Chestnut Shells as Waste Material for Succinic Acid Production from Actinobacillus succinogenes 130Z

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biomass Pretreatment and Hydrolysis
2.2. Small Scale Bottle Fermentation Experiments
2.3. Batch Fermentations
2.4. Analysis of Substrates and Products of Fermentation
3. Results and Discussion
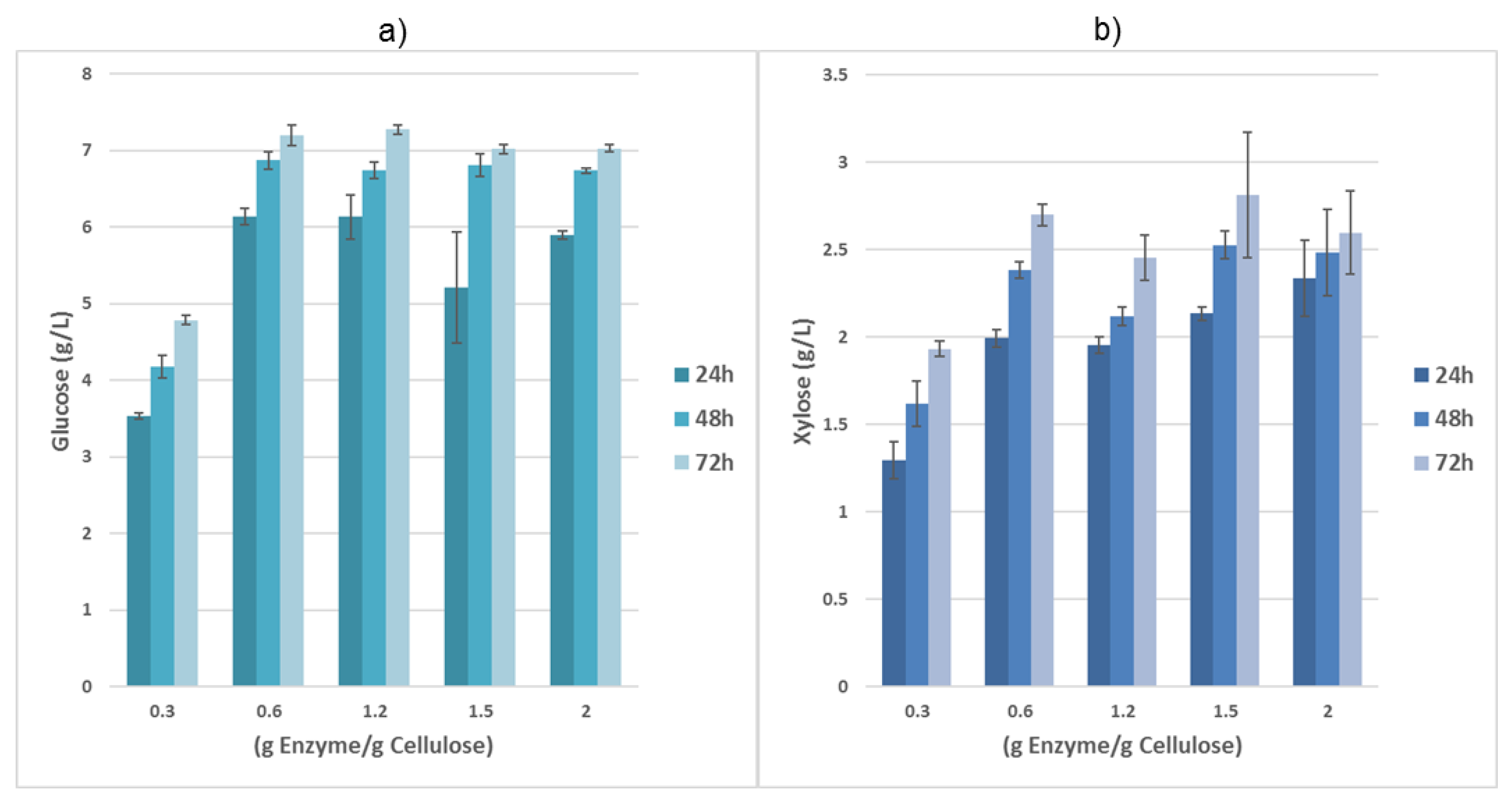
3.1. CSH Pretreatment and Hydrolysis
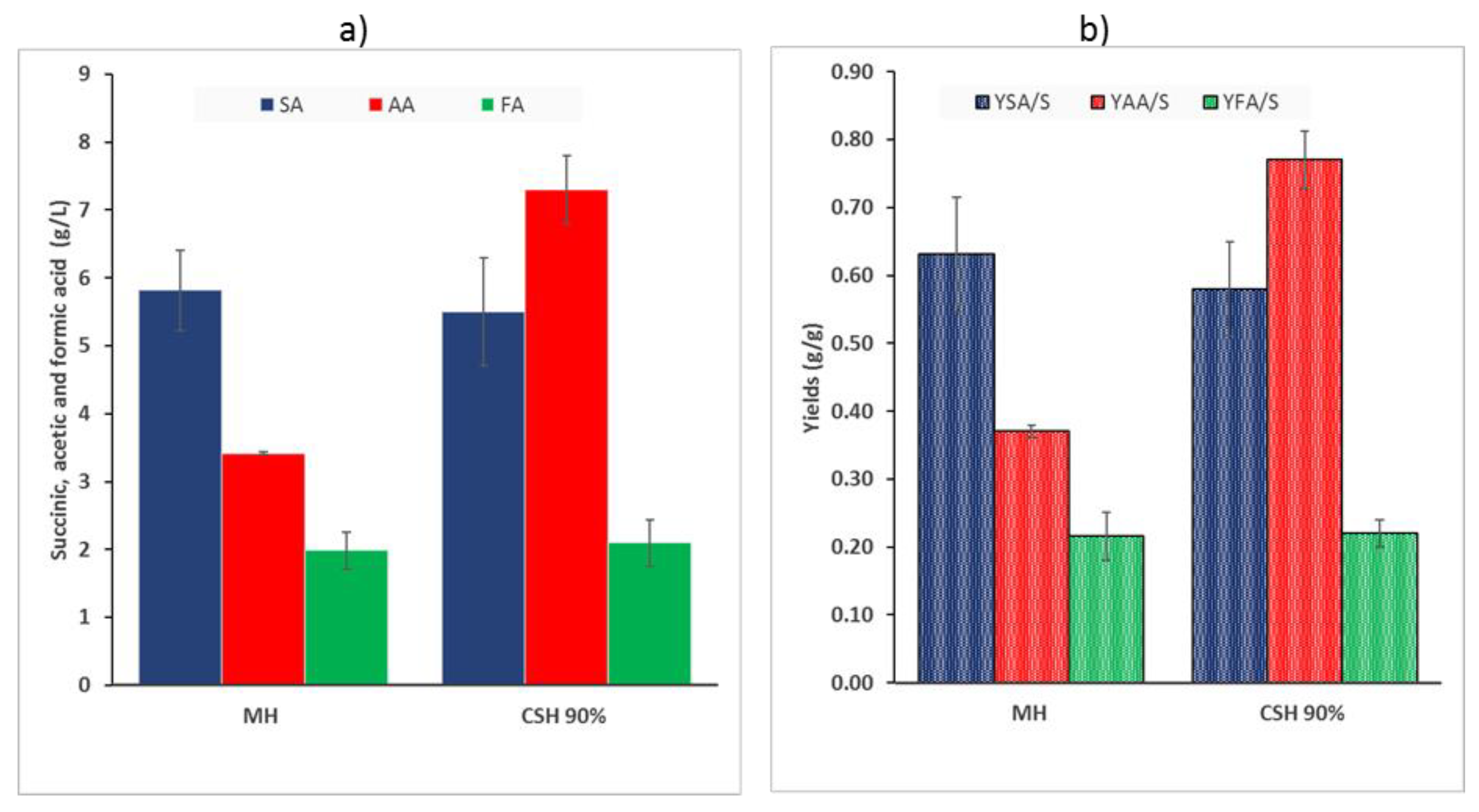
3.2. Bottle Experiments on Pretreated and Hydrolysed Chestnut Shells
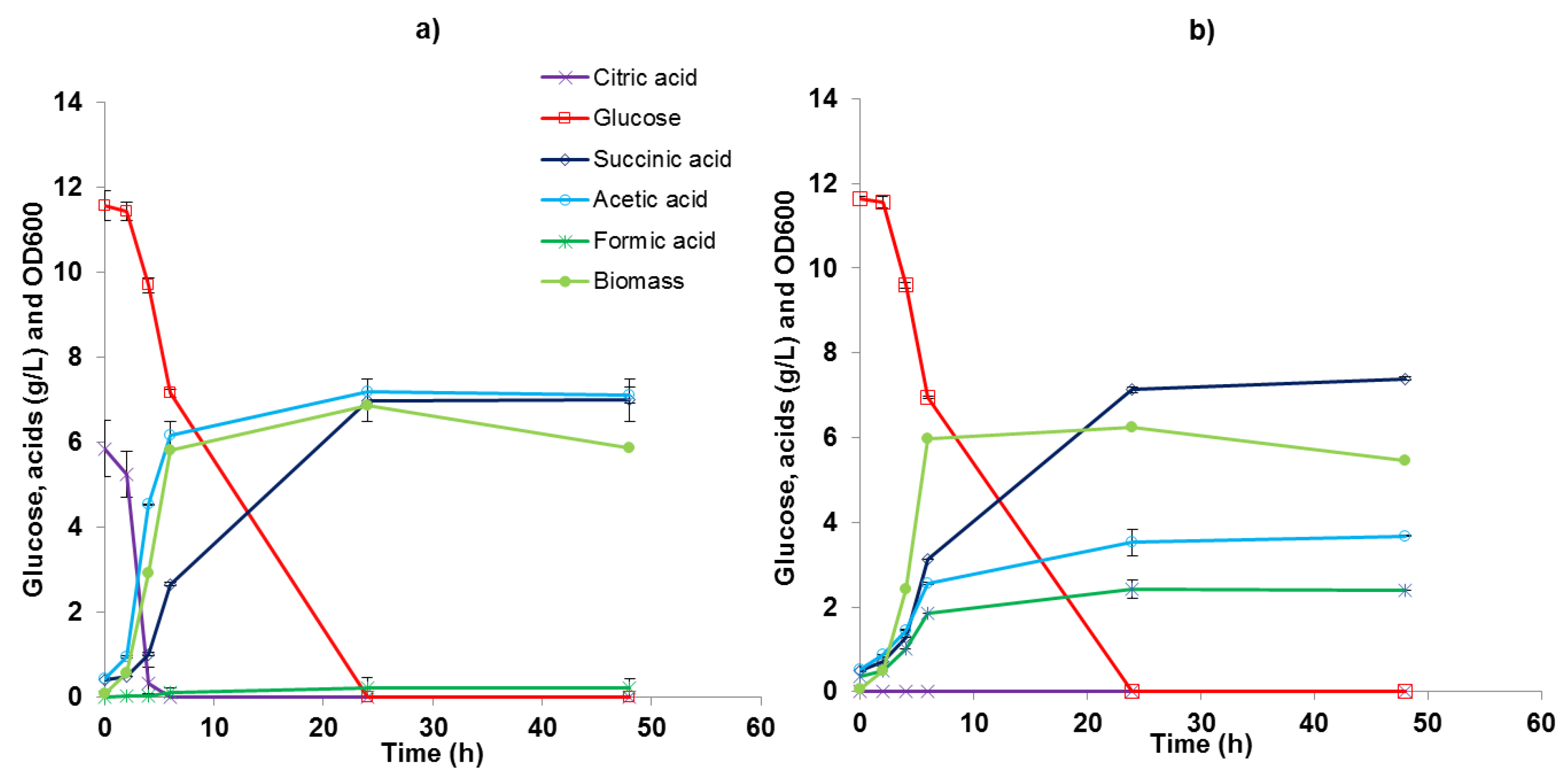
3.3. Batch Experiments in 2 L Bioreactors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Braga, N.; Rodrigues, F.; Oliveira, M.B.P.P. Castanea sativa by-products: A review on added value and sustainable application. Nat. Prod. Res. 2015, 29, 1–18. [Google Scholar] [CrossRef]
- Vázquez, G.; Fernández-Agulló, A.; Gómez-Castro, C.; Freire, M.S.; Antorrena, G.; González-Álvarez, J. Response surface optimization of antioxidants extraction from chestnut (Castanea sativa) bur. Ind. Crop. Prod. 2012, 35, 126–134. [Google Scholar] [CrossRef]
- Vázquez, G.; González-Alvarez, J.; Santos, J.; Freire, M.S.; Antorrena, G. Evaluation of potential applications for chestnut (Castanea sativa) shell and eucalyptus (Eucalyptus globulus) bark extracts. Ind. Crop. Prod. 2009, 29, 364–370. [Google Scholar] [CrossRef]
- Maurelli, L.; Ionata, E.; La Cara, F.; Morana, A. Chestnut Shell as Unexploited Source of Fermentable Sugars: Effect of Different Pretreatment Methods on Enzymatic Saccharification. Appl. Biochem. Biotechnol. 2013, 170, 1104–1118. [Google Scholar] [CrossRef] [PubMed]
