Ungulate Impact on Natural Regeneration in Spruce-Beech-Fir Stands in Černý důl Nature Reserve in the Orlické Hory Mountains, Case Study from Central Sudetes

Abstract
:1. Introduction
2. Materials and Method
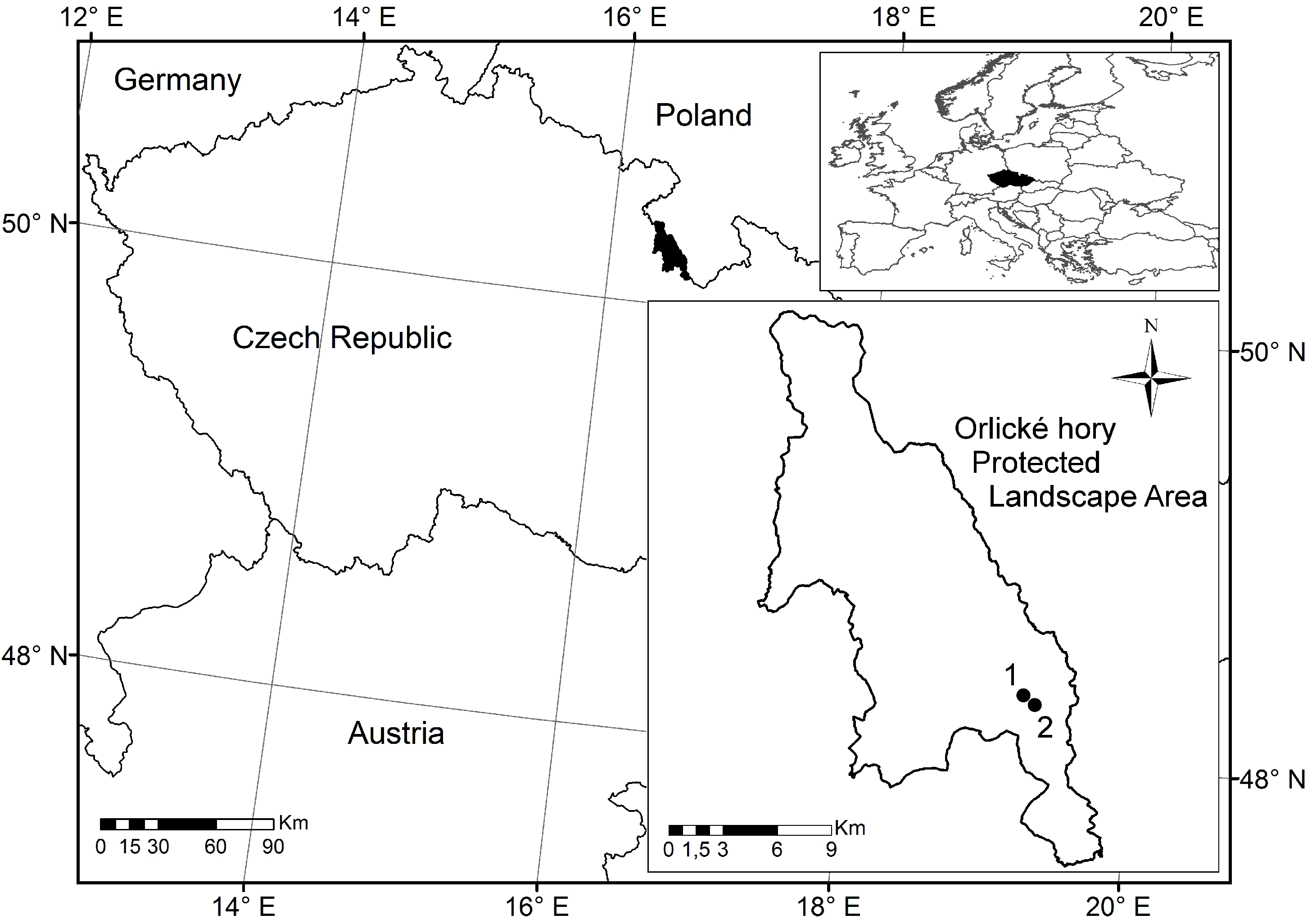
2.1. Area Description


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
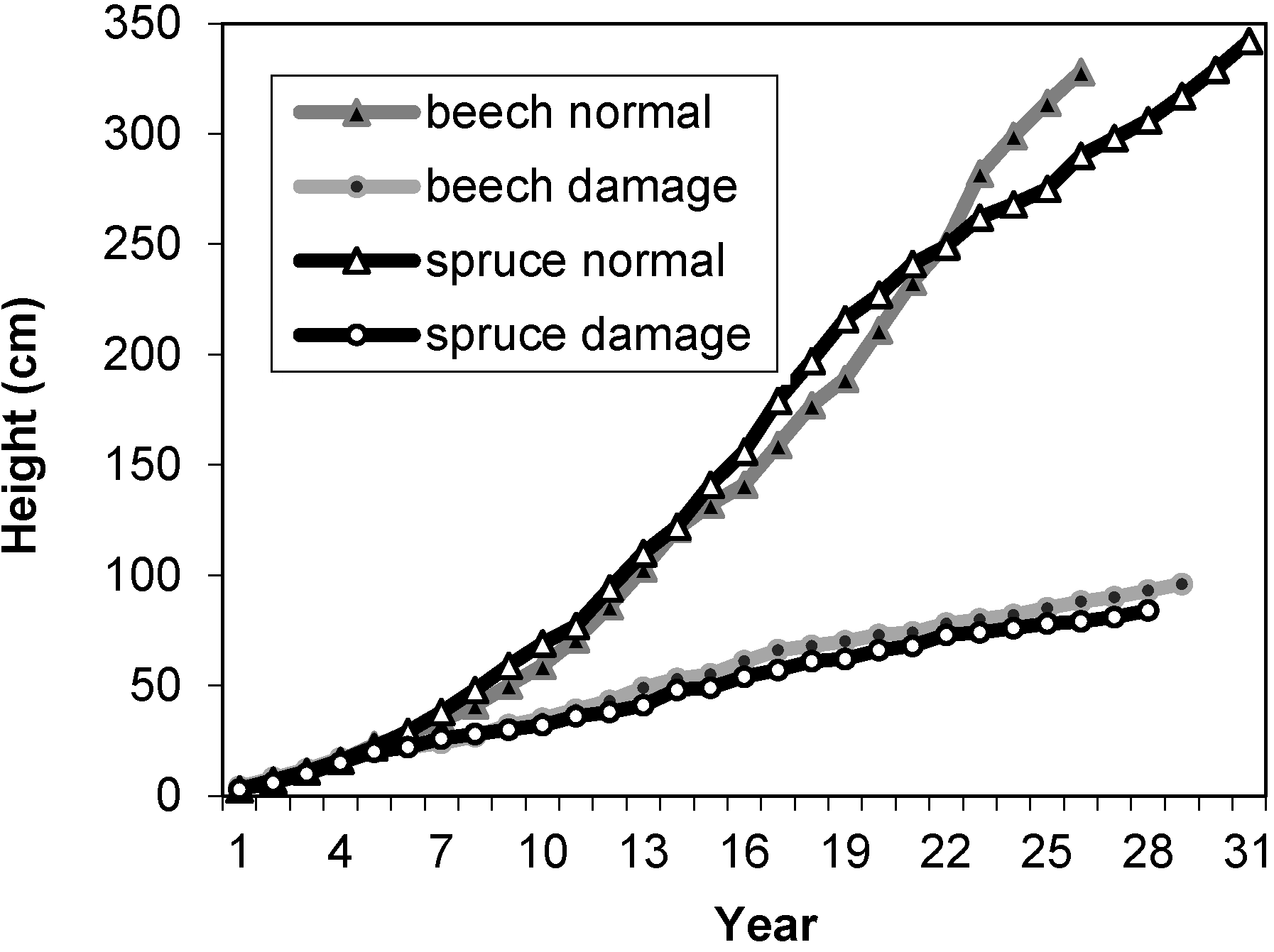{kind=link}
| Plot number | Plot name | Tree species | Height (m) | Diameter (cm) | Stand volume (m3.ha−1) | Stand basal area (m2.ha−1) | Age of tree layer | Canopy | Altitude (m) | Exposure | Slope (°) | Forest site type | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mensurational | Biological | ||||||||||||||
| Lower | Middle | Upper | |||||||||||||
| 1 | Černý důl fenced | beech | 28 | 47 | 181 | 16.8 | 53 | 99 | 185 | 0.98 | 1.87 | 835 | SE | 13 | 6S |
| spruce | 32 | 59 | 220 | 21.8 | |||||||||||
| fir | 33 | 65 | 78 | 7.0 | |||||||||||
| 2 | Černý důl unfenced | Beech | 31 | 53 | 299 | 22.7 | 53 | 99 | 185 | 0.97 | 1.88 | 820 | SE | 11 | 6S |
| spruce | 38 | 71 | 206 | 18.0 | |||||||||||
| fir | 38 | 72 | 50 | 3.4 | |||||||||||
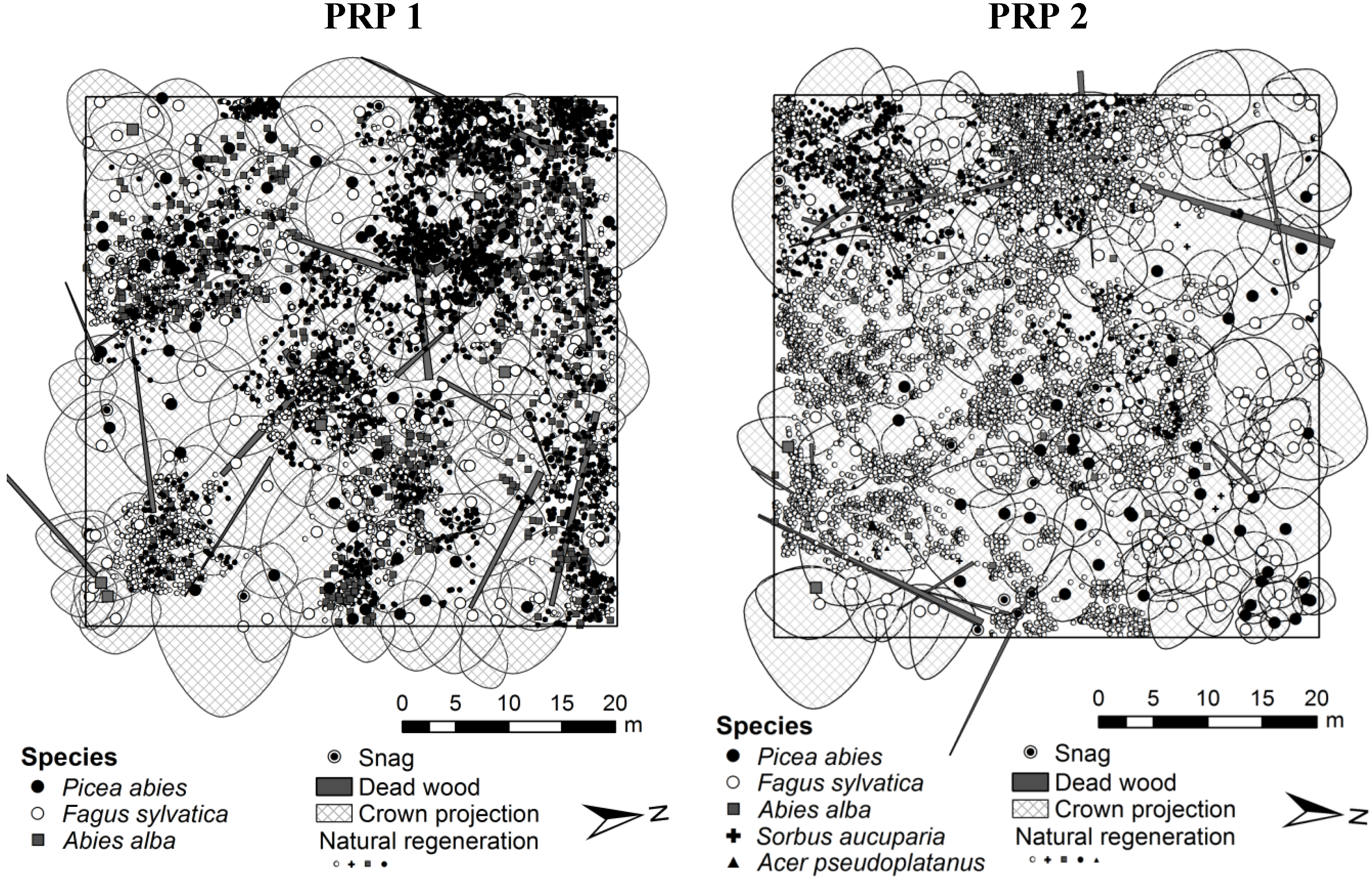
2.2. Data Collection
2.3. Data Analysis
| Index | Mean value | Aggregation | Regularity |
|---|---|---|---|
| Hopkins-Skellam | A = 0.5 | A > 0.5 | A < 0.5 |
| Pielou-Mountford | α = 1 | α > 1 | α < 1 |
| David-Moore | ICS = 0 | ICS > 0 | ICS < 0 |
| Clark-Evans | R = 1 | R < 1 | R > 1 |
3. Results
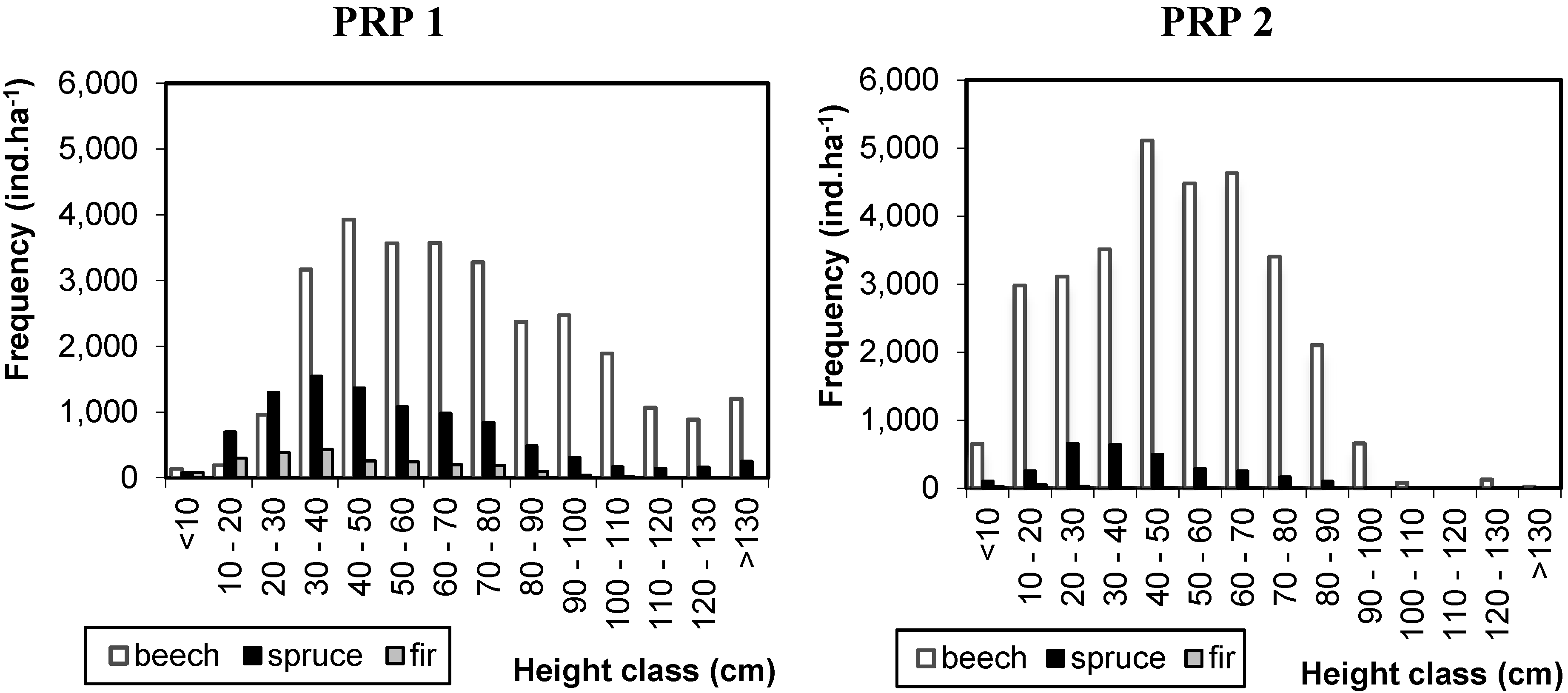
3.1. Structure of Natural Regeneration


| Index | Beech | Spruce | Fir | Rowan | all regeneration |
|---|---|---|---|---|---|
| Fenced PRP 1 | |||||
| Hopkins-Skellam | 0.980 * | 0.952 * | 0.922 * | 0.788 * | 0.841 * |
| Pielou-Mountford | 15.060 * | 11.566 * | 6.595 * | 2.316 * | 10.992 * |
| David-Moore | 28.400 * | 7.313 * | 1.871 * | −0.042 | 13.236 * |
| Clark-Evans | 0.572 * | 0.694 * | 0.702 * | 0.569 * | 0.742 * |
