Non-Saccharomyces in Winemaking: Source of Mannoproteins, Nitrogen, Enzymes, and Antimicrobial Compounds


Dirección de Investigación y Desarrollo, Universidad Privada del Norte (UPN), Trujillo 13001, Peru
Fermentation 2020, 6(3), 76; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030076
Submission received: 21 June 2020
/
Revised: 13 July 2020
/
Accepted: 21 July 2020
/
Published: 29 July 2020
(This article belongs to the Special Issue Enological Repercussions of Non-Saccharomyces Species 2.0)
Abstract
:Traditionally, non-Saccharomyces yeasts have been considered contaminants because of their high production of metabolites with negative connotations in wine. This aspect has been changing in recent years due to an increased interest in the use of these yeasts in the winemaking process. The majority of these yeasts have a low fermentation power, being used in mixed fermentations with Saccharomyces cerevisiae due to their ability to produce metabolites of enological interest, such as glycerol, fatty acids, organic acids, esters, higher alcohols, stable pigments, among others. Additionally, existing literature reports various compounds derived from the cellular structure of non-Saccharomyces yeasts with benefits in the winemaking process, such as polysaccharides, proteins, enzymes, peptides, amino acids, or antimicrobial compounds, some of which, besides contributing to improving the quality of the wine, can be used as a source of nitrogen for the fermentation yeasts. These compounds can be produced exogenously, and later incorporated into the winemaking process, or be uptake directly by S. cerevisiae from the fermentation medium after their release via lysis of non-Saccharomyces yeasts in sequential fermentations.
1. Introduction
The genus Saccharomyces has been the most industrially used in the production of wine. This aspect has been changing in recent years due to an increased interest in the use of non-Saccharomyces yeasts [1]. The high potential of these yeasts makes them a useful tool for improving oenological parameters such as nutrients content, stability, aromatic profile, or bioactive profile, in spite of their low fermentative power, and, in some cases, their high production of certain metabolites with negative connotations in the wine [2,3].
Currently, species such as Torulaspora delbrueckii, Lachancea thermotolerans, Metschnikowia pulcherrima, Schizosaccharomyces pombe, Pichia kluyveri, among others, are sold for wine production [4]. These yeasts are often isolated from the grape, grape-must, and wine, as well as from the soil, winery surfaces, harvesting machinery, and other objects associated with the wine production. In the case of grape-must/wine, the majority are isolated during the first stages of fermentative process given their low fermentation ability and low tolerance to ethanol, with respect to S. cerevisiae, so that its use would mainly be associated with the contribution of the specific enological metabolites such as enzymes, aromatic compounds, glycerol, organic acids, fatty acids, proteins, amino acids, polysaccharides, among others [3].
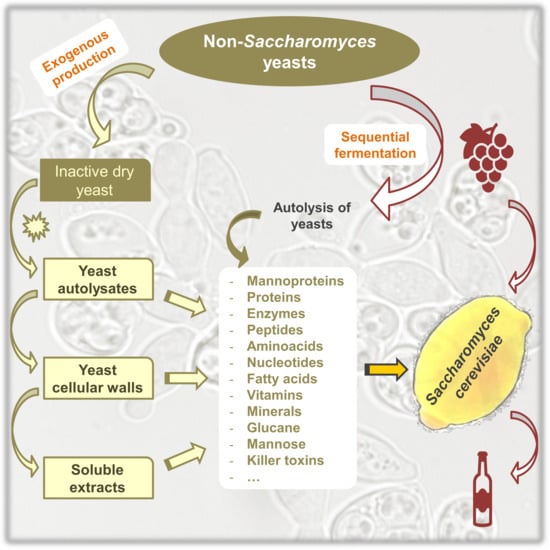
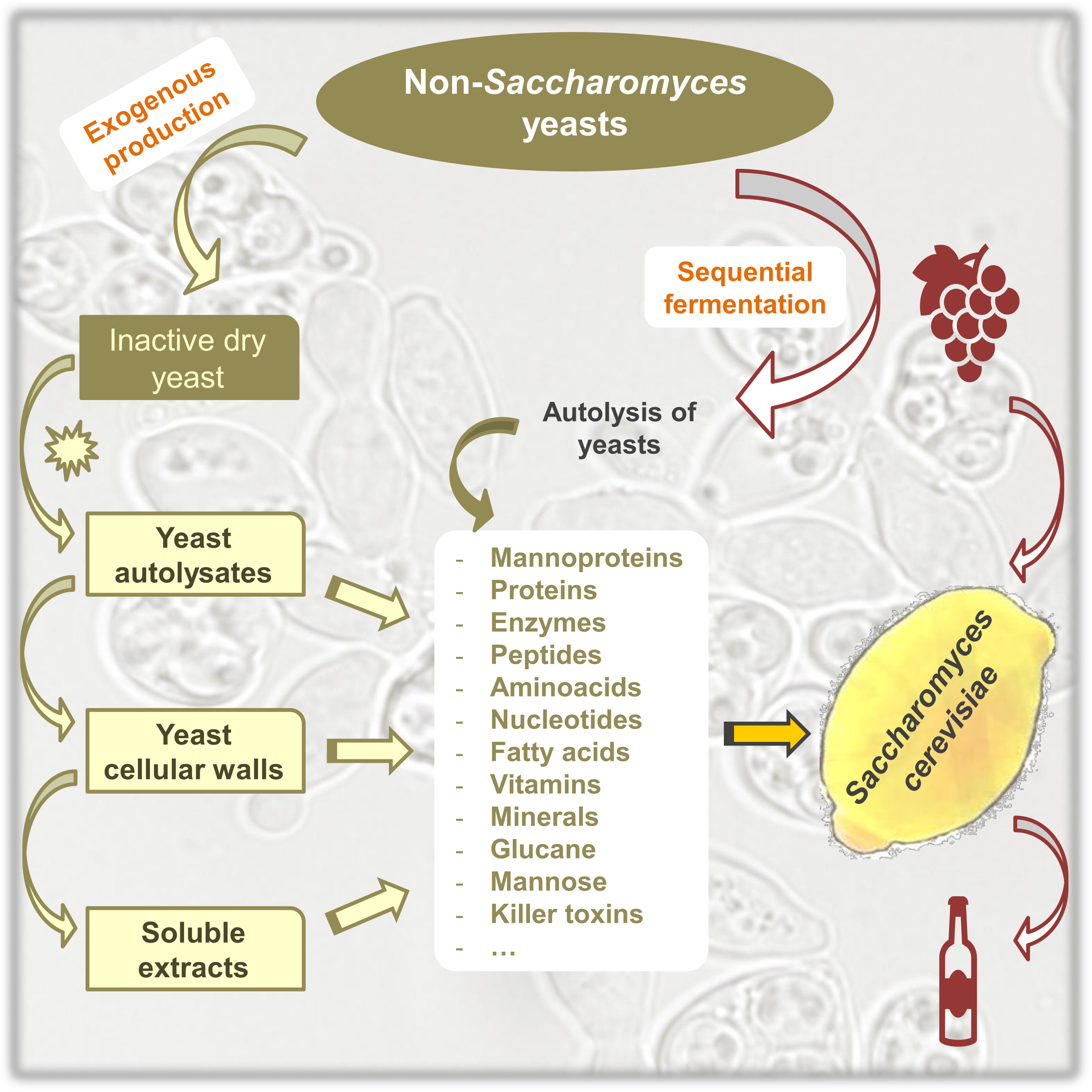
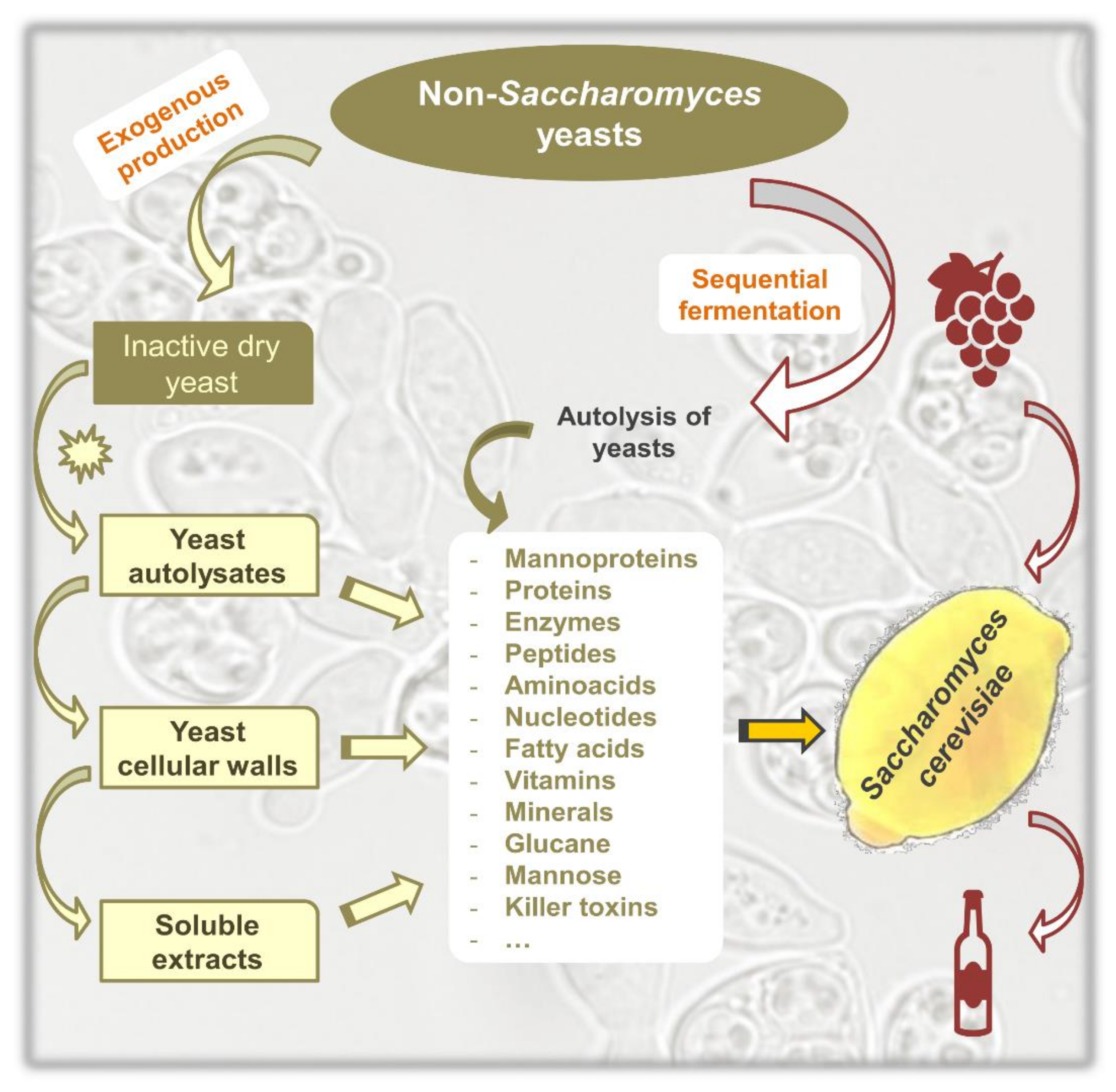
Figure 1 summarizes some of the potential applications of non-Saccharomyces yeasts, taking as reference the available commercial preparations for wine production. These preparations are sold in the form of (a) dry yeast, inactivated through thermal treatment and then dried; (b) cell autolysates, which include soluble and insoluble yeast components, partially degraded by the endogenous enzymes; (c) soluble cell extracts, derived from the cytoplasm; and (d) insoluble cell hulls, mainly containing cell walls [5]. These preparations may be the source of compounds of interest such as polysaccharides, proteins, enzymes, or other metabolites with the potential for use in wine production and can be produced exogenously [6].
In addition to these preparations, compounds of interest can be obtained directly from the fermentation process as a result of their release from the yeast during cell lysis, for example, in mixed cultures.
2. Non-Saccharomyces Yeasts as a Source of Polysaccharides
During aging-on-lees (AOL), a series of changes take place with a direct impact on the wine’s properties, due to the interaction between the wine’s components, the enzymatic activity, and the polysaccharides released by the lees [7]. The use of non-Saccharomyces yeasts for the exogenous production of polysaccharides [6], which can be added to wine, is an interesting alternative. The addition of yeast polysaccharides is a practice authorized by the International Code of Oenological Practice of the International Organization of Vine and Wine (OIV).
