
Genetic Diversity and Population Structure of Toona Ciliata Roem. Based on Sequence-Related Amplified Polymorphism (SRAP) Markers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Code | Province | Latitude (°′ N) | Longitude (°′ E) | Sample Number |
|---|---|---|---|---|---|
| 1 | YF | Guangdong | 22°46′ | 111°34′ | 15 |
| 2 | LC | Guangdong | 25°07′ | 113°20′ | 20 |
| 3 | JX | Anhui | 30°41′ | 118°24′ | 30 |
| 4 | HS | Anhui | 30°16′ | 118°08′ | 6 |
| 5 | CH | Guizhou | 24°59′ | 105°48′ | 30 |
| 6 | GL | Guizhou | 25°52′ | 105°37 | 30 |
| 7 | WM | Guizhou | 25°10′ | 106°05′ | 30 |
| 8 | LD | Guizhou | 25°25′ | 106°44′ | 30 |
| 9 | XY | Guizhou | 25°06′ | 104°54′ | 30 |
| 10 | PP | Yunnan | 25°04′ | 99°06′ | 30 |
| 11 | PE | Yunnan | 22°46′ | 100°58′ | 30 |
| 12 | PW | Yunnan | 22°23′ | 101°04′ | 30 |
| 13 | YR | Yunnan | 25°01′ | 101°32′ | 30 |
| 14 | GS | Jiangxi | 27°19′ | 115°26′ | 30 |
| 15 | JLS | Jiangxi | 24°54′ | 114°47′ | 30 |
| 16 | JGS | Jiangxi | 26°44′ | 114°17′ | 30 |
| 17 | WYS | Jiangxi | 28°18′ | 117°42′ | 30 |
| 18 | MTS | Jiangxi | 27°41′ | 117°03′ | 30 |
| 19 | HPS | Hunan | 29°01′ | 111°41′ | 30 |
| 20 | CB | Hunan | 27°14′ | 111°28′ | 30 |
| 21 | XN | Hunan | 28°18′ | 109°44′ | 32 |
| 22 | TL | Guangxi | 24°17′ | 106°13′ | 30 |
| 23 | LL | Guangxi | 24°46′ | 105°20′ | 30 |
| 24 | XL | Guangxi | 24°29′ | 105°05′ | 30 |
| 25 | XJ | Zhejiang | 28°45′ | 119°92′ | 30 |
| 26 | SC | Zhejiang | 28°59′ | 119°25′ | 30 |
| 27 | NP | Fujian | 26°38′ | 118°10′ | 30 |
| 28 | SP | Hubei | 30°17′ | 109°28′ | 30 |
| 29 | HD | Sichuan | 27°23′ | 102°09′ | 30 |
| 30 | DC | Sichuan | 26°40′ | 102°32′ | 30 |
2.2. DNA Extraction
2.3. SRAP Analysis
| Primer | Primer Combination Sequences | |
|---|---|---|
| Me17Em21 | F:5’TGA GTA CAA ACC GG TAG 3’ | R:5’GAC TGC GTA CGA ATT TCG 3’ |