| Unfenced PRP 2 | |||||
| Hopkins-Skellam | 0.937 * | 0.956 * | 0.651 | 0.882 * | 0.892 * |
| Pielou-Mountford | 31.041 * | 11.132 * | 1.401 | 2.936 * | 14.132 * |
| David-Moore | 17.417 * | 7.983 * | 0.063 | 1.744 * | 12.192 * |
| Clark-Evans | 0.568 * | 0.536 * | 0.845 * | 0.281 * | 0.692 * |


3.2. Species Diversity
| PRP | species richness | species heterogeneity | species evenness | ||||
|---|---|---|---|---|---|---|---|
| m | D1 (Mai) | D2 (Mei) | λ (Sii) | H’ (Shi) | E1 (Pii) | E2 (Hii) | |
| 1 (fenced) | 4 | 0.283 | 0.020 | 0.439 | 0.756 | 0.545 | 0.882 |
| 2 (unfenced) | 5 | 0.287 | 0.021 | 0.169 | 0.328 | 0.237 | 0.349 |
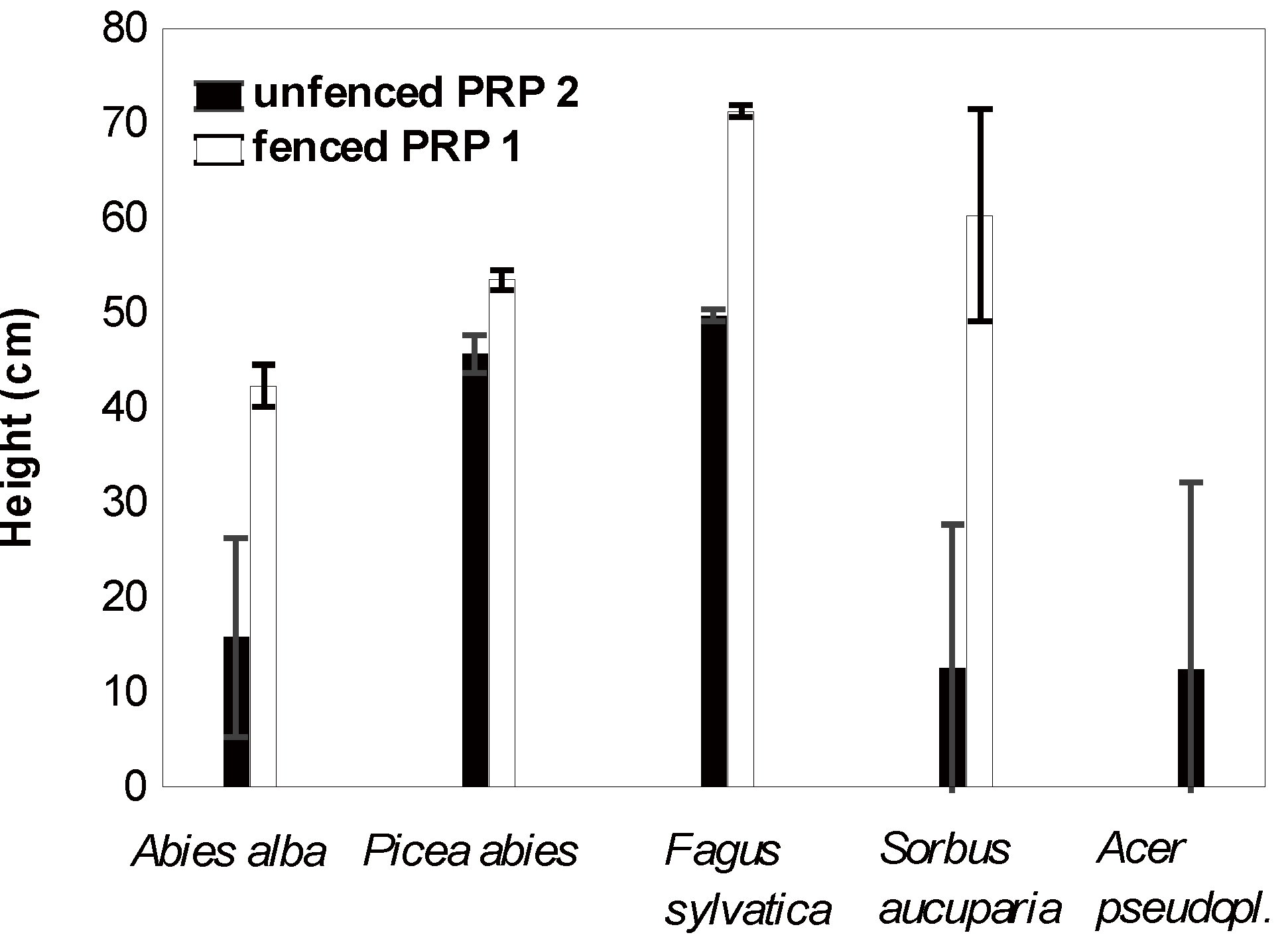
3.3. Impact of Ungulates on Natural Regeneration

| Type of shoot browsing | Beech | Spruce | Fir | Rowan | |||||
|---|---|---|---|---|---|---|---|---|---|
| ind. ha−1 | % | ind. ha−1 | % | ind. ha−1 | % | ind. ha−1 | % | ||
| Terminal | 1 time | 8,372 | 27.1 | 356 | 11.9 | 22 | 21.2 | 8 | 15.4 |
| 2–4 times | 6,737 | 21.8 | 217 | 7.3 | 26 | 25.0 | 17 | 32.7 | |
| 5+ times | 4,832 | 15.7 | 96 | 3.2 | 37 | 35.6 | 19 | 36.5 | |
| total | 19,941 | 64.6 | 669 | 22.4 | 85 | 81.7 | 44 | 84.6 | |
| Lateral | 1,945 | 45.2 | 416 | 13.9 | 81 | 77.9 | 31 | 59.6 | |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Veblen, T.T. Regeneration dynamics. In PlantSuccession, Theory and Prediction; Glenn-Lewin, D.C., Peet, R.K., Veblen, T.T., Eds.; Chapmann and Hall: London, UK, 1992; pp. 152–187. [Google Scholar]
- Korpeľ, Š. Die Urwälder der Westkarpaten; Gustav Fischer Verlag: Stuttgart, Jena, Germany, 1995; p. 310. [Google Scholar]
- Gratzer, G.; Canham, Ch.; Dieckmann, U.; Fischer, A.; Iwasa, Y.; Law, R.; Lexer, M.J.; Sandmann, H.; Spies, T.A.; Splechtna, B.E.; et al. Spatio-temporal development of forests-current trends in field methods and models. Oikos 2004, 107, 3–15. [Google Scholar] [CrossRef]