These polysaccharides, particularly in red wines, can improve the mouthfullness and body [8], sweetness and roundness [9], aromatic persistence [10], protein and tartaric stability [11,12], and interact with tannins and reduce astringency [13], as well as have an antioxidant effect (due to glutathione) that protects the aromatic compounds and anthocyanins [14], enabling its antioxidant and anti-inflammatory potential to be maintained [15]. They also interact with the tertiary aromatic compounds, reducing the perception of woody aromas in long-aged wines [16], encourage malolactic fermentation [17], and adsorb undesirable and dangerous compounds such as ochratoxin A (OTA) [18]. In sparkling wines, they improve the quality of the foam [19]. However, during AOL, a reduction of aromatic compounds such as terpenes, esters, and aldehydes [20], and, in some cases, anthocyanins in red wines [16,21], has also been reported.
The most important polysaccharides released during AOL are the mannoproteins (Table 1), which are fixed on a three-dimensional network of glucan and chitin in the cell wall [21,22]. The mannoproteins represent between 80% and 100% of the fraction of polysaccharides founded in the wine (molecular mass 100–2000 kDa: 90% mannose and 10% protein), while glucomannoproteins (molecular mass 20–90 kDa: 25% glucose, 25% mannose, and 50% protein) represent between 10% and 20% of the total [23]. Furthermore, α-galactomannose rather than mannose has been found as part of the structure of polysaccharides in S. pombe [22].
The enzymes involved in the cell autolysis and subsequent releasing of polysaccharides into the wine are endo β(1→3)-glucanases, endo β-(1→6)-glucanases, exo-(1→6)-α-D-mannose, exo-(1→2)-α-mannose, and α-D-mannosidase [23].
2.1. Non-Saccharomyces Species as a Source of Polysaccharides
Even though S. cerevisiae is the most commonly used yeast as a source of polysaccharides, the literature reports different non-Saccharomyces species with the potential to produce and release polysaccharides during AOL: S. pombe, Saccharomycodes ludwigii, Wickerhamomyces anomalus, Hanseniaspora vineae, L. thermotolerans, M. pulcherrima, Starmerella bacillaris, T. delbrueckii, Zygosaccharomyces rouxii, and Zygotorulaspora florentina (formerly Zygosaccharomyces florentinus), among others.
The genus Schizosaccharomyces has shown high rates of release of polysaccharides during alcoholic fermentation, of up to seven times higher than S. cerevisiae [24]. During AOL, S. pombe together with S’codes ludwigii are found among the species with high potentials for releasing polysaccharides [21]. S. pombe primarily releases galactomannoproteins (Table 1), at concentrations up to 10 times higher than some strains of Saccharomyces and Pichia [21]. This potential could mean an advantage with regard to accelerating aging processes.
S’codes ludwigii has also shown a high potential for releasing polysaccharides during fermentation and AOL [21,25,26]. Palomero et al. [21] obtained higher rates of polysaccharide release from S’codes. ludwigii (110.51 gm/L) and S. pombe (103.61 mg/L) with respect to S. cerevisiae (36.65 mg/L). Furthermore, the polysaccharides from these non-Saccharomyces yeasts show a greater molecular size and may potentially impact the wine’s palatability.
However, one aspect to take into account during AOL in red wines is color loss, due to the interaction between the lees and anthocyanins, mainly through adsorption of pigments by the lees [16] and through degradation due to anthocyanin-β-glucosidase activity [27]. The lees from M. pulcherrima, S’codes ludwigii, or S. pombe have shown a low adsorption of anthocyanins with respect to the lees of S. cerevisiae and other non-Saccharomyces yeasts such as T. delbrueckii or L. thermotolerans [21,28]. Furthermore, there may be less color loss in wines in the presence of higher pyranoanthocyanins content, which are more stable due to its chemical structure [29], with S. pombe being one of the yeasts with the greatest synthesis of these pigments [30].
Additionally, S. pombe has also shown the ability to decrease the biogenic amine content in sparkling wines, with better results than S. cerevisiae [30], which would be related to the adsorption of the amines on the lees during second fermentation and subsequent aging in the bottle [31]. However, considering the short duration of AOL studies, it is necessary to evaluate the evolution of amines over longer aging periods, such as those carried out in traditional sparkling wine production, given that other studies have reported an increase in the biogenic amines content in wines in contact with lees [32].
In terms of other non-Saccharomyces yeasts, Hanseniaspora vineae has shown a higher rate of cell lysis with respect to commercial S. cerevisiae strains [33]. Other yeasts that have shown a notable polysaccharide contribution are L. thermotolerans [34], Z. rouxii [26], and Z. florentina, particularly in mixed fermentations with S. cerevisiae [35].
For its part, the information regarding the use of other yeasts such as W. anomalus as a source of mannoproteins is scarce [26]. This yeast has a high potential for the production of polysaccharides and other metabolites of interest, given its ability to metabolize a large variety of nitrogen sources, including nitrate [36], which would allow production costs to be reduced.