| Me19Em13 | F:5’TGA GTA CAA ACC GG TTG 3’ | R:5’GAC TGC GTA CGA ATT CTA 3’ |
| Me19Em15 | F:5’TGA GTA CAA ACC GG TTG 3’ | R:5’GAC TGC GTA CGA ATT CTT 3’ |
| Me19Em27 | F:5’TGA GTA CAA ACC GG TTG 3’ | R:5’GAC TGC GTA CGA ATT CCA 3’ |
| Me22Em21 | F:5’TGA GTA CAA ACC GG GTC 3’ | R:5’GAC TGC GTA CGA ATT CCA 3’ |
| Me22Em27 | F:5’TGA GTA CAA ACC GG GTC 3’ | R:5’GAC TGC GTA CGA ATT CCA 3’ |
| Me23Em22 | F:5’TGA GTA CAA ACC GG AGT 3’ | R:5’GAC TGC GTA CGA ATT GTC 3’ |
| Me23Em31 | F:5’TGA GTA CAA ACC GG AGT 3’ | R:5’GAC TGC GTA CGA ATT CTCA 3’ |
| Me27Em28 | F:5’TGA GTA CAA ACC GG GAT 3’ | R:5’GAC TGC GTA CGA ATT TGA 3’ |
| Me27Em7 | F:5’AGC GAG CAA GCC GG GAT 3’ | R:5’GAC TGC GTA CGA ATT GAG 3’ |
| Me28Em18 | F:5’GAC CAG TAA ACC GG TGG 3’ | R:5’GAC TGC GTA CGAATT AGC 3’ |
| Me31Em24 | F:5’TGA GTA CAA ACC GG ATG 3’ | R:5’GAC TGC GTA CGA ATT CAG 3’ |
| Me32Em17 | F:5’GAG CGT CGA ACC GG GAA 3’ | R:5’GAC TGC GTA CGAATTCAG 3’ |
| Me32Em26 | F:5’GTA CAT AGA ACC GG GAA 3’ | R:5’GAC TGC GTA CGA ATT CAG 3’ |
| Me34Em13 | F:5’GTA CAT AGA ACC GG TAT 3’ | R:5’GAC TGC GTA CGAATT CAG 3’ |
| Me35Em13 | F:5’GTA CAT AGA ACC GG ATG 3’ | R:5’GAC TGC GTA CGAATT CAG 3’ |
| Me35Em14 | F:5’TAC GAC GAA TCC GG ATG 3’ | R:5’GAC TGC GTA CGAATT CAG 3’ |
| Me35Em32 | F:5’CAC AGT CAT GCC GG ATG 3’ | R:5’GAC TGC GTA CGA ATT ATT 3’ |
| Me38Em21 | F:5’CAC AGT CAT GCC GG AGT 3’ | R:5’GAC TGC GTA CGAATT ATT 3’ |
| Me38Em24 | F:5’ATC AGT CGG ACC GG AGT 3’ | R:5’GAC TGC GTA CGAATT ATT 3’ |
| Me39Em32 | F:5’GTA CAT AGA ACC GG ACT 3’ | R:5’GAC TGC GTA CGAATT ATT 3’ |
| Me40Em13 | F:5’TAC GAC GAA TCC GG ACT 3’ | R:5’GAC TGC GTA CGA ATT CTA 3’ |
| Me40Em22 | F:5’CAC AGT CAT GCC GG ACT 3’ | R:5’GAC TGC GTA CGAATT GTC 3’ |
| Me43Em19 | F:5’CAC AGT CAT GCC GG ATT 3’ | R:5’GAC TGC GTA CGAATT ACG 3’ |
