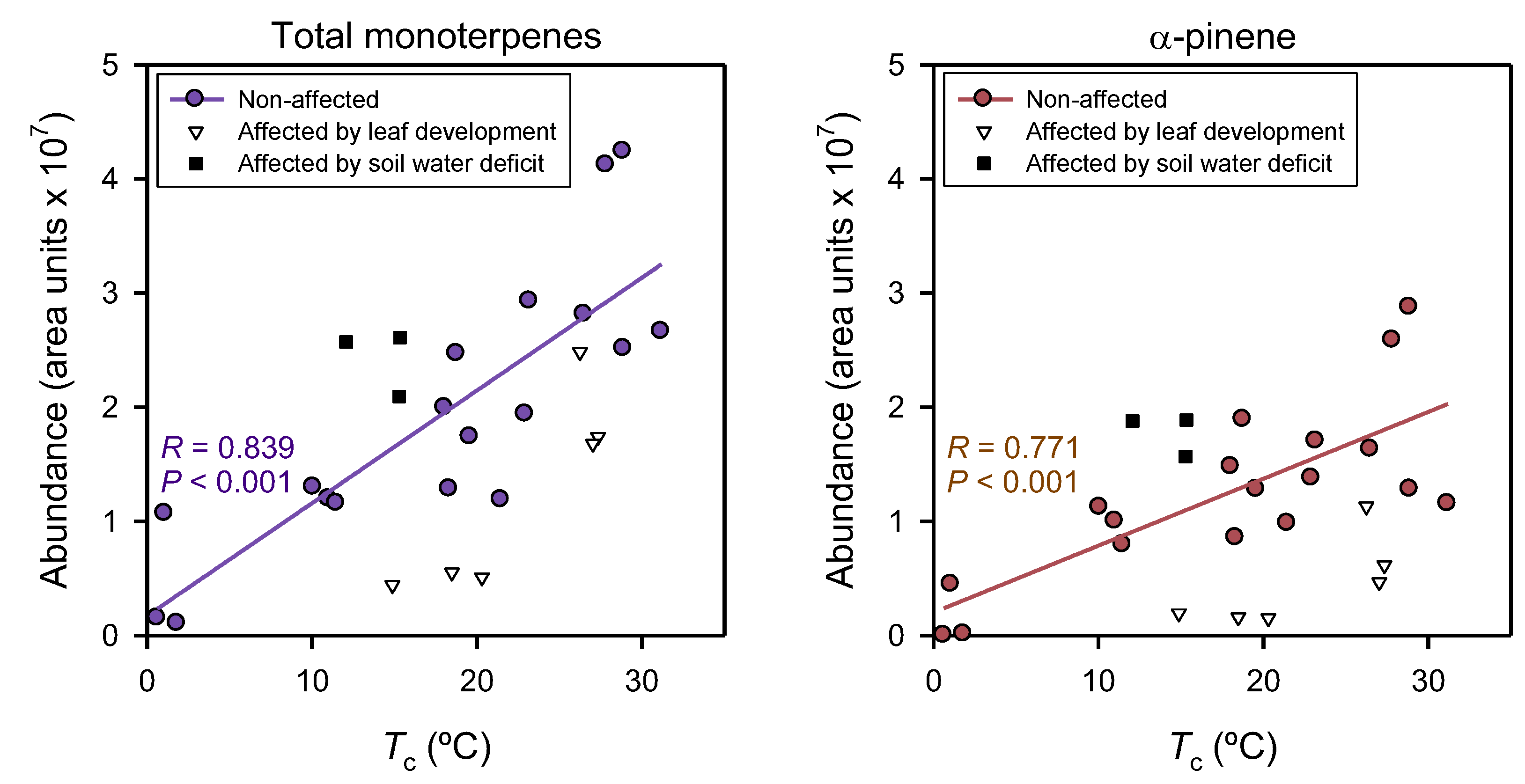
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barreiro-Hurle, J.; Gracia, A.; de-Magistris, T. The effects of multiple health and nutrition labels on consumer food choices. J. Agric. Econ. 2010, 61, 426–443. [Google Scholar] [CrossRef]
- Leelapattana, W.; Hsu, S.Y.; Thongma, W.; Chen, C.; Chiang, F.M. Understanding the impact of environmental education on tourists’ future visit intentions to leisure farms in mountain regions. Sustainability 2019, 11, 1567. [Google Scholar] [CrossRef] [Green Version]
- Ceylan, E.N.; Tülbentçi, T. Example of an ecotourism farm in the context of sustainability: Pastoral valley ecological life farm. Int. J. Adv. Appl. 2020, 7, 116–132. [Google Scholar] [CrossRef]
- Wang, S.Y.; Wang, J.; Wang, Y.; Yan, J.; Li, J. Environmental knowledge and consumers’ intentions to visit green hotels: The mediating role of consumption values. J. Travel Tour. Mark. 2018, 35, 1261–1271. [Google Scholar] [CrossRef]
- Büntgen, U.; Latorre, J.; Egli, S.; Martinez-Peña, F. Socio-economic, scientific, and political benefits of mycotourism. Ecosphere 2017, 8, e01870. [Google Scholar] [CrossRef]
- De Frutos, P.; Rodriguez-Prado, B.; Latorre, J.; Martinez-Peña, F. Environmental valuation and management of wild edible mushroom picking in Spain. For. Policy Econ. 2019, 100, 177–187. [Google Scholar] [CrossRef]
- Verbos, R.I.; Altschuler, B.; Brownlee, M.T.J. Weather studies in outdoor recreation and nature-based tourism: A research synthesis and gap analysis. Leis. Sci. 2018, 40, 533–556. [Google Scholar] [CrossRef]
- Becken, S. Measuring the effect of weather on tourism: A destination- and activity-based analysis. J. Travel Res. 2012, 52, 156–167. [Google Scholar] [CrossRef]
- Lise, W.; Tol, R.S.J. Impact of climate on tourist demand. Clim. Chang. 2002, 55, 429–449. [Google Scholar] [CrossRef]
