Invasion of Winter Moth in New England: Effects of Defoliation and Site Quality on Tree Mortality

Abstract
:1. Introduction
Winter Moth in Massachusetts
2. Methods
2.1. Study Region
2.2. Study Sites
2.3. Vegetation Sampling
2.4. Determination of Winter Moth Presence
2.5. Dendrochronology
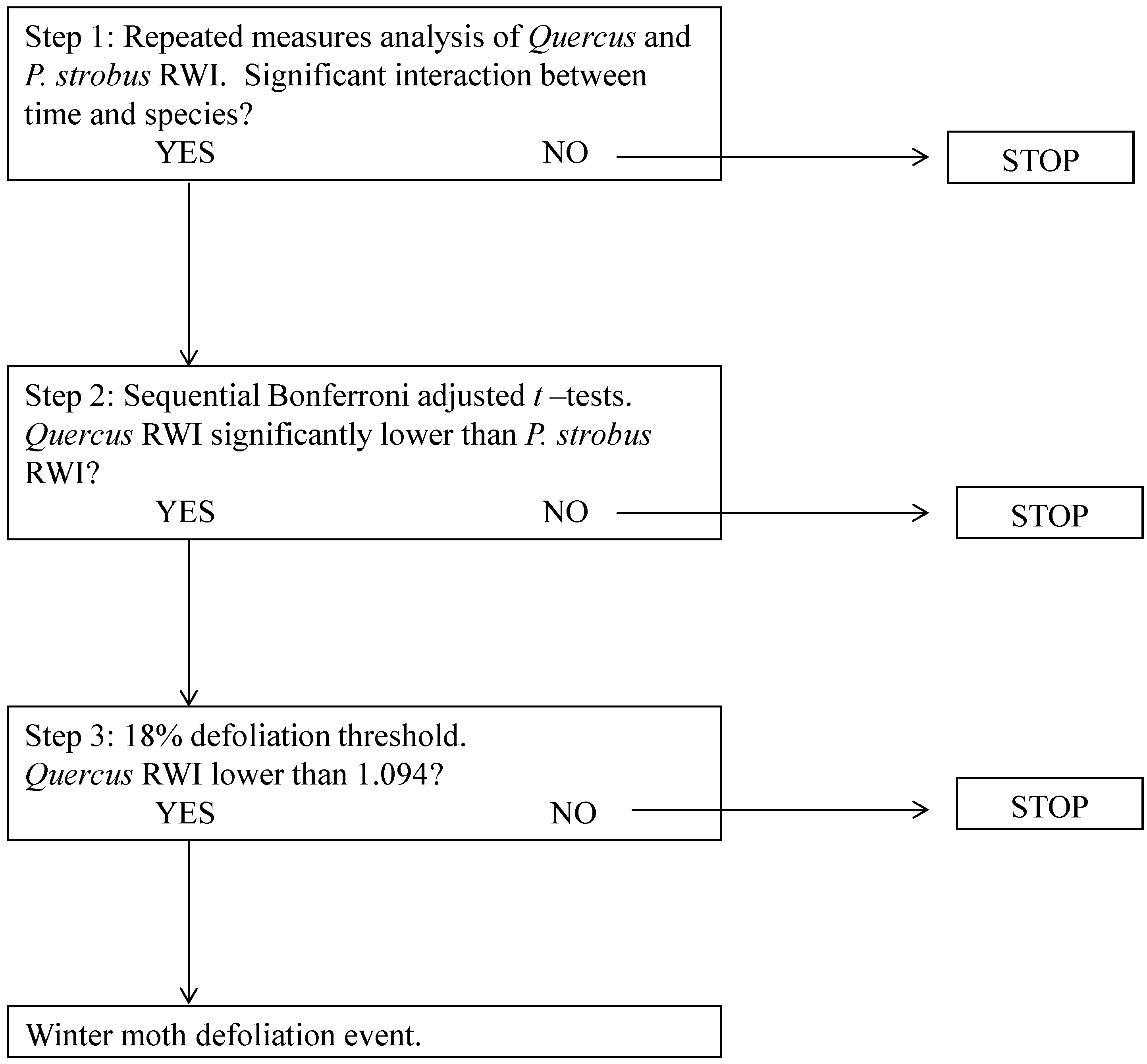
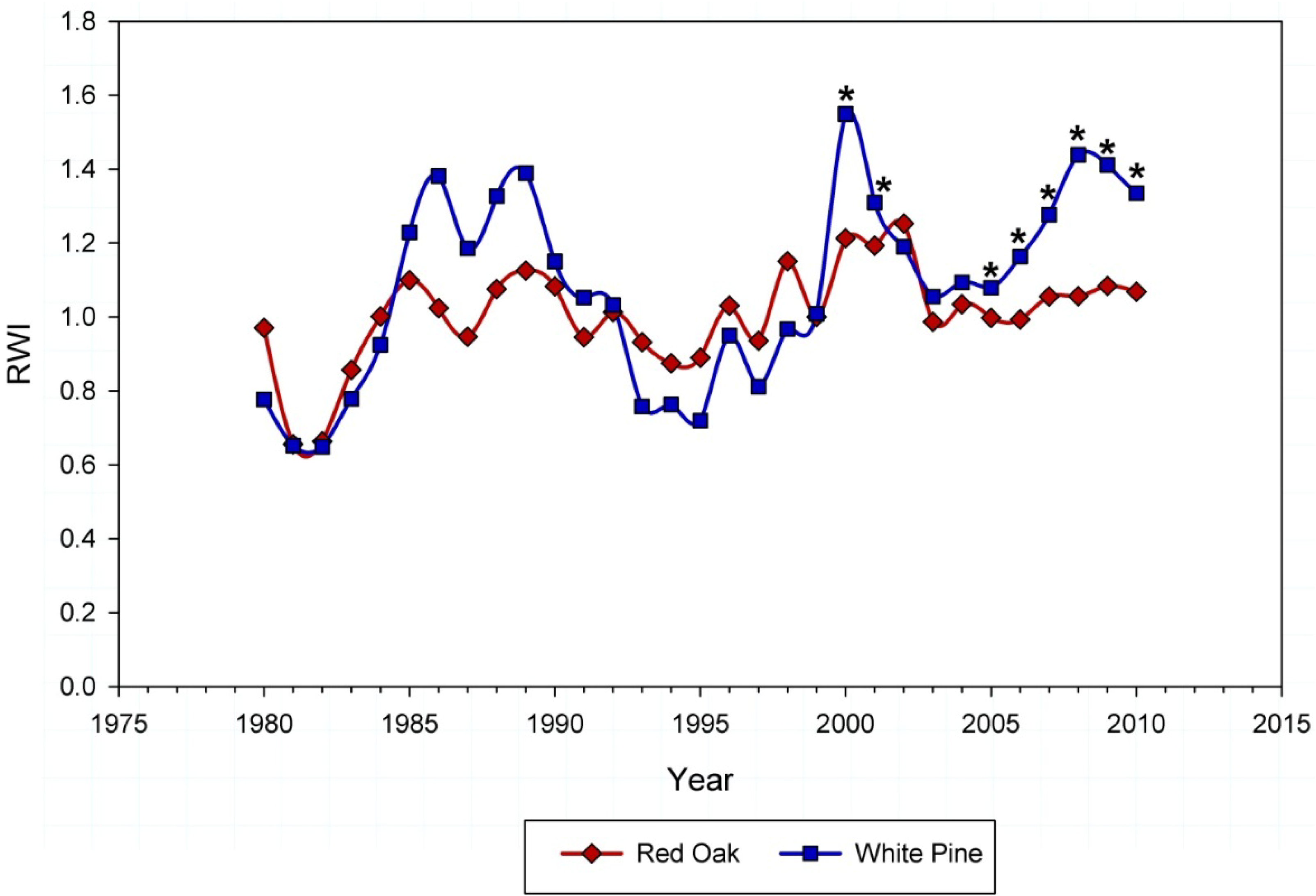
2.6. Defining Winter Moth Activity

2.7. Percentage of Quercus Basal Area Consisting of Dead Stems
2.8. Understory Density
2.9. Drought
3. Results
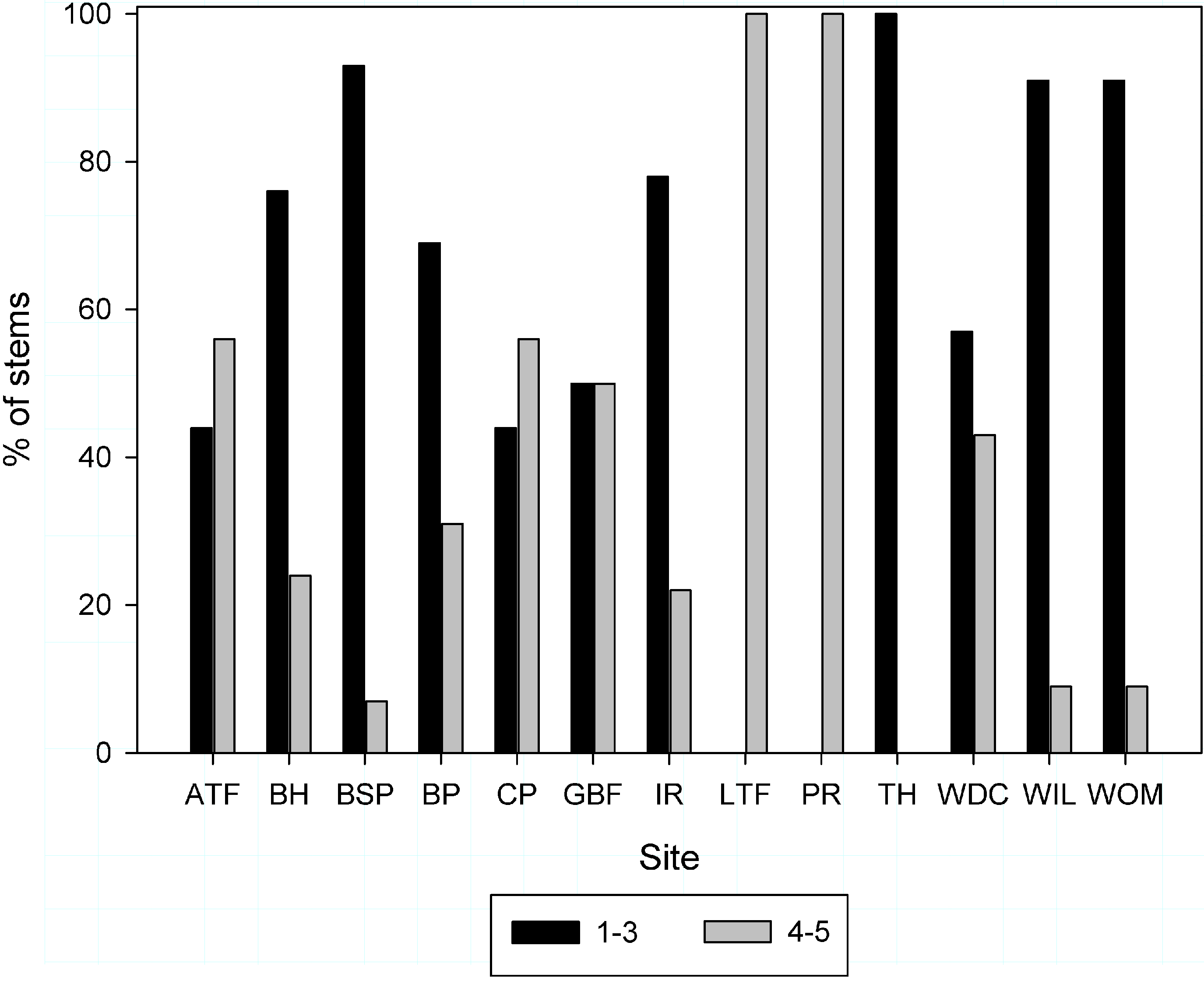
3.1. Study Sites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location | Quercus Density (stems/ha) | Quercus Mean DBH (cm) | Understory Plant Density |
|---|---|---|---|---|
| Amesbury TF | 42°51′ N; −70°58′ W | 315.2 | 32.3 ± 1.3 | 35,230 |
| Blue Hills SR | 42°13′ N; −71°07′ W | 245.3 | 25.9 ± 1.1 | 261,313 |
| Borderland SP | 42°04′ N; −71°10′ W | 344.4 | 23.5 ± 0.8 | 136,943 |
| Bradley Palmer SP | 42°39’N; −70°54′ W | 114.9 | 33.7 ± 2.8 | 95,226 |
| Centennial Park | 42°18′ N; −71°16′ W | 237.4 | 35.9 ± 1.3 | 42,794 |
| Great Brook Farm SP | 42°33′ N; −71°21′ W | 177.0 | 37.0 ± 1.5 | 51,524 |
| Iron Rail | 42°36′ N; −70°51′ W | 271.0 | 32.7 ± 2.2 | 48,168 |
| Lockwood TF | 42°38′ N; −70°59′ W | 141.4 | 36.5 ± 1.8 | 60,830 |
| Profile Rock SP | 41°48′ N; −71°03′ W | 140.0 | 39.5 ± 2.9 | 112,795 |
| Turkey Hill CA | 42°41′ N; −70°52′ W | 78.7 | 39.6 ± 1.8 | 59,600 |
| Wildcat CA | 42°41′ N; −71°01′ W | 309.4 | 27.8 ± 0.7 | 11,306 |
| Willowdale SF | 42°40′ N; −70°54′ W | 201.5 | 37.2 ± 2.1 | 155,749 |
| Wompatuck SP | 42°13′ N; −70°52′ W | 248.4 | 29.3 ± 0.8 | 92,554 |
| Site | Quercus SI | Stand RD | Quercus Age | P. strobus Age | % Quercus in Dead Stems | Years and # of WM Def. Events |
|---|---|---|---|---|---|---|
| Amesbury TF | 77 | 0.83 | 76 | 86 | 6% | 1999; 2001 (2) |
| Blue Hills SR | 45 | 0.76 | 85 | 84 | 18% | 2008–2010 (3) |
| Borderland SP | 50 | 0.69 | 106 | 68 | 12% | 2006; 2008-2010 (4) |