2.4. Data Analyses
3. Results
3.1. SRAP Fragments Amplified Polymorphism Analysis
| Primer Combinations | Total No. of Bands | Polymorphic Bands | Polymorphism (%) | PIC |
|---|---|---|---|---|
| Me17Em21 | 27 | 21 | 77.78 | 0.38 |
| Me19Em13 | 25 | 21 | 84.00 | 0.42 |
| Me19Em15 | 26 | 23 | 88.46 | 0.42 |
| Me19Em27 | 25 | 21 | 84.00 | 0.41 |
| Me22Em21 | 26 | 21 | 80.77 | 0.41 |
| Me22Em27 | 28 | 20 | 71.43 | 0.41 |
| Me23Em22 | 30 | 26 | 86.67 | 0.44 |
| Me23Em31 | 28 | 21 | 75.00 | 0.41 |
| Me27Em28 | 25 | 21 | 84.00 | 0.39 |
| Me27Em7 | 25 | 21 | 84.00 | 0.44 |
| Me28Em18 | 36 | 27 | 75.00 | 0.40 |
| Me31Em24 | 36 | 29 | 80.56 | 0.43 |
| Me32Em17 | 28 | 20 | 71.43 | 0.43 |
| Me32Em26 | 32 | 28 | 87.50 | 0.42 |
| Me34Em13 | 25 | 19 | 76.00 | 0.39 |
| Me35Em13 | 27 | 15 | 55.56 | 0.42 |
| Me35Em14 | 26 | 15 | 57.69 | 0.40 |
| Me35Em32 | 27 | 14 | 51.85 | 0.45 |
| Me38Em21 | 24 | 19 | 79.17 | 0.39 |
| Me38Em24 | 29 | 21 | 72.41 | 0.42 |
| Me39Em32 | 19 | 18 | 94.74 | 0.43 |
| Me40Em13 | 28 | 23 | 82.14 | 0.42 |
| Me40Em22 | 21 | 15 | 71.43 | 0.33 |
| Me43Em19 | 33 | 26 | 78.79 | 0.43 |
| Total | 656 | 505 | 1850.36 | |
| Mean | 27.33 | 21.04 | 77.10 | 0.41 |
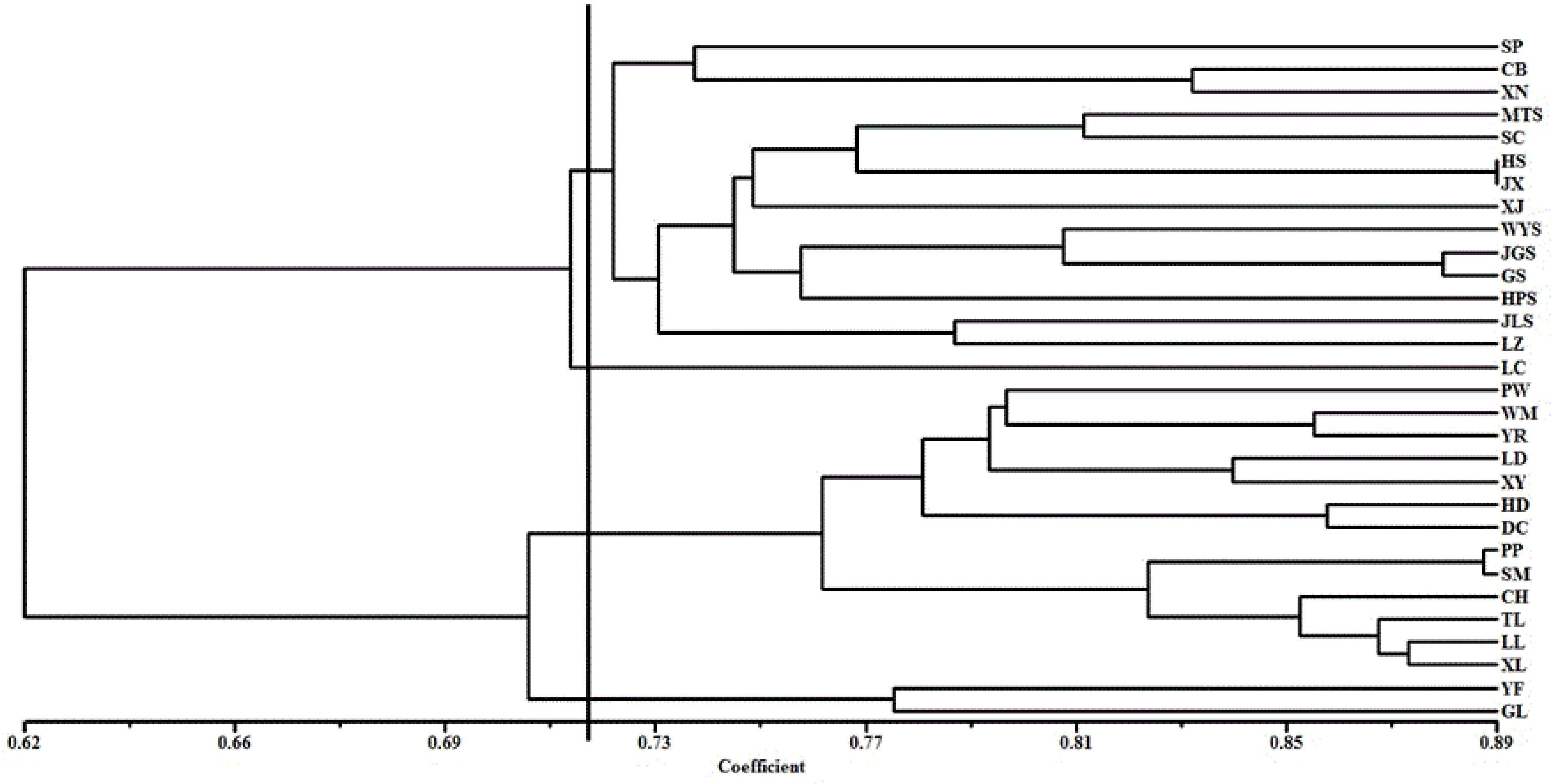
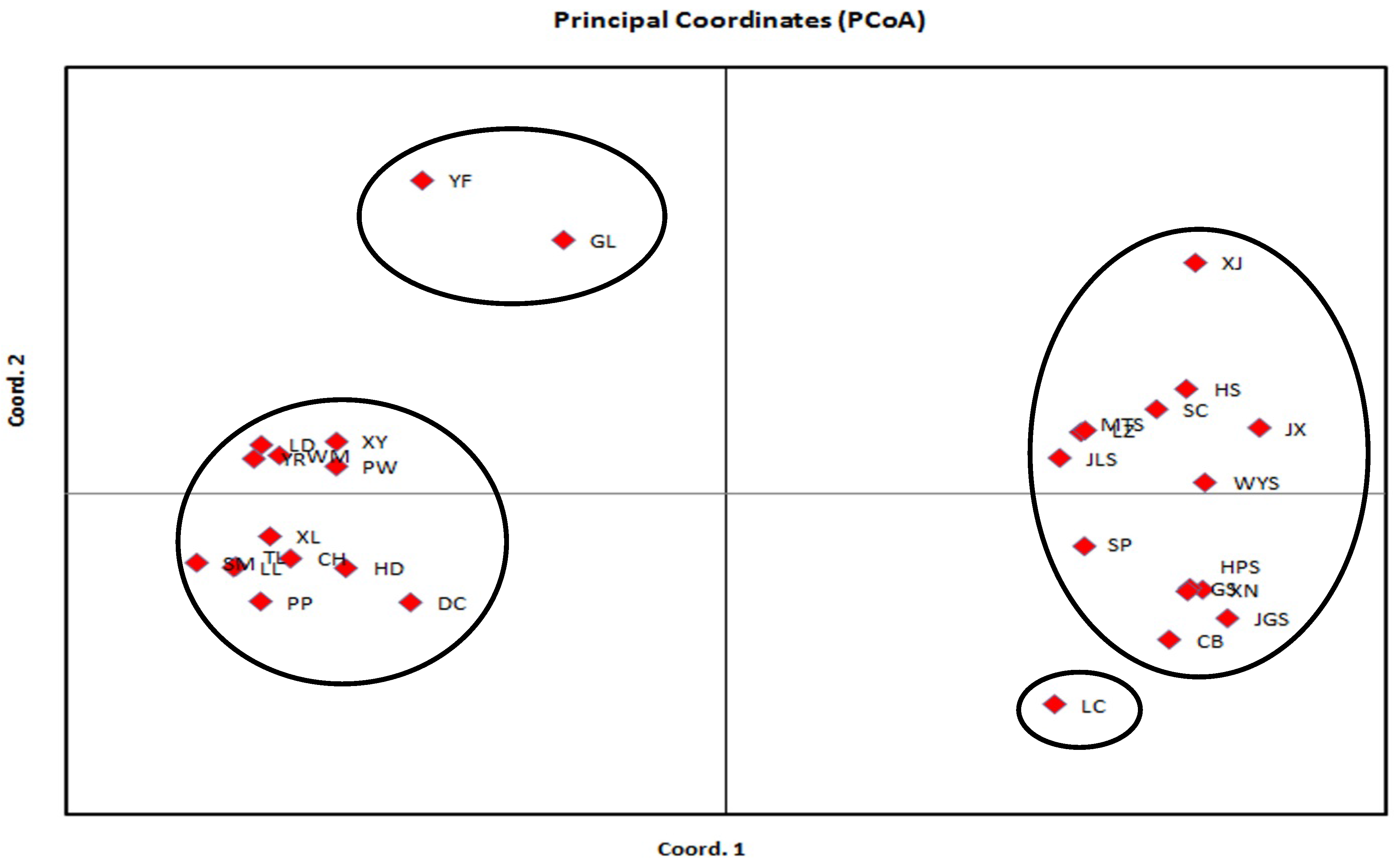
3.2. Genetic Relationship Analysis Based on SRAP

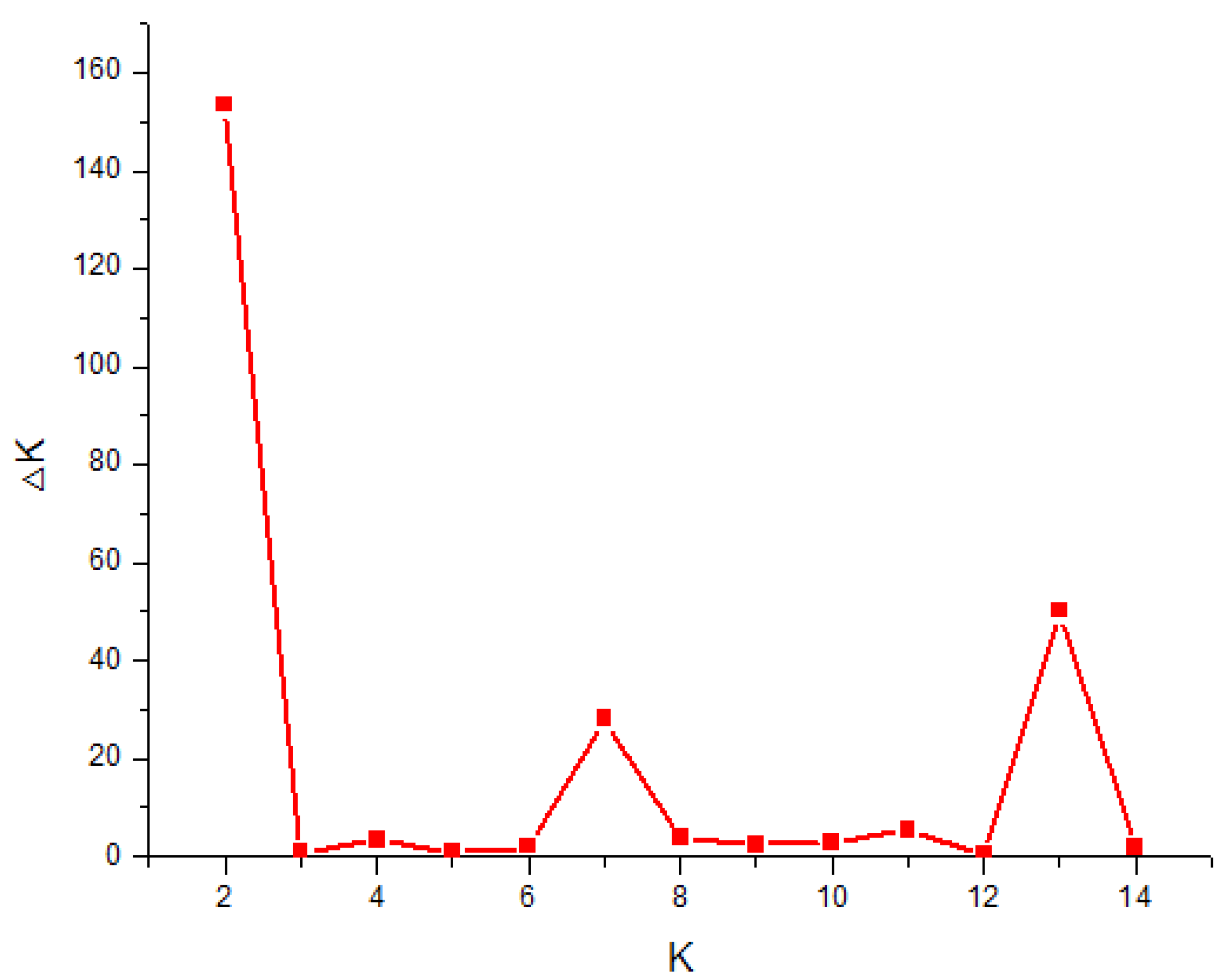
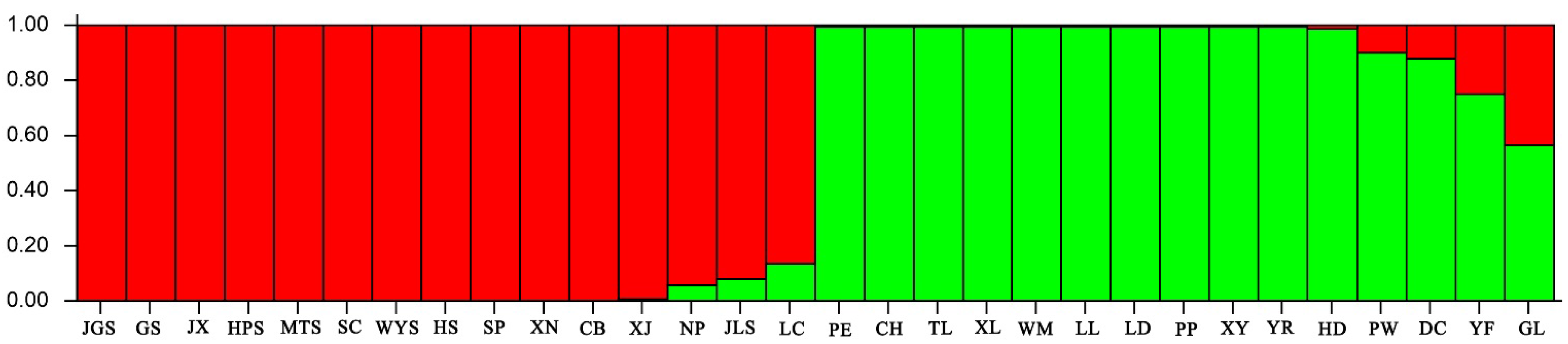
3.3. Genetic Structure Analysis
3.4. AMOVA Analysis

| 1 | 2 | 3 | 4 | |
|---|---|---|---|---|
| 1 | **** | |||
| 2 | 0.3347 | **** | ||
| 3 | 0.2321 | 0.5383 | **** | |
| 4 | 0.3038 | 0.6408 | 0.2070 | **** |


| Source | df | Sum of Squares | Mean Squares | Variation Component | Percentage of Variance (%) | Φst | P |
|---|---|---|---|---|---|---|---|