- Morana, A.; Squillaci, G.; Paixão, S.; Alves, L.; Cara, F.; Moura, P. Development of an Energy Biorefinery Model for Chestnut (Castanea sativa Mill.) Shells. Energies 2017, 10, 1504. [Google Scholar] [CrossRef] [Green Version]
- Eryasar, K.; Karasu-Yalcin, S. Evaluation of some lignocellulosic byproducts of food industry for microbial xylitol production by Candida tropicalis. 3 Biotech 2016, 6, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, challenges, and future perspectives of succinic acid production by Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Yi, J.; Kim, T.Y.; Choi, S.; Ahn, J.H.; Song, H.; Lee, M.H.; Lee, S.Y. Homo-succinic acid production by metabolically engineered Mannheimia succiniciproducens. Metab. Eng. 2016, 38, 409–417. [Google Scholar] [CrossRef]
- Lange, A.; Becker, D.J.; Schulze, D.; Cahoreau, E.; Portais, J.; Haefner, S.; Schröder, H.; Krawczyk, J.; Zelder, O.; Wittmann, C. Bio-based succinate from sucrose: High-resolution 13 C metabolic flux analysis and metabolic engineering of the rumen bacterium Basfia succiniciproducens. Metab. Eng. 2017, 44, 198–212. [Google Scholar] [CrossRef]
- Ahn, J.H.; Seo, H.; Park, W.; Seok, J.; Lee, J.A.; Kim, W.J.; Kim, G.B.; Kim, K.; Lee, S.Y. Enhanced succinic acid production by Mannheimia employing optimal malate dehydrogenase. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, J.; Idris, A.; Abd, A.R. Recent advances in production of succinic acid from lignocellulosic biomass. Appl. Microbiol. Biotechnol. 2014, 98, 987–1000. [Google Scholar] [CrossRef]