| Bradley Palmer SP | 66 | 0.84 | 89 | 107 | 28% | 2007–2010 (4) |
| Centennial Park | 65 | 0.86 | 89 | 65 | 4% | 0 |
| Great Brook Farm SP | 71 | 0.74 | 95 | 81 | 2% | 1996; 2005 (2) |
| Iron Rail TP | 72 | 0.71 | 59 | 53 | 5% | 0 |
| Lockwood TF | 62 | 0.60 | 89 | 78 | 0% | 2001; 2010 (2) |
| Profile Rock SP | 60 | 0.81 | 94 | 64 | 3% | 2005 (1) |
| Turkey Hill CA | 65 | 0.71 | 67 | 84 | 4% | 0 |
| Wildcat CA | 66 | 0.60 | 72 | 85 | 7% | 0 |
| Willowdale SF | 68 | 0.88 | 90 | 89 | 4% | 2008-2010 (3) |
| Wompatuck SP | 46 | 0.61 | 97 | 89 | 30% | 2000; 2008 (2) |

3.2. Dendrochronology

| Tree | DBH (cm) | Sang Class | Year of Mortality | Years Since Mortality |
|---|---|---|---|---|
| 1 | 30.5 | 2 | 2008 | 2 |
| 2 | 27.3 | 2 | 2006 | 4 |
| 3 | 35.8 | 2 | 1999 | 11 |
| 4 | 25.6 | 3 | 1997 | 13 |
| 5 | 32.5 | 3 | 1996 | 14 |
| 6 | 24.0 | 4 | 1987 | 23 |
3.3. Drought
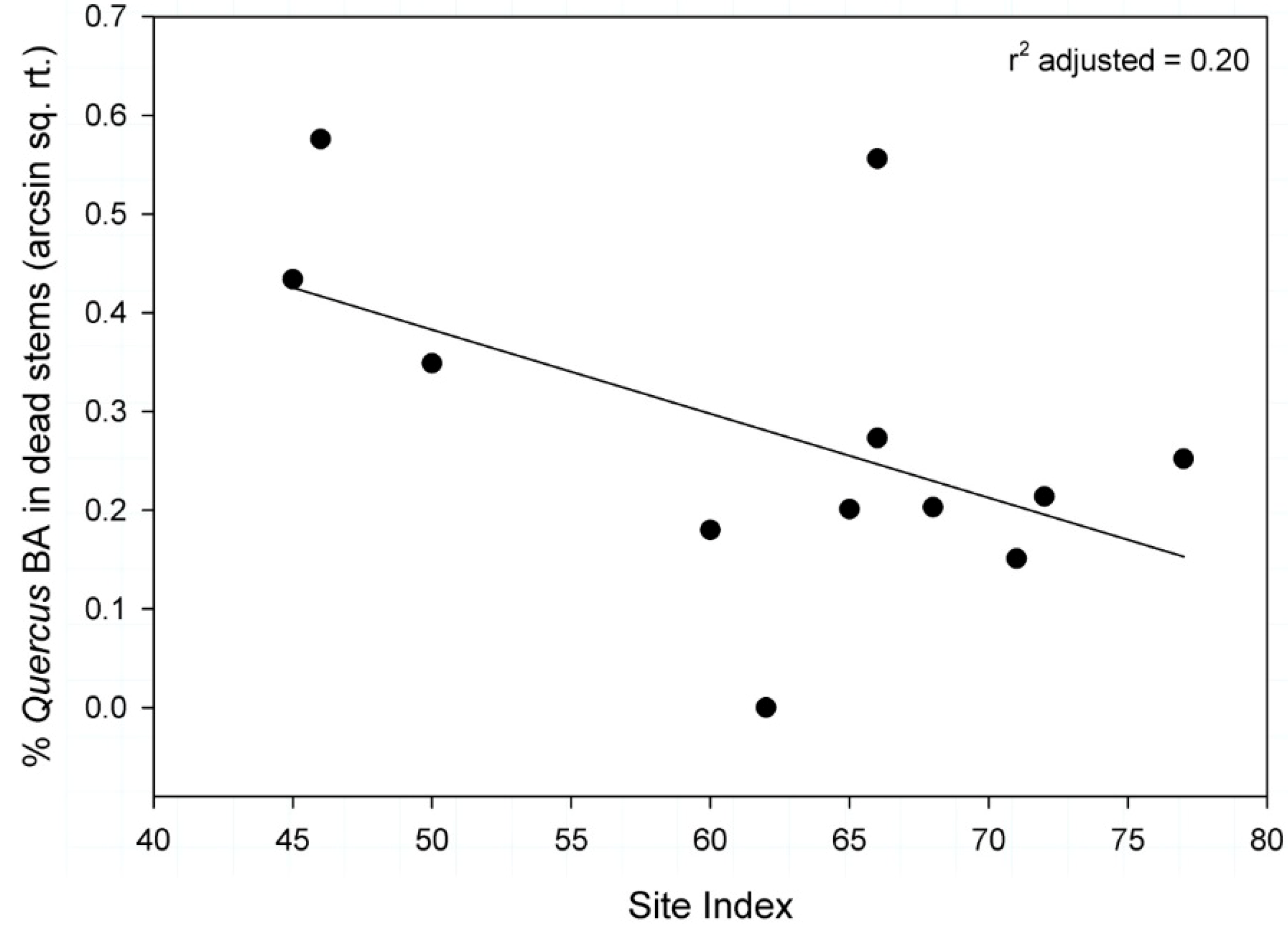
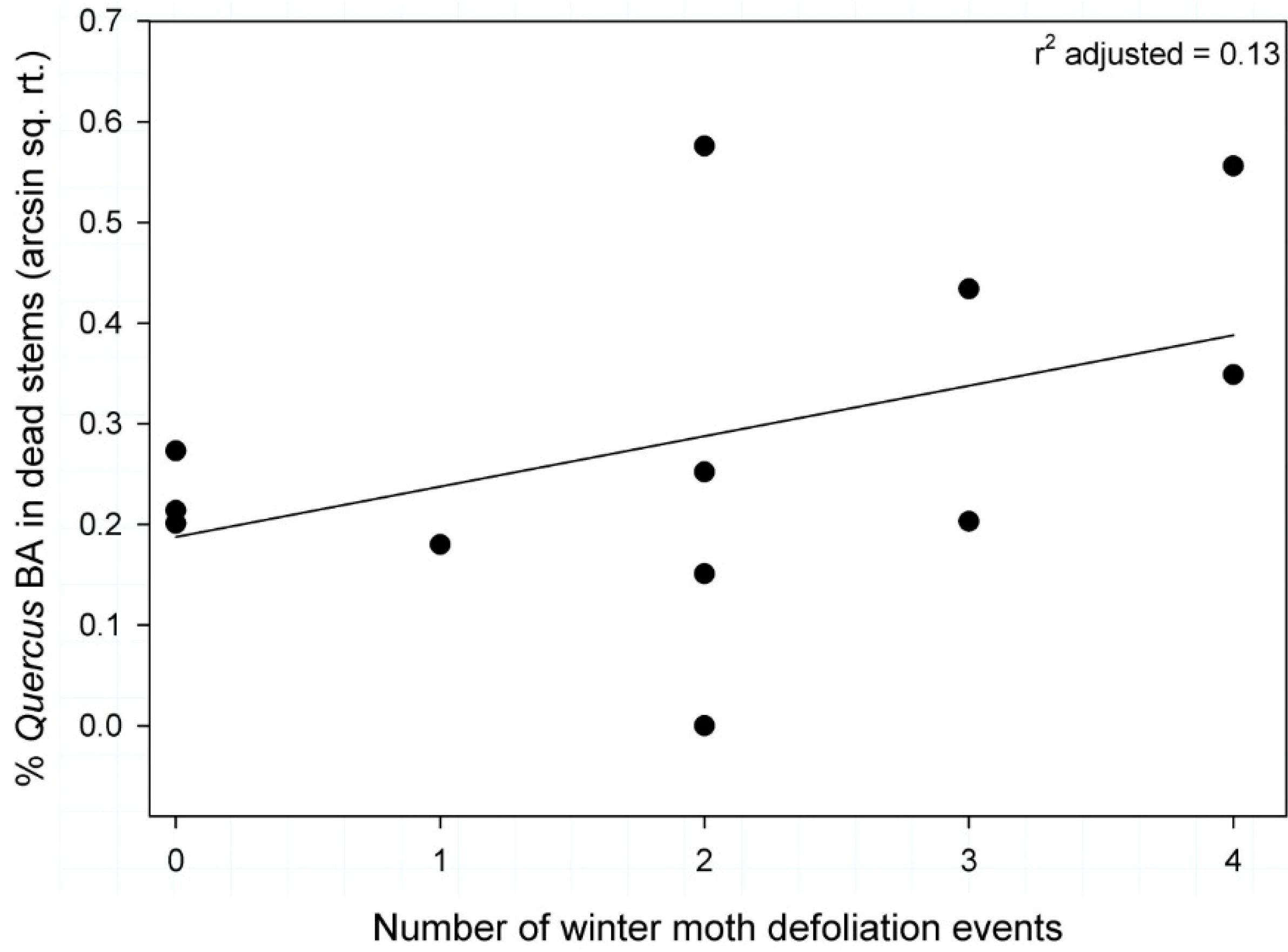
3.4. Percentage of Quercus Basal Area Consisting of Dead Stems

3.5. Understory Density

| Model Parameters | ΔAICc | r2adj | Parameter Estimate | SE | Log Likelihood | Akaike Weight | Evidence Ratio |
|---|---|---|---|---|---|---|---|
| Stand RD | 4.03 | −0.09 | 0.030 | 0.497 | 0.13 | 0.06 | 7.51 |
| Site Index | 3.48 | 0.18 | −0.010 | 0.005 | 0.18 | 0.08 | 5.69 |
| Stand RD | 0.381 | 0.463 | |||||
| Site Index | 2.78 | 0.22 | −0.007 | 0.005 | 0.25 | 0.12 | 4.02 |
| # WM defoliation events | 0.034 | 0.030 | |||||
| # WM defoliation events | 1.04 | 0.13 | 0.050 | 0.030 | 0.59 | 0.28 | 1.68 |
| Site Index | 0.00 | 0.20 | −0.009 | 0.004 | 1.00 | 0.46 | 1.00 |
| Model Parameters | ΔAICc | r2adj | Parameter Estimate | SE | Log Likelihood | Akaike Weight | Evidence Ratio |
|---|---|---|---|---|---|---|---|
| Stand RD | 9.42 | 0.19 | 77,065.34 | 178,423.80 | 0.01 | 0.01 | 110.89 |
| # WM defoliation events | 26,524.87 | 11,992.21 | |||||
| Stand RD * # WM defoliation events | −50,165.78 | 135,010.40 | |||||