| Among Pops | 29 | 40,273.140 | 1388.729 | 81.979 | 79.24% | 0.792 | 0.001 |
| Within Pops | 475 | 10,197.632 | 21.469 | 21.469 | 20.76% | ||
| Total | 504 | 50,470.772 | 103.448 | 100% |
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, L.W.; Shi, Q.; Liang, G.; Cai, L.; He, G.Z. Tissue culture of precious timber species Toona ciliata Roem. Subtrop. Plant Sci. 2014, 43, 164–167. [Google Scholar]
- Liu, J.; Sun, Z.X.; Chen, Y.T.; Jiang, J.M. Isolation and characterization of microsatellite loci from an endangered tree species, Toona ciliata var. pubescens. Genet. Mol. Res. 2012, 11, 4411–4417. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.L.; Liao, R.Y.; Dai, J. Endangered causes and protection strategy of Toona ciliata. Guangxi For. Sci. 2011, 40, 201–203. [Google Scholar]
- Wen, W.H.; Wu, J.Y.; Chen, M.G.; Chen, J.F.; Cheng, Y.; Wang, X.J.; Liu, Q. Seedling growth performance of Toona ciliate elite trees progeny. Chin. Agric. Sci. Bull. 2012, 28, 36–39. [Google Scholar]
- Zhou, G.S. The study of introdution and cultivation on valuable fast-growing species of Toona ciliata and Toona ciliata var. J. Fujian Coll. For. 1994, 14, 271–276. [Google Scholar]
- Liu, J.; Jiang, J.M.; Chen, Y.T. Genetic diversity of central and peripheral populations of Toona ciliata var. pubescens, an endangered tree species endemic to China. Genet. Mol. Res. 2014, 13, 4579–4590. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Zhang, L.; Liao, C.K. Seed rain, soil seed bank, and natural regeneration of natural Toona ciliata var. pubescens forest. Ying Yong Sheng Tai Xue Bao 2012, 23, 972–978. [Google Scholar] [PubMed]
- Zhang, Z.R. Studied on Essential System of Forest Law of China. Master’s Thesis, Northeast Foresty University, Harbin, China, March 2004. [Google Scholar]