2.2. Accelerated Release of Polysaccharides
One disadvantage of AOL is that it is a very slow process, which requires up to 9 months to obtain the desired effects in the treated wine. Among the strategies to improve AOL, existing literature reports (a) the selection of yeast species and strains with rapid autolysis [21,39], (b) acceleration of the cell autolysis through mixed cultures involving sensitive and killer yeasts [40], mixed cultures among different yeast species, which enable the regulation of cell death [41], the addition of β-glucanase [42], and the application of ultrasound [39,43].
Several studies have reported a higher rate of polysaccharide release with ultrasound (US) treatments [39,43,44]. Lees from S’codes ludwigii, S. pombe, M. pulcherrima, and S. cerevisiae, among others, were evaluated over a 7-week aging period in a hydroalcoholic medium, applying US for 10 minutes a day, with S’codes ludwigii lees showing the highest rate of polysaccharide release after the third week (around 460 mg/L) [39]. In the same study [39], a decrease in anthocyanins content was observed in the treated red wine, without affecting the pyranoanthocyanins content [21,29], particularly with S’codes ludwigii lees, which also allowed to reduce proanthocyanidins content, and consequently, astringency and bitterness.
More recently, Del Fresno et al. [43] evaluated the effect of AOL (lees from S. cerevisiae) in the presence of oak chips in red wines. The evaluation period was 135 days at 14°C, under dark conditions, agitating the samples once a week to simulate “bâtonnage”. The samples were treated with US twice a week for 5 minutes for the first 5 weeks. From this moment, the process was accelerated increasing to two US treatments a week (15 minutes per treatment). In a parallel experiment, a hydroalcoholic medium was used to adequately quantify the polysaccharides released from the lees. In general, the polysaccharide release increased after using US for 135 days, releasing around 11.8 mg/L, more than double that of the untreated samples (approx. 5.3 mg/L). Additionally, an increase in the protein content of the US-treated samples was observed after 120 days of AOL.
However, the same authors [43] reported the increase in dissolved oxygen in the treated red wines as a disadvantage, whose effect was evident in the lower anthocyanins content in the US-treated wines in the absence of lees. This finding reveals the protective effect of polysaccharides against oxidation.
3. Non-Saccharomyces Yeasts as a Source of Nitrogen for Saccharomyces cerevisiae
Yeast assimilable nitrogen (YAN) is nitrogen that yeasts can assimilate and metabolize, preferably from sources such as ammonium (NH4+), amino acids, and peptides of up to five amino acids [45]. Its content in the grape-must varies between 50 and 500 mg/L [46].
Among YAN’s functions are reproduction and cell growth, the protein for sugar transport synthesis, and enzyme synthesis, as well as the functions accomplished by the amino acids as the precursors of aromatic compounds, mainly higher alcohols, produced by deamination and decarboxylation [47].The most predominant amino acids as NH4+ transporters in grape-must are α-alanine, serine, arginine, proline, glutamic acid, and glutamine [48]. During fermentation, their use varies, with higher uptake of glutamic acid, glutamine, and arginine [4], whereas proline, a proteinogenic amino acid not metabolized by S. cerevisiae, is among the least used [4]. Additionally, the demand for arginine is increased by lactic acid bacteria during malolactic fermentation [48].
S. cerevisiae has shown a preference for NH4+ and amino acids, thanks to its nitrogen catabolite repression (NCR) mechanism [49], through which the genes involved in the transport and metabolism of NH4+ and glutamine are activated and, once they are depleted, the genes involved in the transport and metabolism of other sources such as arginine, glutamate, and alanine, among others, become activated.
Concentrations of YAN below 150 mg/L in the grape-must carry the risk of sluggish or stuck fermentation [4], as well as a low synthesis of some aromatic compounds such as of esters, volatile fatty acids, and higher alcohols [50]. This N deficiency is remedied in the winery through the addition of extra N sources such as diammonium phosphate (DAP). However, the excessive use of DAP can lower phenylpropanoid production (affecting the complexity of the wine) [51,52], increase the wine acidification, produce high levels of residual phosphate, stimulate the production of esters such as ethyl acetate, and increase the levels of hydrogen sulfide (H2S), especially when there is a deficit of other essential nutrients such as vitamins, minerals, and lipids; further, excessive levels of DAP can increase turbidity, promote microbial instability, and facilitate the production of unpleasant aromas and harmful compounds such as ethyl carbamate and biogenic amines [53,54,55,56].
3.1. Non-Saccharomyces Species as a Source of Nitrogen
As an alternative N source, yeast cell structures can be used in the form of hulls, hydrolyzed, or extracts, which can be produced exogenously [6], and then added to the fermentative medium. Additionally, this source of N can be obtained from the rest of the cells after the death and lysis of non-Saccharomyces yeasts used at the beginning of the sequential fermentations with S. cerevisiae.
Aureobasidium pullulans (an yeast-like fungus) is a potential source of essential amino acids [57], and it can growth in low-cost carbon sources like agricultural and food waste, due to its amylase [58], cellulase [59], lipase [60], xylanase [61], laccase [62], mannanase [63], and protease [64] activities. However, the use of A. pullulans for the production of protein on a large scale has still not been explored [65].
Similarly, the use of species such as S’codes ludwigii, S. pombe, Candida stellata, M. pulcherrima, W. anomalus, H. vineae, Z. rouxii, Zygosaccharomyces bailii, L. thermotolerans, and Z. florentina, among others, as potential N sources is limited, considering their high capacity for production and release of mannoproteins [21,25,26,33,34,35], which is up to 29% of protein (Table 1). From the little background information available, W. anomalus has been used as a source of single cell protein at industrial level, specifically to produce protein for fish farming [66]. This demonstrates the potential of W. anomalus as source of N, taking into account the ability of this yeast to use a wide range of N sources, including nitrate [36].
One aspect to consider is the depletion of nutrients during fermentation with different yeasts. Difficulties have been reported in the introduction of S. cerevisiae after Hanseniaspora/Kloeckera in sequential fermentations [67], an effect that is associated with the depletion of thiamine and calcium pantothenate [68,69], reducing the availability of these nutrients for S. cerevisiae (second inoculum).
Similarly, sequential fermentations can cause a depletion of YAN by first phase yeasts, especially those with high nutrient demands and a low ability to release nitrogenous compounds. This situation can be overcome with the use of yeasts such as M. pulcherrima, with high proteolytic activity and amino acid release as a source of N for S. cerevisiae (second inoculum) [70,71].
In a recent study, Prior et al. [4] carried out sequential fermentations of L. thermotolerans/S. cerevisiae and T. delbrueckii/S. cerevisiae to evaluate whether the rest of these non-Saccharomyces yeasts (first phase) can be used as a source of N for S. cerevisiae. After 48 hours of fermentation with non-Saccharomyces, the medium was filtered and then inoculated with S. cerevisiae. A reduction in sugar consumption by S. cerevisiae was observed in the filtered medium, in other words, fermentative yield decreased.
The results obtained in the aforementioned studies show that in sequential fermentations, non-Saccharomyces yeast rests (first phase) after death and lysis can release various cell components into the fermentative medium to be used as N sources for S. cerevisiae [4]. This strategy can be used more efficiently with non-Saccharomyces species with high ability to release mannoproteins (Table 1) and with high β-glucanase and protease activities.
3.2. Nitrogen Requirements for Sparkling Wines Production
The selection of yeasts with high rates of nitrogenous compounds release can ensure an adequate supply of N. Two of the non-Saccharomyces species which have shown the greatest ability to release amino acids during the second fermentation are S’codes ludwigii and S. pombe [30], increasing the amino acids content with respect to the base wine. The advantage of these yeasts over S. cerevisiae is related to their amino acid release mechanisms, with their different consumption rates and their cell composition [21,47].
One of the most important properties in sparkling wine is the ability to form foam or foamability. Mannoproteins improve foam formation and stability thanks to their hydrophobicity, high glycosylation, and high molecular mass, which enable them to surround and stabilize the gas bubbles in the foam [72]. The proteins also participate in this process which, together with the peptides and amino acids, are released mainly during aging in the bottle as a result of the enzymatic degradation of the cell walls and other cell structures [7,73], contributing to the complexity of sparkling wines.
The higher production of mannoproteins in sequential fermentations by S. bacillaris/S. cerevisiae [74], L. thermotolerans/S. cerevisiae, and T. delbrueckii/S. cerevisiae can also be utilized, which also contributes to reducing volatile acidity and increasing 2-phenylethanol [75,76], and improving foamability and foam stability (sequential T. delbrueckii/S. cerevisiae and M. pulcherrima/S. cerevisiae fermentations) [77,78].
Finally, during AOL (in the bottle), it has been observed that the content of free amino acids and peptides depends on the yeast species and strains [73], which have an influence on the flocculation ability to facilitate lees movement during riddling [79]. A potential field for future studies is the optimization of aging and disgorging operation in sparkling wines, for example, by inserting magnetic nanoparticles to accelerate sedimentation and lees removal [80].