- Matzarakis, A.; Hämmerle, M.; Endler, C.; Muthers, S.; Koch, E. Assessment of tourism and recreation destinations under climate change conditions in Austria. Meteorol. Z. 2012, 21, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Brosy, C.; Zaninovic, K.; Matzarakis, A. Quantification of climate tourism potential of Croatia based on measured data and regional modeling. Int. J. Biometeorol. 2013, 58, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Martín, M. Weather, climate, and tourism: A geographical perspective. Ann. Tour. Res. 2005, 32, 571–591. [Google Scholar] [CrossRef]
- Bigano, A.; Hamilton, J.M.; Tol, R.S.J. The impact of climate on holiday destination choice. Clim. Chang. 2006, 76, 389–406. [Google Scholar] [CrossRef]
- Andrade, H.; Alcoforado, M.J.; Oliveira, S. Perception of temperature and wind by users of public outdoor spaces: Relationships with weather parameters and personal characteristics. Int. J. Biometeorol. 2011, 55, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Pantavou, K.; Lykoudis, S. Modeling thermal sensation in a Mediterranean climate—A comparison of linear and ordinal models. Int. J. Biometeorol. 2014, 58, 1355–1368. [Google Scholar] [CrossRef]
- Rutty, M.; Scott, D. Bioclimatic comfort and the thermal perceptions and preferences of beach tourists. Int. J. Biometerol. 2014, 59, 37–45. [Google Scholar] [CrossRef]
- Rutty, M.; Scott, D. Thermal range of coastal tourism resort microclimates. Tour. Geogr. 2014, 16, 346–363. [Google Scholar] [CrossRef]
- Gómez-Martín, M.; Martínez-Ibarra, E. Tourism demand and atmospheric parameters: Nonintrusive observation techniques. Clim. Res. 2012, 51, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Lindner-Cendrowska, K. Assessment of bioclimatic conditions in cities for tourism and recreational purposes (a Warsaw case study). Geogr. Pol. 2013, 86, 55–66. [Google Scholar] [CrossRef]
- Matzarakis, A.; Hämmerle, M.; Koch, E.; Rudel, E. The climate tourism potential of Alpine destinations using the example of Sonnblick, Rauris and Salzburg. Theor. Appl. Climatol. 2012, 110, 645–658. [Google Scholar] [CrossRef]
- Hansen, M.M.; Jones, R.; Tocchini, K. Shinrin-Yoku (Forest Bathing) and Nature Therapy: A State-of-the-Art Review. Int. J. Environ. Res. Public Health 2017, 14, 851. [Google Scholar] [CrossRef] [Green Version]