- Cheng, D.S.; Cui, T.L. Utilization Value and Cultivation Techniques of Toona sureni. For. Prod. Spec. China 2010, 4, 39–40. [Google Scholar]
- Jones, N.; Ougham, H.; Thomas, H.; Pasakinskiene, I. Markers and mapping revisited: Finding your gene. New Phytol. 2009, 183, 935–966. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Gao, W.; Lin, S.; Zhu, J.; Xie, B.; Lin, Z. Analysis of genetic diversity in ganoderma population with a novel molecular marker SRAP. Appl. Microbiol. Biotechnol. 2006, 72, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Quiros, C. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. Theor. Appl. Genet. 2001, 103, 455–461. [Google Scholar] [CrossRef]
- Feng, N.; Xue, Q.; Guo, Q.; Zhao, R.; Guo, M. Genetic diversity and population structure of celosia argentea and related species revealed by SRAP. Biochem. Genet. 2009, 47, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, Y.; Li, H.; Zhao, S.; Wang, S.; Liu, Y.; Wu, D.; Xu, F. Genetic diversity of pleurotus pulmonarius revealed by RAPD, ISSR, and SRAP fingerprinting. Curr. Microbiol. 2014, 68, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Jing, Z.B.; Wang, X.P.; Cheng, J.M. Analysis of genetic diversity among chinese wild vitis species revealed with SSR and SRAP markers. Genet. Mol. Res. 2013, 12, 1962–1973. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S.; Al-Faifi, S.; Migdadi, H.; Khan, M.; EL-Harty, E.; Ammar, M. Molecular diversity assessment using sequence related amplified polymorphism (srap) markers in Vicia faba L. Int. J. Mol. Sci. 2012, 13, 16457–16471. [Google Scholar] [CrossRef] [PubMed]
- Aneja, B.; Yadav, N.R.; Chawla, V.; Yadav, R.C. Sequence-related amplified polymorphism (SRAP) molecular marker system and its applications in crop improvement. Mol. Breed. 2012, 30, 1635–1648. [Google Scholar] [CrossRef]