| Stand RD | 5.17 | −0.03 | 159,237.48 | 196,194.80 | 0.08 | 0.06 | 13.27 |
| Stand RD | 4.04 | 0.26 | 80,855.00 | 170,282.20 | 0.13 | 0.11 | 7.55 |
| # WM defoliation events | 26,031.14 | 11,393.15 | |||||
| # WM defoliation events | 0.00 | 0.31 | 27,121.03 | 10,759.48 | 1.00 | 0.82 | 1.00 |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liebhold, A.M.; McCullough, D.G.; Blackburn, L.M.; Frankel, S.J.; Von Holle, B.; Aukema, J.E. A highly aggregated geographical distribution of forest pest invasions in the USA. Divers. Distrib. 2013, 19, 1208–1216. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol. Invasions 2010, 12, 389–405. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS One 2011, 6, 7. [Google Scholar] [CrossRef]
- Baker, W.L. Effect of gypsy moth defoliation on certain forest trees. J. For. 1941, 39, 1017–1022. [Google Scholar]
- Orwig, D.A.; Foster, D.R. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Dodds, K.J.; Orwig, D.A. An invasive urban forest pest invades natural environments—Asian longhorned beetle in northeastern US hardwood forests. Can. J. For. Res. 2011, 41, 1729–1742. [Google Scholar] [CrossRef]
- Cuming, F.G. The distribution, life history, and economic importance of the winter moth, Operophtera brumata (L.) (Lepidoptera: Geometridae) in Nova Scotia. Can. Entomol. 1961, 93, 135–142. [Google Scholar] [CrossRef]
- Feeny, P. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars. Ecology 1970, 51, 565–581. [Google Scholar] [CrossRef]
- Varley, G.C.; Gradwell, G.R.; Hassell, M.P. Insect Population Ecology: An Analytical Approach; University of California Press: Berkeley, CA, USA, 1974. [Google Scholar]
- Elkinton, J.S.; Boettner, G.H.; Sermac, M.; Gwiazdowski, R.; Hunkins, R.R.; Callahan, J.; Scheufele, S.B.; Donahue, C.P.; Porter, A.H.; Khrimian, A.; et al. Survey for winter moth (Lepidoptera: Geometridae) in northeastern North America with pheromone-baited traps and hybridization with the native Bruce spanworm (Lepidoptera: Geometridae). Ann. Entomol. Soc. Am. 2010, 103, 135–145. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Liebhold, A.; Boettner, G.H.; Sremac, M. Invasion spread of Operophtera brumata in northeastern United States and hybridization with O. bruceata. Biol. Invasions 2014, 16, 2263–2272. [Google Scholar] [CrossRef]
- Wesolowski, T.; Rowinski, P. Tree defoliation by winter moth Operophtera brumata L. during an outbreak affected by structure of forest landscape. For. Ecol. Manag. 2006, 221, 299–305. [Google Scholar] [CrossRef]
- Gillespie, D.R.; Finlayson, T.; Tonks, N.V.; Ross, D.A. Occurrence of winter moth, Operophtera brumata (Lepidoptera: Geometridae), on southern Vancouver Island, British Columbia. Can. Entomol. 1978, 110, 223–224. [Google Scholar] [CrossRef]
- Roland, J. Decline in winter moth populations in North America—Direct versus indirect effect of introduced parasites. J. Anim. Ecol. 1988, 57, 523–531. [Google Scholar] [CrossRef]
- Tikkanen, O.P.; Lyytikainen-Saarenmaa, P. Adaptation of a generalist moth, Operophtera brumata, to variable budburst phenology of host plants. Entomol. Exp. Appl. 2002, 103, 123–133. [Google Scholar] [CrossRef]
- Tikkanen, O.P.; Julkunen-Tiitto, R. Phenological variation as protection against defoliating insects: The case of Quercus robur and Operophtera brumata. Oecologia 2003, 136, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Embree, D.G. The winter moth Operophtera brumata in eastern Canada, 1962–1988. For. Ecol. Manag. 1991, 39, 47–54. [Google Scholar] [CrossRef]
- Varley, G.C.; Gradwell, G.R. The effect of partial defoliation by caterpillars on the timber production of oak trees in England. In Proceedings of the 11th International Congress of Entomology, Vienna, Austria, 17–25 August, 1960; pp. 211–214.
- Embree, D.G. Effects of winter moth on growth and mortality of red oak in Nova Scotia. For. Sci. 1967, 13, 295–299. [Google Scholar]