- Tsunetsugu, Y.; Park, B.J.; Miyazaki, Y. Trends in research related to ‘‘Shinrin-yoku’’ (taking in the forest atmosphere or forest bathing) in Japan. Environ. Health Prev. Med. 2010, 15, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Yan, Q.; Pan, Y.; Gu, X.; Liu, Y. Medical empirical research on forest bathing (Shinrin-yoku): A systematic review. Environ. Health Prev. Med. 2019, 24, 70. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.S.; Lim, Y.-R.; Lee, K.; Lee, J.; Lee, J.H.; Lee, I.-S. Terpenes from forests and human health. Toxicol. Res. Vol. 2017, 33, 97–106. [Google Scholar] [CrossRef]
- Antonelli, M.; Donelli, D.; Barbieri, G.; Valussi, M.; Maggini, V.; Firenzuoli, F. Forest volatile organic compounds and their effects on human health: A state-of-the-art review. Int. J. Environ. Res. Public Health 2020, 17, 6506. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Mewalal, R.; Rai, D.K.; Kainer, D.; Chen, F.; Külheim, C.; Peter, G.F.; Tuskan, G.A. Plant-derived terpenes: A feedstock for specialty biofuels. Trends Biotechnol. 2017, 35, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Gershenzon, J.; Bohlmann, J. Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce. Plant Physiol. 2003, 132, 1586–1599. [Google Scholar] [CrossRef] [Green Version]
- Bach, A.; Peñuelas, J.; Clarà, J.; Llusià, J.; Campillo, I.; López, F.; Maneja, R. How should forests be characterized in regard to human health? Evidence from existing literature. Int. J. Environ. Res. Public Health 2020, 17, 1027. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Lluisá, J. The complexity of factors driving volatile organic compound emissions by plants. Biol. Plant. 2001, 44, 481–487. [Google Scholar] [CrossRef]
- Kopaczyk, J.M.; Warguła, J.; Jelonek, T. The variability of terpenes in conifers under developmental and environmental stimuli. J. Exp. Bot. 2020, 180, 104197. [Google Scholar] [CrossRef]
- Gil-Pelegrín, E.; Saz, M.A.; Cuadrat, J.M.; Peguero-Pina, J.J.; Sancho-Knapik, D. Oaks under mediterranean-type climates: Functional response to summer aridity. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Gil-Pelegrín, E., Saz, M.A., Cuadrat, J.M., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 137–193. [Google Scholar]