- Huang, C.; Liu, G.; Bai, C.; Wang, W. Genetic analysis of 430 Chinese cynodon dactylon accessions using sequence-related amplified polymorphism markers. Int. J. Mol. Sci. 2014, 15, 19134–19146. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, N.; Rahimmalek, M.; Talebi, M.; Khorrami, M. Assessment of genetic diversity among and within carthamus species using sequence-related amplified polymorphism (SRAP) Markers. Plant Syst. Evol. 2013, 299, 1285–1294. [Google Scholar] [CrossRef]
- Youssef, M.; James, A.C.; Rivera-Madrid, R.; Ortiz, R.; Escobedo-GraciaMedrano, R.M. Musa genetic diversity revealed by SRAP and AFLP. Mol. Biotechnol. 2011, 47, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Liu, J.; Yang, D.; Chen, J.; Luan, M.; Hong, J. Sequence-related amplified polymorphism (SRAP) marker as a new method for identification of endophytic fungi from Taxus. World J. Microbiol. Biotechnol. 2012, 28, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Polat, I.; Kacar, Y.A.; Yesiloglu, T.; Uzun, A.; Tuzcu, O.; Gulsen, O.; Incesu, M.; Kafa, G.; Turgutoglu, E.; Anil, S. Molecular characterization of sour orange (Citrus Aurantium) accessions and their relatives using SSR and SRAP markers. Genet. Mol. Res. 2012, 11, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jing, Z.B.; Cheng, J.M. Genetic diversity and population structure of stipa bungeana, an endemic species in Loess Plateau of China, revealed using combined ISSR and SRAP markers. Genet. Mol. Res. 2014, 13, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Jia, X.; Ni, H.; Sun, P.; Niu, S.; Chen, X. AFLP Analysis of genetic diversity of jatropha curcas grown in Hainan, China. Trees 2010, 24, 455–462. [Google Scholar] [CrossRef]