4. Non-Saccharomyces Yeasts as a Source of Exogenous Enzymes
Another potential application of non-Saccharomyces yeasts is enzyme production. In general, enzymes of microbial origin are considered to have greater activity and stability than those of plant and animal origin [81]. Considering that the genus Saccharomyces is not characterized as being a good producer of exogenous enzymes [82], there is increasing interest in finding sources of enzymes of enological interest among non-Saccharomyces species, some of which are summarized in Table 2.
There is little information regarding the application of purified β-glucosidase, β-lyase, xylanase, cellulase, among others enzymes, produced by non-Saccharomyces yeasts for winemaking processes, leaving open the possibility of future research which focuses on improving the release of terpenes, thiols, norisoprenoids, and/or their precursors, with positive impacts on the aromatic profile, especially in white wines, considering that approximately 90% of these compounds are conjugated in the grape skin [99]. This is in addition to the selection of suitable strains, because high levels of β-glucosidase can increase the synthesis of undesirable volatile phenols [101], as well as hydrolyze the anthocyanins in red wines, exposing them to oxidation and/or transformation into colorless forms [27,112].
In contrast, one problem in white wines is the protein haze (wine turbidity), which is usually corrected by removing the proteins from the grape-must with bentonite, with the disadvantage of removing other compounds of enological interest, mainly aromatic compounds. The protease activity of non-Saccharomyces yeasts can be used (Table 2) and thus reduce the protein content in the grape-must and therefore prevent the wine haze. The proteases hydrolyze the proteins from the grape-must into smaller molecules like peptides and amino acids, with the consequent clarification and subsequent stability of the wine obtained, and with the additional advantage of providing YAN for the fermentative yeasts [87,89].
5. Non-Saccharomyces as Biocontrol Agents Against Contaminating Yeasts
5.1. Antimicrobial Peptides
Some peptides produced by yeasts have shown antimicrobial effects against several grape-must/wine contaminating yeasts. In general, these peptides show lengths of up to 100 amino acids, sorted into variable sequences [113]. Their mechanism of action would be related to changes in the integrity of the cell wall of the target yeasts [114]. Peptides with molecular mass below 10 kDa have shown greater antimicrobial effects [115], such as those produced by Candida intermedia, especially the LAMAP1790 strain, with an effect on several strains of Brettanomyces bruxellensis and without affecting the growth of S. cerevisiae [115].
S. cerevisiae CCMI885 is another yeast that produces antimicrobial peptides with molecular mass lower than 10 kDa and with an effect on B. bruxellensis, Hanseniaspora uvarum, Hanseniaspora guilliermondii, C. stellata, L. thermotolerans, Kluyveromyces marxianus, and T. delbrueckii [116]. However, as these peptides have not shown total inhibition over B. bruxellensis, their application may require the use of other usual winemaking treatments, such as the addition of SO2.
According to Peña et al. [117], to achieve the application of these peptides at an industrial level, it is necessary to understand their behavior in mediums with different pH values and sugar levels, as well as high alcohol levels and in the presence of other winemaking yeasts and/or bacteria. Additionally, the implementation of procedures that enable high production and, therefore, satisfy a potential industrial level demand is needed.
5.2. Killer Toxins
Some yeasts produce molecules called “killer toxins”, which are glycosylated proteins with effect against sensitive yeast strains [118]. For instance, the action of killer toxins CpKT1 and CpKT2 produced by Candida pyralidae (YWBT Y1140 strain) on the cell wall of B. bruxellensis have been reported [119]. These toxins have a molecular mass of over 50 kDa, are stable at acidic pH values (3.5–4.5), temperatures between 15 and 25 °C, at high alcoholic content, and at different sugar concentrations. In other words, they are stable under normal winemaking conditions.
In the same way, a killer toxin produced by T. delbrueckii NPCC 1033 (TdKT) has shown potential to control yeasts such as Brettanomyces bruxellensis, Pichia guilliermondii, Pichia mandshurica, and Pichia membranifaciens [120], being the mechanism of action, the attack at the cell wall level, related to their glucanase and chitinase enzymatic activities. The toxin has a molecular mass of over 30 kDa, its killer activity is stable at pH values of 4.2–4.8, and is inactivated at temperature above 40 °C, confirming their potential use as a biocontrol tool at oenological conditions.
Similarly, Kwkt toxins produced by Kluyveromyces wickerhamii [121] and PMKT2 produced by P. membranifaciens have shown effects on B. bruxellensis [122], although in the case of PMKT2, effects on S. cerevisiae have also been observed.
W. anomalus also produces killer toxins with effects on Dekkera/Brettanomyces, especially the Pikt toxin [121,123] produced by the D2 and DBVPG 3003 strains, whose fungicidal effect on wine can be sustained for 10 days [121]. The mechanism of action would be the attack at the cell wall level, specifically of β-1,6 glucans. [124], in a similar manner to the W. anomalus Cf20 toxin (KTCf20) that binds to the β-1,3 and β-1,6 glucans. β-1,3-glucanase activity has been previously reported in toxins secreted by W. anomalus [100]. Additionally, several toxins secreted by this yeast has shown stability and high activity at pH values of 3.0-5.0 and temperatures up to 30 °C, i.e., these toxins are compatible with the winemaking conditions [100,125].
Other studies have reported the antimicrobial effect of W. anomalus on species such as Pichia guilliermondii or P. membranifaciens [125,126] during the first stages of fermentation, and even on S. cerevisiae [125,127]. This indicates the need for more studies to evaluate the compatibility of the W. anomalus strains that produce these toxins with other yeasts to avoid technological problems such as sluggish and stuck fermentations, as well as take advantage of the potential of the different non-Saccharomyces species involved in the fermentation process, to obtain wines with greater complexity and stability.
Although it is not a non-Saccharomyces yeast, Saccharomyces eubayanus has also shown the ability to produce the killer toxin SeKT, with effect on spoilage yeasts such as B. bruxellensis, P. membranifaciens, Meyerozyma guilliermondii and P. manshurica [128]. The mechanism of action comprises cell wall disruption through β-glucanase and chitinase activities. The toxin have a molecular mass of around 70 kDa, and has shown stability at high glucose and ethanol concentrations (300 g/L and 16% v/v, respectively), at SO2 concentrations of up to 100 mg/L, and at temperatures and pH values less than 26 °C and 5.0, respectively.
The results obtained in the aforementioned studies show the potential use of the killer toxins as a biocontrol tool at oenological conditions, especially against the spoilage yeast B. bruxellensis, for example, during wine aging and storing.
5.3. Other Molecules as Biocontrol Agents
One feature of M. pulcherrima is its potential effect against different contaminating yeast species. In the context of wine production, this activity would be mainly related to the production of the pigment pulcherrimin, from the chelation of Fe in the fermentative medium [129], reducing the availability of this mineral, with harmful effects on Brettanomyces/Dekkera, Pichia, Hanseniaspora, S’codes ludwigii, and Candida [130].
However, the most significant advantage of this antimicrobial mechanism is the absence of harmful effects on S. cerevisiae [130]. In other words, it may be compatible with the main yeast used for wine production, for example, in mixed fermentations, with the consequent reduction of the SO2 dose, usually used as an antimicrobial agent [2].
6. Future Perspectives
One of the greatest challenges related to the industrial use of non-Saccharomyces yeasts for AOL is achieving acceptance from producers, especially in regions with a deep-rooted winemaking tradition and taking into account the unfavorable background of these yeasts, as well as the economic impact of prolonged storage of the wines in AOL and the possible risks of contamination. The optimization of aging conditions is one aspect that requires special attention, especially those conditions which enable the acceleration of the release of polysaccharides from the lees.
Another aspect that requires more research is the use of exogenously produced lees added to the wine [6], especially for the aging of red wines, which involves the identification of species, and especially strains with high rates of polysaccharide release which, in turn, present low anthocyanin adsorption [28] and low expression of anthocyanase activity (anthocyanin-β-glucosidase) that causes hydrolysis of anthocyanins [27]. Additionally, this requires strategies, which can be simultaneously implemented with the selection of yeasts with high capacity to produce pyranoanthocyanins, more stable against degradation [27], thereby minimizing color loss. One of the yeasts that has shown these characteristics is S. pombe.
There is also a need for additional studies on fermentation with natural grape-musts to verify the properties of mannoproteins released by non-Saccharomyces yeasts in synthetic mediums, given that in these last mediums, there is no interaction between the mannoproteins and the components which are naturally present in the grape-must, and to take advantage of the potential of these yeasts for the production of new products with industrial applicability [74].