- Kännaste, A.; Copolovici, L.; Niinemets, Ü. Gas chromatography-mass spectrometry method for determination of biogenic volatile organic compounds emitted by plants. Methods Mol. Biol. 2014, 1153, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Mucciarelli, M.; Camusso, W.; Maffei, M.; Panicco, P.; Bicchi, C. Volatile terpenoids of endophyte-free and infected peppermint (Mentha piperita L.): Chemical partitioning of a symbiosis. Microb. Ecol. 2007, 54, 685–696. [Google Scholar] [CrossRef]
- Li, C.C. Path Analysis—A Primer; The Boxwood Press: Pacific Grove, CA, USA, 1975. [Google Scholar]
- Kimm, H.; Guan, K.; Gentine, P.; Wu, J.; Bernacchi, C.J.; Sulman, B.N.; Griffis, T.J.; Link, C. Redefining droughts for the U.S. Corn Belt: The dominant role of atmospheric vapor pressure deficit over soil moisture in regulating stomatal behavior of maize and soybean. Agric. Forest Meteorol. 2020, 287, 107930. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Pokorska, O.; Dewulf, J.; Amelynck, C.; Schoon, N.; Šimpraga, M.; Steppe, K.; Van Langenhove, H. Isoprene and terpenoid emissions from Abies alba: Identification and emission rates under ambient conditions. Atmos. Environ. 2012, 59, 501–508. [Google Scholar] [CrossRef]
- Simon, V.; Clement, B.; Riba, M.L.; Torres, L. The Landes experiment: Monoterpenes emitted from the maritime pine. J. Geophys. Res. Atmos. 1994, 99, 16501–16510. [Google Scholar] [CrossRef]
- Blanch, J.S.; Peñuelas, J.; Sardans, J.; Llusià, J. Drought, warming and soil fertilization effects on leaf volatile terpene concentrations in Pinus halepensis and Quercus ilex. Acta Physiol. Plant. 2009, 31, 207–218. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, M.H.; Park, O.H.; Park, K.S.; An, S.S.; Seo, H.J.; Jin, S.H.; Jeong, W.S.; Kang, Y.J.; An, K.W.; et al. A study on the distribution characteristics of terpene at the main trails of Mt. Mudeung. J. Environ. Health Sci. 2013, 39, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Zas, R.; Quiroga, R.; Touza, R.; Vázquez-González, C.; Sampedro, L.; Lema, M. Resin tapping potential of atlantic maritime pine forests depends on tree age and timing of tapping. Ind. Crops Prod. 2020, 157, 112940. [Google Scholar] [CrossRef]