- Tikkanen, O.P.; Roininen, H. Spatial pattern of outbreaks of Operophtera brumata in eastern Fennoscandia and their effects on radial growth of trees. For. Ecol. Manag. 2001, 146, 45–54. [Google Scholar] [CrossRef]
- Simmons, M.J.; Lee, T.D.; Ducey, M.J.; Elkinton, J.S.; Boettner, G.H.; Dodds, K.J. Effects of invasive winter moth defoliation on tree radial growth in eastern Massachusetts, USA. Insects 2014, 5, 301–318. [Google Scholar] [CrossRef]
- Monserud, R.A. Simulation of forest tree mortality. For. Sci. 1976, 22, 438–444. [Google Scholar]
- Wyckoff, P.H.; Clark, J.S. Predicting tree mortality from diameter growth: A comparison of maximum likelihood and Bayesian approaches. Can. J. For. Res. 2000, 30, 156–167. [Google Scholar] [CrossRef]
- Gottschalk, K.W.; Colbert, J.J.; Feicht, D.L. Tree mortality risk of oak due to gypsy moth. Eur. J. For. Pathol. 1998, 28, 121–132. [Google Scholar] [CrossRef]
- Campbell, R.W.; Sloan, R.J. Forest stand responses to defoliation by gypsy moth. For. Sci. 1977, 23, 1–34. [Google Scholar]
- Reinikainen, M.; D’Amato, A.W.; Fraver, S. Repeated insect outbreaks promote multi-cohort aspen mixedwood forests in northern Minnesota, USA. For. Ecol. Manag. 2012, 266, 148–159. [Google Scholar] [CrossRef]
- Fajvan, M.A.; Wood, J.M. Stand structure and development after gypsy moth defoliation in the Appalachian Plateau. For. Ecol. Manag. 1996, 89, 79–88. [Google Scholar] [CrossRef]
- Jedlicka, J.; Vandermeer, J.; Aviles-Vazquez, K.; Barros, O.; Perfecto, I. Gypsy moth defoliation of oak trees and a positive response of red maple and black cherry: An example of indirect interaction. Am. Midl. Nat. 2004, 152, 231–236. [Google Scholar] [CrossRef]
- Macphee, A.W. Winter moth, Operophtera brumata (Lepidoptera: Geometridae), a new pest attacking apple orchards in Nova Scotia and its coldhardiness. Can. Entomol. 1967, 99, 829–834. [Google Scholar] [CrossRef]