Similarly, the effect of the addition of inactive yeasts, hulls, or lees as sources of N on parameters related to protein haze and turbidity formation is still not clear. One possible alternative is the addition of protease-hydrolyzed hulls/lees, which are also produced by non-Saccharomyces yeasts. M. pulcherrima has shown high protease activity and, therefore, the release of amino acids as a source of N for S. cerevisiae, especially in mixed cultures. This activity has also been shown by W. anomalus, with the advantage of using a wide range of N sources, including nitrate [36], and by A. pullulans, with the ability to use low-cost carbon sources such as agricultural and food waste, and whose protein production on a large scale has still not been studied [65].
It has been reported that the species and strain of the yeast influence the free amino acids and peptide content during AOL [57,58], thus reducing the content of amino acids in aging periods of over 9 months [73]. Therefore, it is necessary to conduct studies, which simulate normal aging conditions (even up to 10 years in sparkling wines) by evaluating the impact of the proteins, mannose, glucose, and galactose present in the lees on the quality of treated wine, which, until now, has only been studied in model mediums [131]. The effect of proteins from non-Saccharomyces yeasts on the quality of sparkling wines, their effect on lees movement during riddling, as well as the effect on the sensorial profile must also be studied.
Finally, most of the positive contributions of non-Saccharomyces yeasts with regard to S. cerevisiae are related to a higher presence of active enzymes, which depends, in part, on the carbon and nitrogen sources present in the fermentative medium. Small changes in the composition of the medium can affect the nature, quantity, and diversity of the secreted enzymes [132]. Therefore, proper maintenance of N levels in the fermentative medium is of vital importance, which can be achieved by identifying yeast species and strains with high release rates of nitrogenous compounds, mainly amino acids.
Funding
Project UPN-20201003 “Non-Saccharomyces yeasts: potential applications at industrial level” (Universidad Privada del Norte, Peru).
Conflicts of Interest
The author declares no conflict of interest.
References
- Morata, A. Enological repercussions of non-Saccharomyces species in wine biotechnology. Fermentation 2019, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Vejarano, R. Saccharomycodes ludwigii, control and potential uses in winemaking processes. Fermentation 2018, 4, 71. [Google Scholar] [CrossRef] [Green Version]
- Ivit, N.N.; Kemp, B. The impact of non-Saccharomyces yeast on traditional method sparkling wine. Fermentation 2018, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Prior, K.J.; Bauer, F.F.; Divol, B. The utilisation of nitrogenous compounds by commercial non-Saccharomyces yeasts associated with wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Del Barrio-Galán, R.; Ortega-Heras, M.; Sánchez-Iglesias, M.; Pérez-Magariño, S. Interactions of phenolic and volatile compounds with yeast lees, commercial yeast derivatives and non toasted chips in model solutions and young red wines. Eur. Food Res. Technol. 2012, 234, 231–244. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Morata, A. Nuevo Método de Crianza Sobre Lías. Oficina Española de Patentes y Marcas ES2311372B2, 7 October 2009. [Google Scholar]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine—A review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Palacios, A.; Ayestarán, B. Maceration enzymes and mannoproteins: A possible strategy to increase colloidal stability and color extraction in red wines. J. Agric. Food Chem. 2007, 55, 4854–4862. [Google Scholar] [CrossRef]
- Chalier, P.; Angot, B.; Delteil, D.; Doco, T.; Gunata, Z. Interactions between aroma compounds and whole mannoprotein isolated from Saccharomyces cerevisiae strains. Food Chem. 2007, 100, 22–30. [Google Scholar] [CrossRef]
- Gonzalez-Ramos, D.; Cebollero, E.; Gonzalez, R. A recombinant Saccharomyces cerevisiae strain overproducing mannoproteins stabilizes wine against protein haze. Appl. Environ. Microbiol. 2008, 74, 5533–5540. [Google Scholar] [CrossRef] [Green Version]
- Lubbers, S.; Leger, B.; Charpentier, C.; Feuillat, M. Effet colloide-protecteur d’extraits de parois de levures sur la stabilité tartrique d’une solution hydro-alcoolique modèle. OENO One 1993, 27, 13. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.; Ricardo-Da-Silva, J.M.; Lucas, C.; Laureano, O. Effect of commercial mannoproteins on wine colour and tannins stability. Food Chem. 2012, 131, 907–914. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Suárez-Lepe, J.A. Influence of yeasts in wine colour. In Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2016; pp. 285–305. [Google Scholar]
- Vejarano, R.; Gil-Calderón, A.; Díaz-Silva, V.; León-Vargas, J. Improvement of the bioactive profile in wines and its incidence on human health: Technological strategies. In Advances in Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Loira, I.; Vejarano, R.; Morata, A.; Ricardo-Da-Silva, J.M.; Laureano, O.; González, M.C.; Suárez-Lepe, J.A. Effect of Saccharomyces strains on the quality of red wines aged on lees. Food Chem. 2013, 139, 1044–1051. [Google Scholar] [CrossRef]
- Rosi, I.; Gheri, A.; Domizio, P.; Fia, G. Production de macromolecules parietals de Saccharomyces cerevisiae au cours de la fermentation et leur influence sur la fermentation malolactique. Rev. des Œnologues 2000, 94, 18–20. [Google Scholar]
- Moruno, E.G.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Moreno-Arribas, V.; Pueyo, E.; Nieto, F.J.; Martín-Álvarez, P.J.; Polo, M.C. Influence of the polysaccharides and the nitrogen compounds on foaming properties of sparkling wines. Food Chem. 2000, 70, 309–317. [Google Scholar] [CrossRef]
- Gallardo-Chacón, J.J.; Vichi, S.; López-Tamames, E.; Buxaderas, S. Changes in the sorption of diverse volatiles by Saccharomyces cerevisiae lees during sparkling wine aging. J. Agric. Food Chem. 2010, 58, 12426–12430. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar] [CrossRef]
- Kopecká, M.; Fleet, G.H.; Phaff, H.J. Ultrastructure of the cell wall of Schizosaccharomyces pombe following treatment with various glucanases. J. Struct. Biol. 1995, 114, 140–152. [Google Scholar] [CrossRef]
- Hidalgo, J. Tratado de Enología, Tomo I, 2nd ed.; Mundiprensa: Madrid, Spain, 2011; ISBN 84-8476-119-3. [Google Scholar]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Giovani, G.; Rosi, I.; Bertuccioli, M. Quantification and characterization of cell wall polysaccharides released by non-Saccharomyces yeast strains during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Wightman, J.D.; Wrolstad, R.E. β-glucosidase activity in juice-processing enzymes based on anthocyanin analysis. J. Food Sci. 1996, 61, 544–548. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Gómez-Cordovés, M.C.; Calderón, F.; Suárez, J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006, 106, 123–129. [Google Scholar] [CrossRef]
- Ivit, N.N.; Loira, I.; Morata, A.; Benito, S.; Palomero, F.; Suárez-Lepe, J.A. Making natural sparkling wines with non-Saccharomyces yeasts. Eur. Food Res. Technol. 2018, 244, 925–935. [Google Scholar] [CrossRef]
- Granchi, L.; Romano, P.; Mangani, S.; Guerrini, S.; Vincenzini, M. Production of biogenic amines by wine microorganisms. Bull. OIV 2005, 78, 595–609. [Google Scholar]
- Marcobal, A.; Martín-Álvarez, P.J.; Polo, M.C.; Muñoz, R.; Moreno-Arribas, M.V. Formation of biogenic amines throughout the industrial manufacture of red wine. J. Food Prot. 2006, 69, 397–404. [Google Scholar] [CrossRef]
- Martin, V. Hanseniaspora Vineae: Caracterización y su Uso en la Vinificación; Universidad de la Republica: Montevideo, Uruguay, 2016. [Google Scholar]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef]
- Lencioni, L.; Romani, C.; Gobbi, M.; Comitini, F.; Ciani, M.; Domizio, P. Controlled mixed fermentation at winery scale using Zygotorulaspora florentina and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2016, 234, 36–44. [Google Scholar] [CrossRef]
- Zha, Y.; Hossain, A.H.; Tobola, F.; Sedee, N.; Havekes, M.; Punt, P.J. Pichia anomala 29X: A resistant strain for lignocellulosic biomass hydrolysate fermentation. FEMS Yeast Res. 2013, 13, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugelsang, K.C.; Edwards, C.G. Wine Microbiology: Practical Applications and Procedures; Springer Science & Business Media: Berlin, Germany, 2007; ISBN 038733341X. [Google Scholar]
- Loira, I.; Morata, A.; Palomero, F.; González, C.; Suárez-Lepe, J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation 2018, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, P.; Loira, I.; Morata, A.; Tesfaye, W.; González, M.C.; Suárez-Lepe, J.A. Use of non-Saccharomyces yeast strains coupled with ultrasound treatment as a novel technique to accelerate ageing on lees of red wines and its repercussion in sensorial parameters. LWT 2015, 64, 1262. [Google Scholar] [CrossRef]
- Todd, B.E.N.; Fleet, G.H.; Henschke, P.A. Promotion of autolysis through the interaction of killer and sensitive yeasts: Potential application in sparkling wine production. Am. J. Enol. Vitic. 2000, 51, 65–72. [Google Scholar]