- Zhuang, F.M.; Ge, S. Data analysis in population genetics.I. analysis of RAPD data with AMOVA. Biodivers. Sci. 2002, 10, 438–444. [Google Scholar]
- Rohlf, F.J. NTSYSpc, Numerical Taxonomy and Multivariate Analysis System; Version 2.1; Exeter Publication Ltd Setauket: New York, NY, USA, 2000. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 4, 1567–1587. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Wang, S.; Liu, Y.; Ma, L.; Liu, H.; Tang, Y.; Wu, L.; Wang, Z.; Li, Y.; Wu, R.; Pang, X. Isolation and characterization of microsatellite markers and analysis of genetic diversity in Chinese jujube (Ziziphus Jujuba Mill.). PLoS ONE 2014, 9, e99842. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhang, X.; Cai, H.; Liu, W.; Peng, Y. Genetic diversity analysis and transferability of cereal EST-SSR markers to orchardgrass (Dactylis Glomerata L.). Biochem. Syst. Ecol. 2010, 38, 740–749. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- He, X.Y. Studied on Genetic of Zenia insignis Provenances and Their Resistance to Low Temperature Stress. Ph.D. Thesis, Central South University of Forestry and Technology, Changsha, China, July 2007. [Google Scholar]
- Huang, Y. Analysis of camellia meiocarpa genetic diversity based on srap markers. Sci. Silv. Sin. 2013, 49, 43–50. [Google Scholar]
- Hamrick, J.K; Godt, M.J.W.; Sherman, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, F.Q.; Pan, W. Genetic diversity and genetic structure in natural populations of castanopsis hysitrix from its main distribution in China. Sci. Silv. Sin. 2013, 49, 162–166. [Google Scholar]
- Li, C. Investigation on Adaptive Genetic Variation of Castanopsis Fargesii Using EST-SSR Molecular Markers. Master’s Thesis, South China Botanical Garden of Chinese Academy of Sciences, Guangzhou, China, May 2012. [Google Scholar]