- Bach, A.; Yáñez-Serrano, A.M.; Llusià, J.; Filella, I.; Maneja, R.; Penuelas, J. Human breathable air in a mediterranean forest: Characterization of monoterpene concentrations under the canopy. Int. J. Environ. Res. Public Health 2020, 17, 4391. [Google Scholar] [CrossRef]
- Staudt, M.; Bertin, N.; Hansen, U.; Seufert, G.; Cicciolij, P.; Foster, P.; Frenzel, B.; Fugit, J.L. Seasonal and diurnal patterns of monoterpene emissions from Pinus pinea (L.) under field conditions. Atmos. Environ. 1997, 31, 145–156. [Google Scholar] [CrossRef]
- Staudt, M.; Bertin, N.; Frenzel, B.; Seufert, G. Seasonal variation in amount and composition of monoterpenes emitted by young Pinus pinea trees–implications for emission modeling. J. Atmos. Chem. 2000, 35, 77–99. [Google Scholar] [CrossRef]
- Loreto, F.; Forster, A.; Durr, M.; Csiky, O.; Seufert, G. On the monoterpene emission under heat stress and on the increased thermotolerance of leaves of Quercus ilex L. fumigated with selected monoterpenes. Plant Cell Environ. 1998, 21, 101–107. [Google Scholar] [CrossRef]
- Loreto, F.; Pinelli, P.; Manes, F.; Kollist, H. Impact of ozone on monoterpene emissions and evidence for an isoprene-like antioxidant action of monoterpenes emitted by Quercus ilex leaves. Tree Physiol. 2004, 24, 361–367. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Simms, E.L.; Rausher, M.D. Costs and benefits of plant resistance to herbivory. Am. Nat. 1987, 130, 570–581. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- Lerdau, M.; Matson, P.; Fall, R.; Monson, R. Ecological controls over monoterpene emissions from douglas-fir (Pseudotsuga menziesii). Ecology 1995, 76, 2640–2647. [Google Scholar] [CrossRef]
- Burney, O.T.; Davis, A.S.; Jacobs, D.F. Phenology of foliar and volatile terpenoid production for Thuja plicata families under differential nutrient availability. Environ. Exp. Bot. 2012, 77, 44–52. [Google Scholar] [CrossRef]
- Breckle, S.W. Walter’s Vegetation of the Earth. The Ecological Systems of the Geo-Biosphere, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2002; p. 527. [Google Scholar]