- Foster, D.R.; Donahue, B.M.; Kittredge, D.B.; Lambert, K.F.; Hunter, M.L.; Hall, B.R.; Irland, L.C.; Lilieholm, R.J.; Orwig, D.A.; D’Amato, A.W.; et al. Wildlands and Woodlands: A Vision for the New England Landscape; Harvard University Press: Petersham, MA, USA; Cambridge, MA, USA, 2010. [Google Scholar]
- Hall, B.; Motzkin, G.; Foster, D.R.; Syfert, M.; Burk, J. Three hundred years of forest and land-use change in Massachusetts, USA. J. Biogeogr. 2002, 29, 1319–1335. [Google Scholar] [CrossRef]
- Thomas, J.W. Wildlife Habitats in Managed Forests: The Blue Mountains of Oregon and Washington; Wildlife Management Institute: U.S. Department of Interior, Bureau of Land Management: U.S.G.P.O.: Washington, DC, USA, 1979.
- Burns, R.M.; Honkala, B.H. Silvics of North America: Vol. 2. Hardwoods; U.S. Department of Agriculture, Forest Service: U.S.G.P.O.: Washington, DC, USA, 1991.
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Tardif, J.C.; Conciatori, F. A comparison of ring-width and event-year chronologies derived from white oak (Quercus alba) and northern red oak (Quercus rubra), southwestern Quebec, Canada. Dendrochronologia 2006, 23, 133–138. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Team, R.D.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Ducey, M.J.; Knapp, R.A. A stand density index for complex mixed species forests in the northeastern United States. For. Ecol. Manag. 2010, 260, 1613–1622. [Google Scholar] [CrossRef]
- Olson, D.J. Site Index Curves for Upland Oak in the Southeast; Southeastern Forest Experiment Station, Forest Service, U.S. Department of Agriculture: Asheville, NC, USA, 1959. [Google Scholar]
- Carmean, W.H.; Hahn, J.T.; Jacobs, R.D. Site Index Curves for Forest Tree Species in the Eastern United States; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: Saint Paul, MN, USA, 1989. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- National Oceanic and Atmospheric Administration, National Climatic Data Center. Massachusetts Palmer Drought Severity Index. Available online: http://www.ncdc.noaa.gov/cag/time-series/us (accessed on 31 August 2014).