- Englezos, V.; Rantsiou, K.; Giacosa, S.; Río Segade, S.; Rolle, L.; Cocolin, L. Cell-to-cell contact mechanism modulates Starmerella bacillaris death in mixed culture fermentations with Saccharomyces cerevisiae. Int. J. Food Microbiol. 2019, 289, 106–114. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; González, M.C.; Suárez-Lepe, J.A. Conventional and enzyme-assisted autolysis during ageing over lees in red wines: Influence on the release of polysaccharides from yeast cell walls and on wine monomeric anthocyanin content. Food Chem. 2007, 105, 838–846. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Loira, I.; Morata, A.; González, C.; Suárez-Lepe, J.A.; Cuerda, R. Application of ultrasound to improve lees ageing processes in red wines. Food Chem. 2018, 261, 157–163. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Morata, A.; Escott, C.; Loira, I.; Cuerda, R.; Suárez-Lepe, J.A. Sonication of yeast biomasses to improve the ageing on lees technique in red wines. Molecules 2019, 24, 635. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.S. Fermentation. In Wine Science: Principles and Applications; Jackson, R.S., Ed.; Academic Press/Elsevier: Hoboken, NJ, USA, 2014; ISBN 9780123814692. [Google Scholar]
- Bely, M.; Rinaldi, A.; Dubourdieu, D. Influence of assimilable nitrogen on volatile acidity production by Saccharomyces cerevisiae during high sugar fermentation. J. Biosci. Bioeng. 2003, 96, 507–512. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [Green Version]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Nitrogen compounds. In Handbook of Enology: The Chemistry of Wine Stabilization and Treatments; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2006; ISBN 978-0-470-01037-2. [Google Scholar]
- Cooper, T.G. Transmitting the signal of excess nitrogen in Saccharomyces cerevisiae from the Tor proteins to the GATA factors: Connecting the dots. FEMS Microbiol. Rev. 2002, 26, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Koker, S. Nitrogen Utilisation of Selected Non-Saccharomyces Yeasts and the Impact on Volatile Compound Production; Stellenbosch University: Stellenbosch, South Africa, 2015. [Google Scholar]
- Martin, V.; Giorello, F.; Fariña, L.; Minteguiaga, M.; Salzman, V.; Boido, E.; Aguilar, P.S.; Gaggero, C.; Dellacassa, E.; Mas, A.; et al. De novo synthesis of benzenoid compounds by the yeast Hanseniaspora vineae increases the flavor diversity of wines. J. Agric. Food Chem. 2016, 64, 4574–4583. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.; Boido, E.; Giorello, F.; Mas, A.; Dellacassa, E.; Carrau, F. Effect of yeast assimilable nitrogen on the synthesis of phenolic aroma compounds by Hanseniaspora vineae strains. Yeast 2016, 33, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burin, V.M.; Gomes, T.M.; Caliari, V.; Rosier, J.P.; Bordignon Luiz, M.T. Establishment of influence the nitrogen content in musts and volatile profile of white wines associated to chemometric tools. Microchem. J. 2015, 122, 20–28. [Google Scholar] [CrossRef]
- Mendes-Ferreira, A.; Barbosa, C.; Lage, P.; Mendes-Faia, A. The impact of nitrogen on yeast fermentation and wine quality. Cienc. Tec. Vitivinic. 2011, 26, 17–32. [Google Scholar]
- Sturgeon, J.Q.; Bohlscheid, J.C.; Edwards, C.G. The effect of nitrogen source on yeast metabolism and H2S formation. J. Wine Res. 2013, 24, 182–194. [Google Scholar] [CrossRef]
- Ribereau-Gayon, P.; Dubourdieu, D.; Doneche, B.; Lonvaud, A. Handbook of Enology: The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2006; Volume 1, ISBN 978-0-470-01034-1. [Google Scholar]
- Chi, Z.; Yan, K.; Gao, L.; Li, J.; Wang, X.; Wang, L. Diversity of marine yeasts with high protein content and evaluation of their nutritive compositions. J. Mar. Biol. Assoc. United Kingdom 2008, 88, 1347–1352. [Google Scholar] [CrossRef]
- Manitchotpisit, P.; Skory, C.D.; Leathers, T.D.; Lotrakul, P.; Eveleigh, D.E.; Prasongsuk, S.; Punnapayak, H. α-Amylase activity during pullulan production and α-amylase gene analyses of Aureobasidium pullulans. J. Ind. Microbiol. Biotechnol. 2011, 38, 1211–1218. [Google Scholar] [CrossRef]
- Leite, R.S.R.; Bocchini, D.A.; Martins, E.S.; Silva, D.; Gomes, E.; Da Silva, R. Production of cellulolytic and hemicellulolytic enzymes from Aureobasidium pullulans on solid state fermentation. Appl. Biochem. Biotechnol. 2007, 137–140, 281–288. [Google Scholar]
- Leathers, T.D.; Rich, J.O.; Anderson, A.M.; Manitchotpisit, P. Lipase production by diverse phylogenetic clades of Aureobasidium pullulans. Biotechnol. Lett. 2013, 35, 1701–1706. [Google Scholar] [CrossRef]
- Manitchotpisit, P.; Leathers, T.D.; Peterson, S.W.; Kurtzman, C.; Li, X.L.; Eveleigh, D.E.; Lotrakul, P.; Prasongsuk, S.; Dunlap, C.A.; Vermillion, K.E.; et al. Multilocus phylogenetic analyses, pullulan production and xylanase activity of tropical isolates of Aureobasidium pullulans. Mycol. Res. 2009, 113, 1107–1120. [Google Scholar] [CrossRef]
- Rich, J.O.; Leathers, T.D.; Anderson, A.M.; Bischoff, K.M.; Manitchotpisit, P. Laccases from Aureobasidium pullulans. Enzyme Microb. Technol. 2013, 53, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Wang, F.; Chi, Z.; Yue, L.; Liu, G.; Zhang, T. Bioproducts from Aureobasidium pullulans, a biotechnologically important yeast. Appl. Microbiol. Biotechnol. 2009, 82, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Chi, Z.; Ma, C.; Madzak, C. Cloning, characterization, and expression of the gene encoding alkaline protease in the marine yeast Aureobasidium pullulans 10. Mar. Biotechnol. 2008, 10, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Bozoudi, D.; Tsaltas, D. The multiple and versatile roles of Aureobasidium pullulans in the vitivinicultural sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Huyben, D.; Nyman, A.; Vidaković, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of non-Saccharomyces yeasts affects nutrient availability for Saccharomyces cerevisiae during wine fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef]
- Bataillon, M.; Rico, A.; Sablayrolles, J.M.; Salmon, J.M.; Barre, P. Early thiamin assimilation by yeasts under enological conditions: Impact on alcoholic fermentation kinetics. J. Ferment. Bioeng. 1996, 82, 145–150. [Google Scholar] [CrossRef]
- Wang, X.D.; Bohlscheid, J.C.; Edwards, C.G. Fermentative activity and production of volatile compounds by Saccharomyces grown in synthetic grape juice media deficient in assimilable nitrogen and/or pantothenic acid. J. Appl. Microbiol. 2003, 94, 349–359. [Google Scholar] [CrossRef]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and ecological diversity of food and beverage yeasts. In Yeasts in Food and Beverages; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from Douro Wine Region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Vincenzi, S.; Crapisi, A.; Curioni, A. Foamability of Prosecco wine: Cooperative effects of high molecular weight glycocompounds and wine PR-proteins. Food Hydrocoll. 2014, 34, 202–207. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, A.J.; Carrascosa, A.V.; Martín-Álvarez, P.J.; Moreno-Arribas, V.; Polo, M.C. Influence of the yeast strain on the changes of the amino acids, peptides and proteins during sparkling wine production by the traditional method. J. Ind. Microbiol. Biotechnol. 2002, 29, 314–322. [Google Scholar] [CrossRef]
- Lemos Junior, W.J.F.; Nadai, C.; Rolle, L.; da Silva Gulao, E.; Miguez da Rocha Leão, M.H.; Giacomini, A.; Corich, V.; Vincenzi, S. Influence of the mannoproteins of different strains of Starmenella bacillaris used in single and sequential fermentations on foamability, tartaric and protein stabilities of wines. OENO One 2020, 54, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Del Fresno, J.M.; Morata, A.; Loira, I.; Bañuelos, M.A.; Escott, C.; Benito, S.; González Chamorro, C.; Suárez-Lepe, J.A. Use of non-Saccharomyces in single-culture, mixed and sequential fermentation to improve red wine quality. Eur. Food Res. Technol. 2017, 243, 2175–2185. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Medina-Trujillo, L.; González-Royo, E.; Sieczkowski, N.; Heras, J.; Canals, J.M.; Zamora, F. Effect of sequential inoculation (Torulaspora delbrueckii/Saccharomyces cerevisiae) in the first fermentation on the foaming properties of sparkling wine. Eur. Food Res. Technol. 2017, 243, 681–688. [Google Scholar] [CrossRef]
- Perpetuini, G.; Di Gianvito, P.; Arfelli, G.; Schirone, M.; Corsetti, A.; Tofalo, R.; Suzzi, G. Biodiversity of autolytic ability in flocculent Saccharomyces cerevisiae strains suitable for traditional sparkling wine fermentation. Yeast 2016, 33, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Loira, I.; Morata, A.; Escott, C.; Del Fresno, J.M.; Tesfaye, W.; Palomero, F.; Suárez-Lepe, J.A. Applications of nanotechnology in the winemaking process. Eur. Food Res. Technol. 2020, 246, 1533–1541. [Google Scholar] [CrossRef]
- Anbu, P.; Gopinath, S.C.B.; Chaulagain, B.P.; Lakshmipriya, T. Microbial enzymes and their applications in industries and medicine 2016. Biomed Res. Int. 2017, 2017, 2195808. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Advances in the study of Candida stellata. Fermentation 2018, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Díaz, M.; Arroyo, T. Biotechnological potential of non-Saccharomyces yeasts isolated during spontaneous fermentations of Malvar (Vitis vinifera cv. L.). Eur. Food Res. Technol. 2013, 236, 193–207. [Google Scholar] [CrossRef]