- Jeník, J. Ecological meaning of stability. In Stability of Spruce Forest Ecosystems; Symposium MAB: Brno, Czech Republic, 1979; pp. 7–15. [Google Scholar]
- Paluch, J.G. The spatial pattern of a natural European beech (Fagus sylvatica L.)-silver fir (Abies alba Mill.) forest: A patch mosaic perspective. For. Ecol. Manag. 2007, 253, 161–170. [Google Scholar] [CrossRef]
- Šamonil, P.; Vrška, T. Long term vegetation dynamics in the Šumava Mts. Natural spruce-fir-beech forests. Plant Ecol. 2008, 196, 197–214. [Google Scholar] [CrossRef]
- Brang, P. Virgin forests as a knowledge source for central European silviculture: Reality or myth? For. Snow Landsc. Res. 2005, 79, 19–32. [Google Scholar]
- Motta, R. Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the eastern Italian Alps. For. Ecol. Manag. 2003, 181, 139–150. [Google Scholar] [CrossRef]
- Senn, J.; Suter, W. Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: Beliefs in search of supporting data. For. Ecol. Manag. 2003, 181, 151–164. [Google Scholar] [CrossRef]
- Motta, R. Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For. Ecol. Manag. 1996, 88, 93–98. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Zimmermann, S.; Bugmann, H. Browsing regime and growth response of naturally regenerated Abies alba saplings along light gradients. For. Ecol. Manag. 2013, 310, 393–404. [Google Scholar] [CrossRef]
- Čermák, P.; Horsák, P.; Špiřík, M.; Mrkva, R. Relationships between browsing damage and woody species dominance. J. For. Sci. 2009, 55, 23–31. [Google Scholar]
- Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Jr. (Ed.) Cambridge University Press: Cambridge, UK, 1999; p. 714.
- Tabaku, V. Struktur von Buchen-Urwäldern in Albanien im Vergleich mit deutschen Buchen-Naturwaldreservaten und Wirtschaftswäldern; Cuvillier Verlag: Göttingen, Germany, 2000; p. 206. [Google Scholar]
- Götmark, F. Careful partial harvesting in conservation stands and retention of large oaks favour oak regeneration. Biol. Conserv. 2007, 140, 349–358. [Google Scholar] [CrossRef]