- Turtola, S.; Manninen, A.M.; Rikala, R.; Kainulainen, P. Drought stress alters the concentration of wood terpenoids in Scots pine and Norway spruce seedlings. J. Chem. Ecol. 2003, 29, 1981–1995. [Google Scholar] [CrossRef]
- Sancho-Knapik, D.; Sanz, M.Á.; Peguero-Pina, J.J.; Niinemets, Ü.; Gil-Pelegrín, E. Changes of secondary metabolites in Pinus sylvestris L. Needles under increasing soil water deficit. Ann. For. Sci. 2017, 74, 24. [Google Scholar] [CrossRef]
- Llusià, J.; Peñuelas, J. Changes in terpene content and emission in potted Mediterranean woody plants under severe drought. Can. J. Bot. 1998, 76, 1366–1373. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Reichstein, M.; Staudt, M.; Seufert, G.; Tenhunen, J.D. Stomatal constraints may affect emission of oxygenated monoterpenoids from the foliage of Pinus pinea. Plant Physiol. 2002, 130, 1371–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copolovici, L.; Niinemets, U. Environmental impacts on plant volatile emission. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 175–210. [Google Scholar]
- Hodges, J.D.; Lorio, P.L. Moisture stress and composition of xylem oleoresin in loblolly pine. For. Sci. 1975, 21, 283–290. [Google Scholar] [CrossRef]
- Wainhouse, D. Hylobius abietis—Host utilisation and resistance. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Gregoire, J.C., Evans, H.F., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 365–379. [Google Scholar]
- Amelung, B.; Viner, D. Mediterranean tourism: Exploring the future with the Tourism Climatic Index. J. Sustain. Tour. 2006, 14, 349–366. [Google Scholar] [CrossRef]
- Bafaluy, D.; Amengual, A.; Romero, R.; Homar, V. Present and future climate resources for various types of tourism in the Bay of Palma, Spain. Reg. Environ. Chang. 2014, 14, 1995–2006. [Google Scholar] [CrossRef]
- Paudyal, R.; Stein, T.V.; Kc, B.; Adams, D.C. Effects of weather factors on recreation participation in a humid subtropical region. Int. J. Biometeorol. 2019, 63, 1025–1038. [Google Scholar] [CrossRef]
- Wilkins, E.J.; Chikamoto, Y.; Miller, A.B.; Smith, J.W. Climate change and the demand for recreational ecosystem services on public lands in the continental United States. Glob. Environ. Chang. 2021, 70, 102365. [Google Scholar] [CrossRef]