- Swangkeaw, J.; Vichitphan, S.; Butzke, C.E.; Vichitphan, K. Characterization of β-glucosidases from Hanseniaspora sp. and Pichia anomala with potentially aroma-enhancing capabilities in juice and wine. World J. Microbiol. Biotechnol. 2011, 27, 423–430. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of non-Saccharomyces yeasts with enzymatic activities of interest in wine-making. Int. J. Food Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Hock, R.; Benda, I.; Schreier, P. Formation of terpenes by yeasts during alcoholic fermentation. Z. Lebensm. Unters. Forsch. 1984, 179, 450–452. [Google Scholar] [CrossRef]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Jin, G.J.; Xu, Y.H.; Tao, Y.S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Bovo, B.; Carlot, M.; Lombardi, A.; Lomolino, G.; Lante, A.; Giacomini, A.; Corich, V. Exploring the use of Saccharomyces cerevisiae commercial strain and Saccharomycodes ludwigii natural isolate for grape marc fermentation to improve sensory properties of spirits. Food Microbiol. 2014, 41, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Morata de Ambrosini, V.I. Highly cold-active pectinases under wine-like conditions from non-Saccharomyces yeasts for enzymatic production during winemaking. Lett. Appl. Microbiol. 2015, 60, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Onetto, C.A.; Borneman, A.R.; Schmidt, S.A. Investigating the effects of Aureobasidium pullulans on grape juice composition and fermentation. Food Microbiol. 2020, 90, 103451. [Google Scholar] [CrossRef]
- Schlander, M.; Distler, U.; Tenzer, S.; Thines, E.; Claus, H. Purification and properties of yeast proteases secreted by Wickerhamomyces anomalus 227 and Metschnikovia pulcherrima 446 during growth in a white grape juice. Fermentation 2017, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Theron, L.W.; Bely, M.; Divol, B. Monitoring the impact of an aspartic protease (MpAPr1) on grape proteins and wine properties. Appl. Microbiol. Biotechnol. 2018, 102, 5173–5183. [Google Scholar] [CrossRef] [PubMed]
- Reid, V.J.; Theron, L.W.; Toit, M.D.; Divol, B. Identification and partial characterization of extracellular aspartic protease genes from Metschnikowia pulcherrima IWBT Y1123 and Candida apicola IWBT Y1384. Appl. Environ. Microbiol. 2012, 78, 6838–6849. [Google Scholar] [CrossRef] [Green Version]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; Van Rensburg, P. Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Schwentke, J.; Sabel, A.; Petri, A.; König, H.; Claus, H. The yeast Wickerhamomyces anomalus AS1 secretes a multifunctional exo-β-1,3-glucanase with implications for winemaking. Yeast 2014, 31, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Claus, H.; Mojsov, K. Enzymes for wine fermentation: Current and perspective applications. Fermentation 2018, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Izgü, F.; Altinbay, D.; Acun, T. Killer toxin of Pichia anomala NCYC 432; purification, characterization and its exo-β-1,3-glucanase activity. Enzyme Microb. Technol. 2006, 39, 669–676. [Google Scholar] [CrossRef]
- Morata, A.; Vejarano, R.; Ridolfi, G.; Benito, S.; Palomero, F.; Uthurry, C.; Tesfaye, W.; González, C.; Suárez-Lepe, J.A. Reduction of 4-ethylphenol production in red wines using HCDC+ yeasts and cinnamyl esterases. Enzyme Microb. Technol. 2013, 52, 99–104. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Effects of non-Saccharomyces yeasts on color, anthocyanin, and anthocyanin-derived pigments of Tannat grapes during fermentation. Am. J. Enol. Vitic. 2018, 69, 148–156. [Google Scholar] [CrossRef]
- Benito, S.; Morata, A.; Palomero, F.; González, M.C.; Suárez-Lepe, J.A. Formation of vinylphenolic pyranoanthocyanins by Saccharomyces cerevisiae and Pichia guillermondii in red wines produced following different fermentation strategies. Food Chem. 2011, 124, 15–23. [Google Scholar] [CrossRef]
- Lubbers, M.W.; Rodriguez, S.B.; Honey, N.K.; Thornton, R.J. Purification and characterization of urease from Schizosaccharomyces pombe. Can. J. Microbiol. 1996, 42, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Uthurry, C.A.; Suárez Lepe, J.A.; Lombardero, J.; García Del Hierro, J.R. Ethyl carbamate production by selected yeasts and lactic acid bacteria in red wine. Food Chem. 2006, 94, 262–270. [Google Scholar] [CrossRef]
- De Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans can lower ochratoxin a contamination in wine grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Apaliya, M.T.; Mahunu, G.K.; Chen, L.; Li, W. Control of ochratoxin A-producing fungi in grape berry by microbial antagonists: A review. Trends Food Sci. Technol. 2016, 51, 88–97. [Google Scholar] [CrossRef]
- Belda, I.; Conchillo, L.B.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Selection and use of pectinolytic yeasts for improving clarification and phenolic extraction in winemaking. Int. J. Food Microbiol. 2016, 223, 1–8. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased β-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the enzymatic basis of wine “Flavorome”: A phylo-functional study of wine related yeast species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.M.H.; Boss, P.K.; Walker, M.E.; Sumby, K.M.; Grbin, P.R.; Jiranek, V. Evaluation of indigenous non-Saccharomyces yeasts isolated from a South Australian vineyard for their potential as wine starter cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef]
- Manzanares, P.; Rojas, V.; Genovés, S.; Vallés, S. A preliminary search for anthocyanin-β-D-glucosidase activity in non-Saccharomyces wine yeasts. Int. J. Food Sci. Technol. 2000, 35, 95–103. [Google Scholar] [CrossRef]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Branco, P.; Viana, T.; Albergaria, H.; Arneborg, N. Antimicrobial peptides (AMPs) produced by Saccharomyces cerevisiae induce alterations in the intracellular pH, membrane permeability and culturability of Hanseniaspora guilliermondii cells. Int. J. Food Microbiol. 2015, 205, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña, R.; Ganga, M.A. Novel antimicrobial peptides produced by Candida intermedia LAMAP1790 active against the wine-spoilage yeast Brettanomyces bruxellensis. Antonie van Leeuwenhoek 2019, 112, 297–304. [Google Scholar]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef]
- Peña, R.; Chávez, R.; Rodríguez, A.; Ganga, M.A. A control alternative for the hidden enemy in the wine cellar. Fermentation 2019, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.J.; Breinig, F. The viral killer system in yeast: From molecular biology to application. FEMS Microbiol. Rev. 2002, 26, 257–276. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int. J. Food Microbiol. 2014, 188, 83–91. [Google Scholar] [CrossRef]
- Villalba, M.L.; Susana Sáez, J.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef]
- Comitini, F.; Ingeniis De, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.; San Mauro, M.; Bravo, E.; Marquina, D. PMKT2, a new killer toxin from Pichia membranifaciens, and its promising biotechnological properties for control of the spoilage yeast Brettanomyces bruxellensis. Microbiology 2009, 155, 624–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oro, L.; Ciani, M.; Bizzaro, D.; Comitini, F. Evaluation of damage induced by Kwkt and Pikt zymocins against Brettanomyces/Dekkera spoilage yeast, as compared to sulphur dioxide. J. Appl. Microbiol. 2016, 121, 207–214. [Google Scholar] [CrossRef] [PubMed]
- De Ingeniis, J.; Raffaelli, N.; Ciani, M.; Mannazzu, I. Pichia anomala DBVPG 3003 secretes a ubiquitin-like protein that has antimicrobial activity. Appl. Environ. Microbiol. 2009, 75, 1129–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández de Ullivarri, M.; Mendoza, L.M.; Raya, R.R. Characterization of the killer toxin KTCf20 from Wickerhamomyces anomalus, a potential biocontrol agent against wine spoilage yeasts. Biol. Control 2018, 121, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.A.; Sáez, J.S.; Sangorrín, M.P. Differential response of Pichia guilliermondii spoilage isolates to biological and physicochemical factors prevailing in Patagonian wine fermentations. Can. J. Microbiol. 2009, 55, 801–809. [Google Scholar] [CrossRef]
- Csutak, O.; Vassu, T.; Corbu, V.; Cîrpici, I.; Ionescu, R. Killer activity of Pichia anomala CMGB 88. Biointerface Res. Appl. Chem. 2017, 7, 2085–2089. [Google Scholar]
- Villalba, M.L.; Mazzucco, M.B.; Lopes, C.A.; Ganga, M.A.; Sangorrín, M.P. Purification and characterization of Saccharomyces eubayanus killer toxin: Biocontrol effectiveness against wine spoilage yeasts. Int. J. Food Microbiol. 2020, 331, 108714. [Google Scholar] [CrossRef]
- Türkel, S.; Ener, B. Isolation and characterization of new Metschnikowia pulcherrima strains as producers of the antimicrobial pigment pulcherrimin. Z. Naturforsch. C 2009, 64, 405–410. [Google Scholar]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Charpentier, C.; Escot, S.; Gonzalez, E.; Dulau, L.; Feuillat, M. The influence of yeast glycosylated proteins on tannins aggregation in model solution. J. Int. Sci. Vigne Vin 2004, 38, 209–218. [Google Scholar] [CrossRef]
- Buerth, C.; Heilmann, C.J.; Klis, F.M.; de Koster, C.G.; Ernst, J.F.; Tielker, D. Growth-dependent secretome of Candida utilis. Microbiology 2011, 157, 2493–2503. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main cell components of non-Saccharomyces yeasts with the potential for use in wine production.