- Zlatník, A. Chráněná území, zvláště rezervace a jejich hospodářské a společenské funkce v krajině. Lesnictví 1970, 16, 857–867. [Google Scholar]
- Průša, E. Die Böhmischen und Mährischen Urwälder; Vegetace ČSSR, A15; Academia: Praha, Czech Republic, 1985; p. 577. [Google Scholar]
- Lindh, B.C.; Muir, P.S. Understory vegetation in young Douglas-fir forests: Does thinning help restore old-growth composition? For. Ecol. Manag. 2004, 192, 285–296. [Google Scholar] [CrossRef]
- Decocq, G.; Aubert, M.; Dupont, F.; Alard, D.; Saquez, R.; Wattez-Franger, A.; de Foucaly, B.; Delelis-Dusollier, A.; Bardat, J. Plant diversity in a managed temperate deciduous forest: Understorey response to two silvicultural systems. J. Appl. Ecol. 2004, 41, 1065–1079. [Google Scholar] [CrossRef]
- Bebber, D.P.; Cole, W.G.; Thomas, S.C.; Balsillie, D.; Duinker, P. Effects of retention harvests on structure of old-growth Pinus strobus L. stands in Ontario. For. Ecol. Manag. 2005, 205, 91–103. [Google Scholar] [CrossRef]
- Götmark, F. Experiments for alternative management of forest reserves: Effects of partial cutting on stem growth and mortality of large oaks. Can. J. For. Res. 2009, 39, 1322–1330. [Google Scholar] [CrossRef]
- Heinken, T.; Raudnitschka, D. Do wild ungulates contribute to the dispersal of vascular plants in Central European forests by epizoochory? A case study in NE Germany. Forstwiss. Cent. 2002, 121, 179–194. [Google Scholar] [CrossRef]
- Ammer, C. Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. For. Ecol. Manag. 1996, 88, 43–53. [Google Scholar] [CrossRef]
- Rooney, T.P. Deer impacts on forest ecosystems: A North American perspective. Forestry 2001, 74, 201–208. [Google Scholar] [CrossRef]
- Vrška, T.; Hort, L.; Odehnalová, P.; Adam, D.; Horal, D. Boubín virgin forest after 24 years (1972–1996)—Development of tree layer. J. For. Sci. 2001, 47, 439–456. [Google Scholar]