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological Drought; Research Paper No. 45. U.S.; Weather Bureau. U.S.; Government Printing Office: Washington, DC, USA, 1965; p. 58. [Google Scholar]
- Abrams, M.D. Distribution, historical development and ecophysiological attributes of oak species in the eastern United States. Ann. Sci. For. 1996, 53, 487–512. [Google Scholar] [CrossRef]
- Swain, P.C.; Kearsley, J.B. Classification of the Natural Communities of Massachusetts; Natural Heritage & Endangered Species Program, Massachusetts Division of Fisheries and Wildlife: Westborough, MA, USA, 2001. Volume 1.3. Available online: http://www.mass.gov/eea/agencies/dfg/dfw/natural-heritage/natural-communities/classification-of-natural-communities.html# (accessed on 8 October 2014).
- Michelot, A.; Breda, N.; Damesin, C.; Dufrene, E. Differing growth responses to climatic variations and soil water deficits of Fagus sylvatica, Quercus petraea and Pinus sylvestris in a temperate forest. For. Ecol. Manag. 2012, 265, 161–171. [Google Scholar] [CrossRef]
- Orwig, D.A.; Abrams, M.D. Variation in radial growth responses to drought among species, site, and canopy strata. Trees Struct. Funct. 1997, 11, 474–484. [Google Scholar] [CrossRef]
- Muzika, R.M.; Liebhold, A.M. Changes in radial increment of host and nonhost tree species with gypsy moth defoliation. Can. J. For. Res. 1999, 29, 1365–1373. [Google Scholar] [CrossRef]
- Wickman, B.E. Increased growth of white fir after a douglas-fir tussock moth outbreak. J. For. 1980, 78, 31–33. [Google Scholar]
- Lilja, R.L.; Simmons, M.J.; (USDA Forest Service, Forest Health Protection, Durham, NH, USA.). Compilation of Massachusetts Insect and Disease Detection Survey Data from 1997–2010. Extracted from the Forest Health Technology Enterprise Team database of annual Insect and Disease Detection Surveys (IDS) conducted by Massachusetts Department of Conservation and Recreation and USDA Forest Service. Unpublished work. 2014.