- Liu, J.; Chen, Y.T.; Sun, Z.X. Spatial Genetic Structure of Toona ciliata var .pubescens Populations in Terms of Spatial Autocorrelation Analysis. Sci. Silv. Sin. 2008, 44, 45–52. [Google Scholar]
- Kong, C. Genentic Response of Toona ciliata var. Pubescens Families to Soil Nitrogen and Phosphorus Level. Master’s Thesis, China Academy of Forestry, Changsha, China, July 2012. [Google Scholar]
- Slatkin, M. Gene flow and the geographic structure of natural populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Greene, S.L.; Kisha, T.K.; Yu, L.X. Conserving plants in gene banks and nature: Investigating complementarity with Trifolium thompsonii Morton. PLoS ONE 2014, 9, e105145. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Zhan, X.; Que, Q.; Qu, W.; Liu, M.; Ouyang, K.; Li, J.; Deng, X.; Zhang, J.; Liao, B.; et al. Genetic Diversity and Population Structure of Toona Ciliata Roem. Based on Sequence-Related Amplified Polymorphism (SRAP) Markers. Forests 2015, 6, 1094-1106. https://0-doi-org.brum.beds.ac.uk/10.3390/f6041094
Li P, Zhan X, Que Q, Qu W, Liu M, Ouyang K, Li J, Deng X, Zhang J, Liao B, et al. Genetic Diversity and Population Structure of Toona Ciliata Roem. Based on Sequence-Related Amplified Polymorphism (SRAP) Markers. Forests. 2015; 6(4):1094-1106. https://0-doi-org.brum.beds.ac.uk/10.3390/f6041094
Chicago/Turabian StyleLi, Pei, Xin Zhan, Qingmin Que, Wenting Qu, Mingqian Liu, Kunxi Ouyang, Juncheng Li, Xiaomei Deng, Junjie Zhang, Boyong Liao, and et al. 2015. "Genetic Diversity and Population Structure of Toona Ciliata Roem. Based on Sequence-Related Amplified Polymorphism (SRAP) Markers" Forests 6, no. 4: 1094-1106. https://0-doi-org.brum.beds.ac.uk/10.3390/f6041094