- Diaci, J.; Rozenbergar, D.; Boncina, A. Stand dynamics of Dinaric old-growth forest in Slovenia: Are indirect human influences relevant? Plant Biosyst. 2010, 144, 194–201. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Remeš, J.; Štefančík, I.; Bulušek, D.; Bílek, L. Struktura a vývoj lesních porostů v NPR Trčkov–CHKO Orlické hory, Česká republika. Lesnícký časopis–For. J. 2013, 59, 249–264. [Google Scholar]
- Suzuki, M.; Miyashita, T.; Kabaya, H.; Ochiai, K.; Asada, M.; Kikvidze, Z. Deer herbivory as an important driver of divergence of ground vegetation communities in temperate forests. Oikos 2013, 122, 104–110. [Google Scholar] [CrossRef]
- Heuze, P.; Schnitzler, A.; Klein, F. Consequences of increased deer browsing winter on silver fir and spruce regeneration in the Southern Vosges mountains: Implications for forest management. Ann. For. Sci. 2005, 62, 175–181. [Google Scholar] [CrossRef]
- Cagnacci, F.; Focardi, S.; Heurich, M.; Stache, A.; Hewison, A.J.M.; Morellet, N.; Kjellander, P.; Linnell, J.D.C.; Mysterud, A.; Neteler, M.; et al. Partial migration in roe deer: Migratory and resident tactics are end points of a behavioural gradient determined by ecological factors. Oikos 2011, 120, 1790–1802. [Google Scholar] [CrossRef]
- Vacek, S.; Podrázský, V.; Souček, J. Dynamika poškození smrkových a bukových porostů v CHKO Orlické hory. VII. Analýza změn v NPR Bukačka a NPR Trčkov. Příroda 1997, 11, 183–199. [Google Scholar]
- Hofmeister, Š.; Svoboda, M.; Souček, J.; Vacek, S. Spatial pattern of Norway spruce and silver fir natural regeneration in uneven-aged mixed forests of northeastern Bohemia. J. For. Sci. 2008, 54, 92–101. [Google Scholar]
- Firm, D.; Nagel, T.A.; Diaci, J. Disturbance history and dynamics of an old-growth mixed species mountain forest in the Slovenian Alps. For. Ecol. Manag. 2009, 257, 1893–1901. [Google Scholar] [CrossRef]