- Salvachúa, D.; Smith, H.; John, P.C.S.; Mohagheghi, A.; Peterson, D.J.; Black, B.A.; Dowe, N.; Beckham, G.T. Succinic acid production from lignocellulosic hydrolysate by Basfia succiniciproducens. Bioresour. Technol. 2016, 214, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Cimini, D.; Zaccariello, L.; D’Ambrosio, S.; Lama, L.; Ruoppolo, G.; Pepe, O.; Faraco, V.; Schiraldi, C. Improved production of succinic acid from Basfia succiniciproducens growing on A. donax and process evaluation through material low analysis. Biotechnol. Biofuels 2019, 12, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.B.; Blandino, A.; Caro, I. Value added products from fermentation of sugars derived from agro-food residues. Trends Food Sci. Technol. 2018, 71, 52–64. [Google Scholar] [CrossRef]
- Ju, X.; Bowden, M.; Engelhard, M.; Zhang, X. Investigating commercial cellulase performances toward specific biomass recalcitrance factors using reference substrates. Appl. Microbiol. Biotechnol. 2014, 98, 4409–4420. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Haven, M.Ø.; Lindedam, J.; Felby, C.; Gama, M. Celluclast and Cellic® CTec2: Saccharification/fermentation of wheat straw, solid–liquid partition and potential of enzyme recycling by alkaline washing. Enzym. Microb. Technol. 2015, 79–80, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Baral, P.; Jain, L.; Kurmi, A.K.; Kumar, V.; Agrawal, D. Augmented hydrolysis of acid pretreated sugarcane bagasse by PEG 6000 addition: A case study of Cellic CTec2 with recycling and reuse. Bioproc. Biosyst. Eng. 2020, 43, 473–482. [Google Scholar] [CrossRef]