- Daniels, L.D.; Dobry, J.; Klinka, K.; Feller, M.C. Determining year of death of logs and snags of Thuja plicata in southwestern coastal British Columbia. Can. J. For. Res. Rev. 1997, 27, 1132–1141. [Google Scholar] [CrossRef]
- Yamasaki, M.; Leak, W.B. Snag longevity in managed northern hardwoods. North. J. Appl. For. 2006, 23, 215–217. [Google Scholar]
- Garber, S.M.; Brown, J.P.; Wilson, D.S.; Maguire, D.A.; Heath, L.S. Snag longevity under alternative silvicultural regimes in mixed-species forests of central Maine. Can. J. For. Res. 2005, 35, 787–796. [Google Scholar] [CrossRef]
- Lee, Y.J. Predicting mortality for even-aged stands of lodgepole pine. For. Chron. 1971, 47, 29–32. [Google Scholar] [CrossRef]
- Hamilton, D.A. A logistic model of mortality in thinned and unthinned mixed conifer stands of northern Idaho. For. Sci. 1986, 32, 989–1000. [Google Scholar]
- Fortin, M.; Bedard, S.; DeBlois, J.; Meunier, S. Predicting individual tree mortality in northern hardwood stands under uneven-aged management in southern Quebec, Canada. Ann. For. Sci. 2008, 65, 12. [Google Scholar] [CrossRef]
- Monserud, R.A.; Sterba, H. Modeling individual tree mortality for Austrian forest species. For. Ecol. Manag. 1999, 113, 109–123. [Google Scholar] [CrossRef]
- Eid, T.; Tuhus, E. Models for individual tree mortality in Norway. For. Ecol. Manag. 2001, 154, 69–84. [Google Scholar] [CrossRef]
- McGarrigle, E.; Kershaw, J.A.; Ducey, M.J.; Lavigne, M.B. A new approach to modeling stand-level dynamics based on informed random walks: Influence of bandwidth and sample size. Forestry 2013, 86, 377–389. [Google Scholar] [CrossRef]
- Davidson, C.B.; Gottschalk, K.W.; Johnson, J.E. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. For. Sci. 1999, 45, 74–84. [Google Scholar]
- Fosbroke, D.E.; Hicks, R.R., Jr. Tree Mortality Following Gypsy Moth Defoliation in Southwestern Pennsylvania. In Proceedings of the 7th Central Hardwood Conference, Carbondale, Illinois, USA, 1989; Rink, G., Budelsky, C.A., Eds.; USDA Forest Service: Carbondale, IL, USA; pp. 74–80.
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Frappier, B.; Eckert, R.T.; Lee, T.D. Experimental removal of the non-indigenous shrub Rhamnus frangula (glossy buckthorn): Effects on native herbs and woody seedlings. Northeast. Nat. 2004, 11, 333–342. [Google Scholar] [CrossRef]
- Burnham, K.M.; Lee, T.D. Canopy gaps facilitate establishment, growth, and reproduction of invasive Frangula alnus in a Tsuga canadensis dominated forest. Biol. Invasions 2010, 12, 1509–1520. [Google Scholar] [CrossRef]
- Brazee, N.J.; Wick, R.L. Armillaria species distribution on symptomatic hosts in northern hardwood and mixed oak forests in western Massachusetts. For. Ecol. Manag. 2009, 258, 1605–1612. [Google Scholar] [CrossRef]
- Brazee, N.J.; Marra, R.E.; Wick, R.L. Genotypic diversity of Armillaria gallica from mixed oak forests in Massachusetts. Mycologia 2012, 104, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Staley, J.M. Decline and mortality of red and scarlet oaks. For. Sci. 1965, 11, 2–17. [Google Scholar]
- Wargo, P.M. Consequences of environmental stress on oak: Predisposition to pathogens. Ann. Sci. For. 1996, 53, 359–368. [Google Scholar] [CrossRef]
- Houston, D.R. A Host-Stress-Saprogen Model for Forest Dieback-Decline Diseases. In Forest Decline Concepts; Manion, P.D., Lachance, D., Eds.; APS Press: St. Paul, MN, USA, 1992. [Google Scholar]
- Wargo, P.M. Armillariella mellea and Agrilus bilineatus and mortality of defoliated oak trees. For. Sci. 1977, 23, 485–492. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simmons, M.J.; Lee, T.D.; Ducey, M.J.; Dodds, K.J. Invasion of Winter Moth in New England: Effects of Defoliation and Site Quality on Tree Mortality. Forests 2014, 5, 2440-2463. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102440
Simmons MJ, Lee TD, Ducey MJ, Dodds KJ. Invasion of Winter Moth in New England: Effects of Defoliation and Site Quality on Tree Mortality. Forests. 2014; 5(10):2440-2463. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102440
Chicago/Turabian StyleSimmons, Michael J., Thomas D. Lee, Mark J. Ducey, and Kevin J. Dodds. 2014. "Invasion of Winter Moth in New England: Effects of Defoliation and Site Quality on Tree Mortality" Forests 5, no. 10: 2440-2463. https://0-doi-org.brum.beds.ac.uk/10.3390/f5102440