- Vrška, T.; Adam, D.; Hort, L.; Kolár, T.; Janík, D. European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) rotation in the Carpathians—A developmental cycle or a linear trend induced by man? For. Ecol. Manag. 2009, 258, 347–356. [Google Scholar] [CrossRef]
- Klopcic, M.; Jerina, K.; Boncina, A. Long-term changes of structure and tree species composition in Dinaric uneven-aged forests: Are red deer an important factor? Eur. J. For. Res. 2010, 129, 277–288. [Google Scholar] [CrossRef]
- Mikyška, R. Die Wälder der Böhmischen Mittleren Sudeten und Ihrer Vorberge; Rozpravy ČSAV, Řada Matematických a Přírodních věd; Academia: Praha, Czech Republic, 1972; Volume 82, p. 162. [Google Scholar]
- Wagnerová, Z. Inventarizační průzkum flóry a vegetace státní přírodní rezervace” Černý důl” v Orlických horách. Příroda 1976, 8, 93–108. [Google Scholar]
- Vacek, S.; Simon, J.; Remeš, J.; Podrázský, V.; Minx, T.; Mikeska, M.; Malík, V.; Jankovský, L.; Turčáni, M.; Jakuš, R.; Schwarz, O.; Kozel, J.; Valenta, M.; Lička, D.; Hlásny, T.; Zúbrik, M.; Krejčí, F.; Třešňák, J.; Hofmeister, Š. Obhospodařování Bohatě Strukturovaných a Přírodě Blízkých Lesů; Lesnická práce, s. r. o.: Kostelec nad Černými lesy, Czech Republic, 2007; p. 447. [Google Scholar]
- Hopkins, B.; Skellam, J.G. A new method for determining the type of distribution of plant individuals. Ann. Bot. 1954, 18, 213–227. [Google Scholar]
- Pielou, E.C. The use of point-to-plant distances in the study of the pattern of plant populations. J. Ecol. 1959, 47, 607–613. [Google Scholar] [CrossRef]
- Mountford, M.D.; On, E.C. Pielou’s index of nonrandomness. J. Ecol. 1961, 49, 271–275. [Google Scholar] [CrossRef]
- Clark, P.; Evans, F.C. Distance to nearest neighbour as a measure of spatial relationship in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Ripley, B.D. Spatial Statistics, 1st ed.; John Wiley & Sons: New York, NY, USA, 1981; p. 252. [Google Scholar]