Figure 1.
Main cell components of non-Saccharomyces yeasts with the potential for use in wine production.


{kind=link}
{kind=link}
Table 1.
Some non-Saccharomyces yeasts with the potential to release mannoproteins.
| Yeast | Protein and Monosaccharide Content in Mannoproteins [25] | Nitrogen Requirement | |||
|---|---|---|---|---|---|
| Protein (%) a | Mannose (%) b | Glucose (%) b | Galactose (%) b | ||
| Saccharomyces cerevisiae | 24 | 88 | 12 | - | |
| Metschnikowia pulcherrima | 25 | 86 | 14 | - | Slow ammonium uptake [4]. Weak or no growth in nitrate agar, and unable to develop in YPD agar at 37 °C [28]. |
| Wickerhamomyces anomalusc | 9 | 74 | 26 | - | Capable to uptake nitrate [36]. |
| Saccharomycodes ludwigiid | 12 | 93 | 7 | - | Unable to uptake nitrate [37]. Capable to uptake cadaverine and ethylamine [37]. |
| Schizosaccharomyces pombee | 11 | 55 | 22 | 23 | |
| Starmerella bombicolaf | 14 | 73 | 27 | - | |
| Pichia fermentans | 15 | 87 | 13 | - | |
| Hanseniaspora uvarumg | 23 | 81 | 19 | - | |
| Hanseniaspora valbyensis | 20 | 75 | 25 | - | |
| Lachancea thermotolerans | 16 | 82 | 18 | - | |
| Torulaspora delbrueckii | 18 | 85 | 15 | - | |
| Zygosaccharomyces bailii | 29 | 79 | 21 | - | |
| Brettanomyces bruxellensis | 16 | 88 | 12 | - | |
a Percentage of dry matter. b Sugars (%) in the polysaccharide fraction. c Wickerhamomyces anomalus (formerly Pichia anomala). d Saccharomycodes ludwigii: high autolytic activity: polysaccharide-releasing [21,25]. e Schizosaccharomyces pombe: high autolytic activity: polysaccharide-releasing [21,38]. f Starmerella bombicola (formerly Candida stellata DBVPG 3827). g Hanseniaspora uvarum (formerly Kloeckera apiculata).
Table 2.
Summary of the enzymes produced by non-Saccharomyces yeasts for wine production.
| Enzyme | Yeast | Application |
|---|---|---|
| β-glucosidase | Lachancea thermotolerans | Release of terpenes and thiols from their precursors: improvement of the aromatic profile [83,84,85]. |
| Torulaspora delbrueckii | Release of thiols from their cysteinylated precursors: improvement of the aromatic profile [84]. | |
| Wickerhamomyces anomalus |
| |
| Metschnikowia pulcherrima | ||
| Candida stellata | Release of terpenes (β-myrcene, limonene, linalool, α-terpineol, and farnesol) from their glycosylated precursors: improvement of the aromatic profile [88]. | |
| Hanseniaspora uvarum |
| |
| Saccharomycodes ludwigii | Activity up to 46% higher than S. cerevisiae at 30 °C [91]. | |
| Aureobasidium pullulans | Release of terpenes from their glycosylated precursors: improvement of the aromatic profile [92,93]. | |
| Protease | Wickerhamomyces anomalus | Aspartic protease WaAPr1 excreted by W. anomalus 227 [94]. |
| Metschnikowia pulcherrima | ||
| Candida stellata | ||
| Hanseniaspora uvarum | Degradation of proteins: improvement of clarification and stabilization [89,97]. | |
| Lachancea thermotolerans, Torulaspora delbrueckii, Zygosaccharomyces bailii, Pichia kluyveri | ||
| Glucanase | Wickerhamomyces anomalus | |
| Schizosaccharomyces pombe | High mannoproteins-releasing during AOL [21,38,99]. | |
| Saccharomycodes ludwigii | High mannoproteins-releasing during AOL [21,25]. | |
| Lachancea thermotolerans, Metschnikowia pulcherrima, Debaryomyces hansenii | Release of mannoproteins [25,83]. | |
| Hydroxycinnamate decarboxylase (HCDC) | Metschnikowia pulcherrima, Hanseniaspora guilliermondii, Hanseniaspora opuntiae, Hanseniaspora vineae, Hanseniaspora clermontiae, Pichia guillermondii | Involved in the synthesis of vinylphenolic pyranoanthocyanins: improvement of color stability in red wines [83,101,102,103]. |
| Urease | Schizosaccharomyces pombe | |
| Carboxypeptidase | Aureobasidium pullulans | |
| Pectinase | Wickerhamomyces anomalus | Degradation of pectins: improvement of clarification and turbidity reduction [85]. |
| Metschnikowia pulcherrima |
| |
| Candida stellata | Degradation of pectins: improvement of clarification and turbidity reduction [85,97]. | |
| Hanseniaspora uvarum | Degradation of pectins: improvement of clarification and turbidity reduction [89,97]. | |
| Aureobasidium pullulans |
| |
| Cellulase | Lachancea thermotolerans, Metschnikowia pulcherrima, Candida stellata, Hanseniaspora uvarum, Aureobasidium pullulans, Debaryomyces hansenii |
|
| Xylanase | Lachancea thermotolerans, Candida stellata, Hanseniaspora uvarum, Aureobasidium pullulans | Degradation of hemicellulose: improvement of wine aroma by increasing of monoterpenyl diglycoside precursors in the grape-must [83,92,97]. |
| β-lyase | Torulaspora delbrueckii, Kluyveromyces marxianus, Meyerozyma guilliermondii (formerly Pichia guilliermondii) | Release of thiols from their cysteinylated precursors: improvement of the aromatic profile [109,110]. |
| Lipase | Lachancea thermotolerans | Increase on free fatty acids concentration [83]. |
| Aureobasidium pullulans | Improvement of wine aroma: synthesis of ethyl esters and ethyl acetates from lipid cleavage [111]. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vejarano, R. Non-Saccharomyces in Winemaking: Source of Mannoproteins, Nitrogen, Enzymes, and Antimicrobial Compounds. Fermentation 2020, 6, 76. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030076
AMA Style
Vejarano R. Non-Saccharomyces in Winemaking: Source of Mannoproteins, Nitrogen, Enzymes, and Antimicrobial Compounds. Fermentation. 2020; 6(3):76. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030076
Chicago/Turabian StyleVejarano, Ricardo. 2020. "Non-Saccharomyces in Winemaking: Source of Mannoproteins, Nitrogen, Enzymes, and Antimicrobial Compounds" Fermentation 6, no. 3: 76. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030076
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.