- Buckley, L.B.; Foushee, M.S. Footprints of climate change in US national park visitation. Int. J. Biometeorol. 2012, 56, 1173–1177. [Google Scholar] [CrossRef]
- Schiller, G. Biometeorology and recreation in east Mediterranean forests. Landsc. Urban Plan. 2001, 57, 1–12. [Google Scholar] [CrossRef]
- Takayama, N.; Kagawa, T.; Kasetani, T.; Park, B.; Tsunetsugu, Y.; Oishi, Y.; Hirano, H.; Miyazaki, Y. The comfortableness of the light/thermal environment for bathing in the forest atmosphere. J. Jpn. Inst. Landsc. Arch. 2005, 68, 819–824. [Google Scholar] [CrossRef] [Green Version]
- Richardson, M.; McEwan, K.; Maratos, F.; Sheffield, D. Joy and calm: How an evolutionary functional model of affect regulation informs positive emotions in nature. Evol. Psychol. Sci. 2016, 2, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Farrow, M.R.; Washburn, K. A review of field experiments on the effect of forest bathing on anxiety and heart rate variability. Glob. Adv. Health Med. 2019, 8, 1–7. [Google Scholar] [CrossRef]
- Kotera, Y.; Richardson, M.; Sheffield, D. Effects of Shinrin-yoku (forest bathing) and nature therapy on mental health: A systematic review and meta-analysis. Int. J. Ment. Health Addict. 2022, 20, 337–361. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
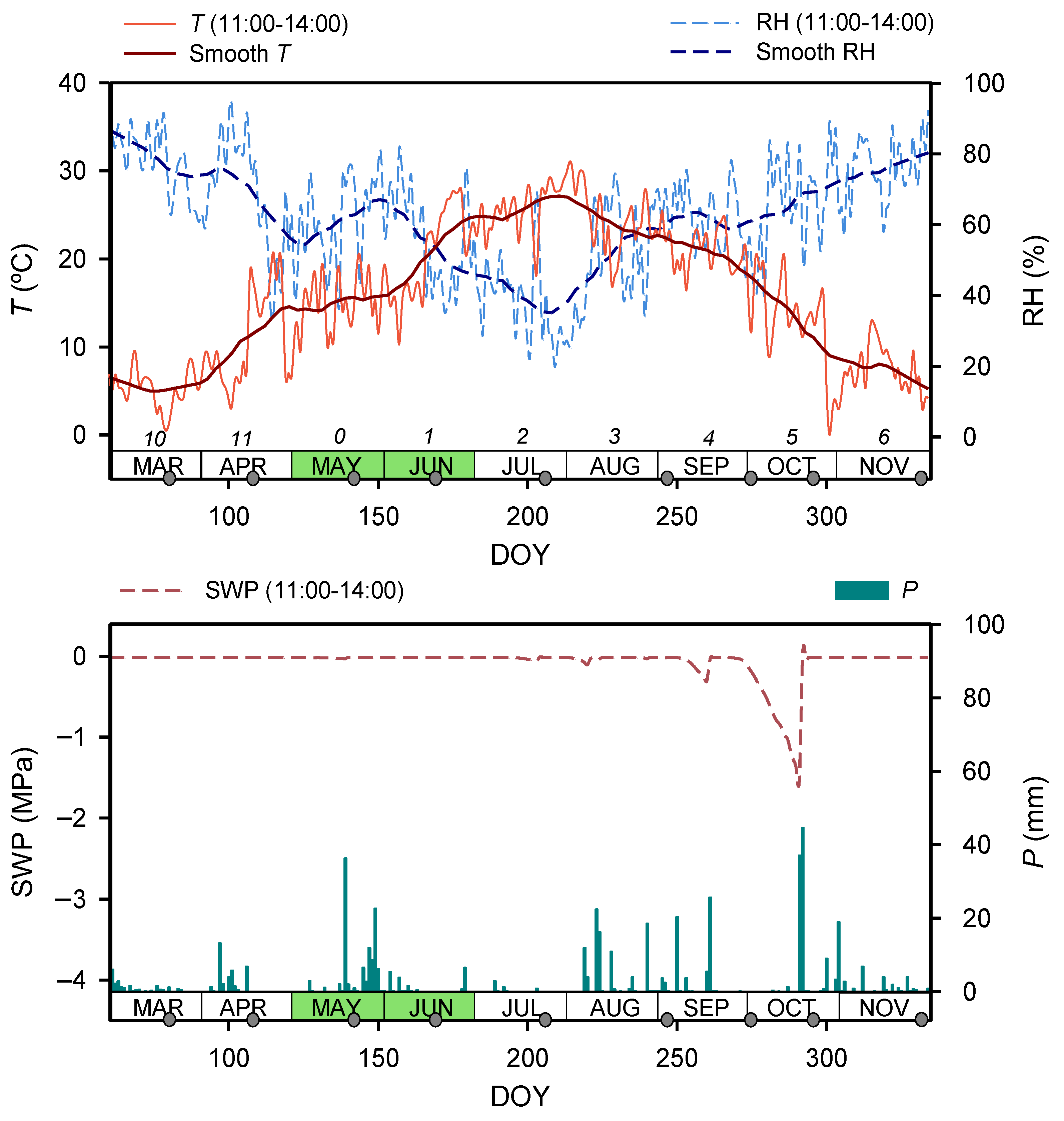
| DOY | Month | T15 (°C) Mean ± se (min; max) | SWP15 (Mpa) | Tc (°C) Mean ± se 1 | SWPc (Mpa) 2 | ||
|---|---|---|---|---|---|---|---|
| 11:00 a.m. h | 12:00 a.m. h | 13:00 p.m. h | |||||
| 79 | March | 9.8 ± 0.7 (5.3; 14.4) | −0.013 | 1.8 ± 0.3 | 0.6 ± 0.1 | 1.1 ± 0.1 | −0.013 |
| 107 | April | 12.2 ± 1.2 (2.8; 19.3) | −0.013 | 18 ± 0.7 | 18.8 ± 0.4 | 19.6 ± 0.3 | −0.013 |
| 142 | May | 18.8 ± 0.7 (13.8; 23.7) | −0.031 | 20.3 ± 0.5 | 18.5 ± 0.2 | 14.9 ± 0.2 | −0.012 |
| 170 | June | 20.4 ± 1.0 (13.2; 27.0) | −0.012 | 26.2 ± 0.5 | 27.3 ± 0.3 | 27 ± 0.3 | −0.012 |
| 205 | July | 28.7 ± 0.7 (20.1; 31.3) | −0.063 | 27.8 ± 0.3 | 28.9 ± 0.3 | 31.2 ± 0.5 | −0.012 |
| 247 | September | 28.6 ± 0.6 (24.9; 32.7) | −0.034 | 23.2 ± 0.3 | 26.8 ± 0.2 | 28.9 ± 0.8 | −0.015 |