- David, F.N.; Moore, P.G. Notes on contagious distributions in plant populations. Ann. Bot. Lond. 1954, 18, 47–53. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Menhinick, C.F. A comparison of some species-individuals diversity indices applied to samples of field insects. Ecology 1964, 45, 859–861. [Google Scholar] [CrossRef]
- Simpson, H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communications. Bell Syst. Technical J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evencess: A unifying notation and is consequenses. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity, 1st ed.; Wiley: New York, NY, USA, 1975; p. 165. [Google Scholar]
- Bottero, A.; Garbarino, M.; Dukic, V.; Govedar, Z.; Lingua, E.; Nagel, T.A.; Motta, R. Gapphase dynamics in the old-growth forest of Lom, Bosnia and Herzegovina. Silva Fenn. 2011, 45, 875–887. [Google Scholar] [CrossRef]
- Štefančík, I. Changes in tree species composition, stand structure, qualitative and quantitative production of mixed spruce, fir and beech stand on Stará Píla research plot. J. For. Sci. 2006, 52, 74–89. [Google Scholar]
- Jaworski, A.; Kolodziej, Z.B.; Porada, K. Structure and dynamics of stands of primeval character in selected areas of the Bieszczady National Park. J. For. Sci. 2002, 48, 185–201. [Google Scholar]
- Saniga, M.; Zrak, J.; Pittner, J.; Balanda, M. Štruktúra, Produkcia, Regeneračné Procesy a Disturbačný Režim Prírodného Lesa v NPR Skalná Alpa; Technická Univerzita: Zvolen, Slovakia, 2013; p. 68. [Google Scholar]
- Saniga, M. Štruktúra, Produkčné a Regeneračné Procesy Dobročského Pralesa; Vedecké štúdie, 2/A; Technická Univerzita: Zvolen, Slovakia, 1999; p. 64. [Google Scholar]
- Šebková, B.; Šamonil, P.; Janík, D.; Adam, D.; Král, K.; Vrška, T.; Hort, L.; Unar, P. Spatial and volume patterns of an unmanaged submontane mixed forest in Central Europe: 160 years of spontaneous dynamics. For. Ecol. Manag. 2011, 262, 873–885. [Google Scholar] [CrossRef]