| 275 | October | 24.6 ± 0.7 (19.4; 29.6) | −0.288 | 18.3 ± 0.3 | 21.5 ± 0.8 | 22.9 ± 0.2 | −0.198 |
| 296 | October | 16.6 ± 0.8 (11.3; 22.6) | −1.495 | 12.1 ± 0.3 | 15.3 ± 0.6 | 15.3 ± 0.4 | −0.011 |
| 331 | November | 9.9 ± 0.7 (6.7; 15.1) | −0.011 | 10.1 ± 1.0 | 11 ± 0.6 | 11.5 ± 0.4 | −0.011 |
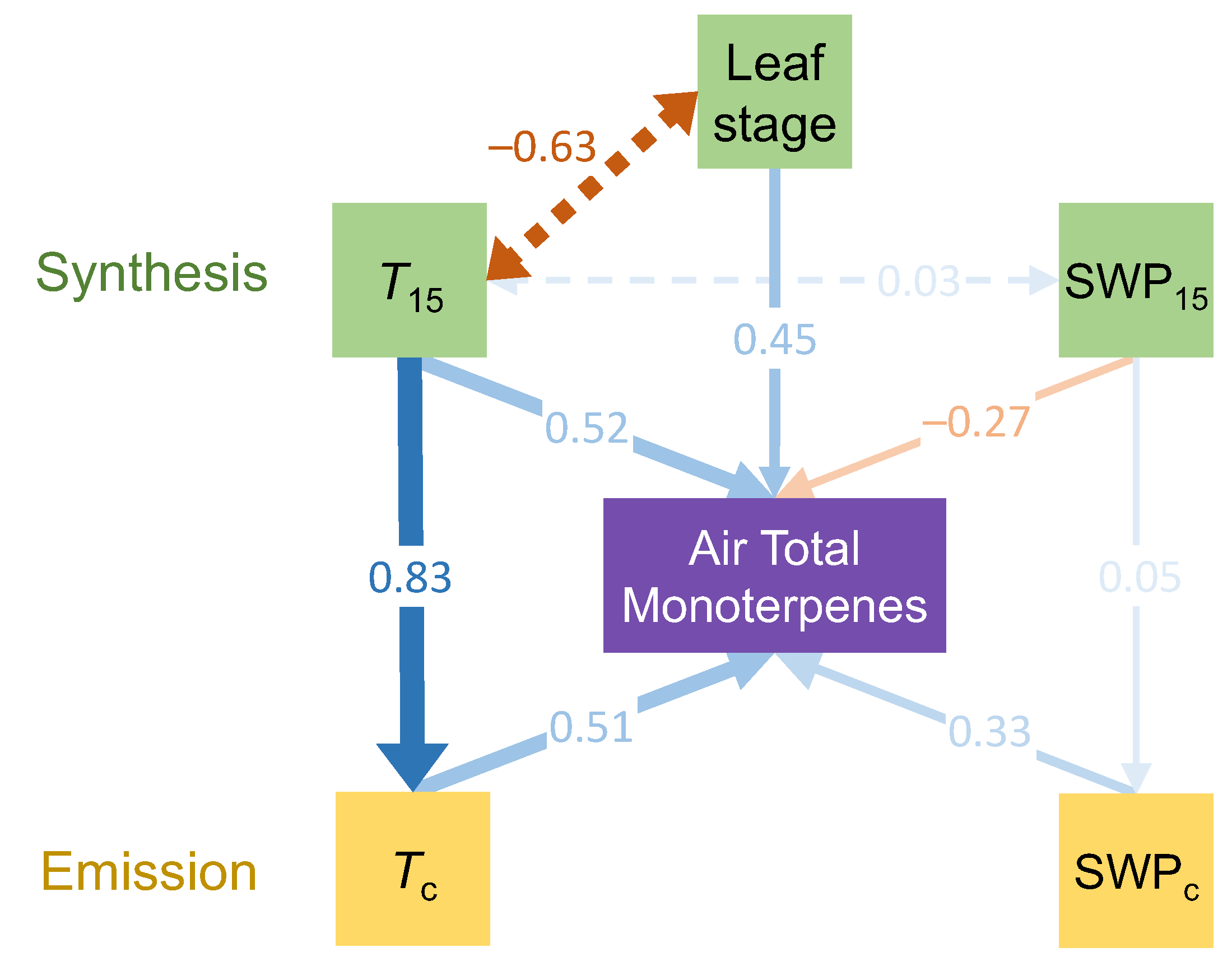
| R2 | UC | SE | SC | P | |
|---|---|---|---|---|---|
| Monoterpenes~Tc + SWPc + Leaf stage + SWP15 + T15 | 0.795 | <0.001 | |||
| Tc | 0.727 | 0.221 | 0.509 | 0.001 | |
| SWPc | 67.507 | 18.091 | 0.325 | <0.001 | |
| Leaf stage | 1.55 | 0.382 | 0.453 | <0.001 | |
| SWP15 | −7.194 | 2.303 | −0.273 | 0.002 | |
| T15 | 0.904 | 0.295 | 0.521 | 0.002 | |
| Tc~T15 | 0.683 | <0.001 | |||
| T15 | 1.004 | 0.132 | 0.826 | <0.001 | |
| SWPc~SWP15 | 0.002 | 0.812 | |||
| SWP15 | 0.006 | 0.024 | 0.046 | 0.812 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho-Knapik, D.; Gil-Pelegrín, E.; Ferrio, J.P.; Alonso-Forn, D.; Martín-Sánchez, R.; Santos Silva, J.V.d.; Imanishi, J.; Peguero-Pina, J.J.; Sanz, M.Á. Changes in the Abundance of Monoterpenes from Breathable Air of a Mediterranean Conifer Forest: When Is the Best Time for a Human Healthy Leisure Activity? Forests 2022, 13, 965. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060965
Sancho-Knapik D, Gil-Pelegrín E, Ferrio JP, Alonso-Forn D, Martín-Sánchez R, Santos Silva JVd, Imanishi J, Peguero-Pina JJ, Sanz MÁ. Changes in the Abundance of Monoterpenes from Breathable Air of a Mediterranean Conifer Forest: When Is the Best Time for a Human Healthy Leisure Activity? Forests. 2022; 13(6):965. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060965
Chicago/Turabian StyleSancho-Knapik, Domingo, Eustaquio Gil-Pelegrín, Juan Pedro Ferrio, David Alonso-Forn, Rubén Martín-Sánchez, José Víctor dos Santos Silva, Junichi Imanishi, José Javier Peguero-Pina, and María Ángeles Sanz. 2022. "Changes in the Abundance of Monoterpenes from Breathable Air of a Mediterranean Conifer Forest: When Is the Best Time for a Human Healthy Leisure Activity?" Forests 13, no. 6: 965. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060965