- Janik, D.; Adam, D.; Hort, L.; Král, K.; Šamonil, P.; Unar, P.; Vrška, T.; Horal, D. Spatiotemporal differences in tree spatial patterns between alluvial hardwood and mountain fir-beech forests: Do characteristic patterns exist? J. Veg. Sci. 2013, 24, 1141–1153. [Google Scholar] [CrossRef]
- Szymura, T.H.; Dunajski, A.; Aman, I.; Makowski, M.; Szymura, M. The spatial pattern and microsites requirements of Abies alba natural regeneration in the Karkonosze Mountains. Dendrobiology 2007, 58, 51–57. [Google Scholar]
- Peck, J.L.E.; Zenner, E.K. Spatial patterns of natural Pinus strobus L. regeneration in a Pinus resinosa Ait. stand. J. Torrey Bot. Soc. 2009, 136, 369–379. [Google Scholar] [CrossRef]
- Vacek, S.; Moucha, P. Péče o Lesní Ekosystémy v Chráněných Územích ČR; Ministerstvo životního prostředí: Praha, Czech Republic, 2012; p. 896. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Remeš, J.; Bulušek, D.; Králíček, I. Ungulate Impact on Natural Regeneration in Spruce-Beech-Fir Stands in Černý důl Nature Reserve in the Orlické Hory Mountains, Case Study from Central Sudetes. Forests 2014, 5, 2929-2946. https://0-doi-org.brum.beds.ac.uk/10.3390/f5112929
Vacek Z, Vacek S, Bílek L, Král J, Remeš J, Bulušek D, Králíček I. Ungulate Impact on Natural Regeneration in Spruce-Beech-Fir Stands in Černý důl Nature Reserve in the Orlické Hory Mountains, Case Study from Central Sudetes. Forests. 2014; 5(11):2929-2946. https://0-doi-org.brum.beds.ac.uk/10.3390/f5112929
Chicago/Turabian StyleVacek, Zdeněk, Stanislav Vacek, Lukáš Bílek, Jan Král, Jiří Remeš, Daniel Bulušek, and Ivo Králíček. 2014. "Ungulate Impact on Natural Regeneration in Spruce-Beech-Fir Stands in Černý důl Nature Reserve in the Orlické Hory Mountains, Case Study from Central Sudetes" Forests 5, no. 11: 2929-2946. https://0-doi-org.brum.beds.ac.uk/10.3390/f5112929