- Gunnarsson, I.B.; Kuglarz, M.; Karakashev, D.; Angelidaki, I. Thermochemical pretreatments for enhancing succinic acid production from industrial hemp (Cannabis sativa L.). Bioresour. Technol. 2015, 182, 58–66. [Google Scholar] [CrossRef]
- Cimini, D.; Argenzio, O.; D’Ambrosio, S.; Lama, L.; Finore, I.; Finamore, R.; Pepe, O.; Faraco, V.; Schiraldi, C. Production of succinic acid from Basfia succiniciproducens up to the pilot scale from Arundo donax hydrolysate. Bioresour. Technol. 2016, 222, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Ventrone, M.; Schiraldi, C.; Cimini, D. Physiology of Actinobacillus succinogenes 130 Z on MH and on chestnut shell hydrolysate. Unpublished work. 2020; Material not intended for publication. [Google Scholar]
- Joshi, R.V.; Schindler, B.D.; McPherson, N.R.; Tiwari, K.; Vieille, C. Development of a Markerless Knockout Method for Actinobacillus succinogenes. Appl. Environ. Microbiol. 2014, 80, 3053–3061. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-P.; Zheng, P.; Sun, Z.-H.; Ni, Y.; Dong, J.-J.; Wei, P. Strategies of pH control and glucose-fed batch fermentation for production of succinic acid byActinobacillus succinogenes CGMCC1593. J. Chem. Technol. Biotechnol. 2008, 83, 722–729. [Google Scholar] [CrossRef]
- Bailey, J.E.; Ollis, D.F. Biochemical Engineering Fundamentals, 2nd ed.; McGraw-Hill Chemical Engineering Series; McGraw-Hill: New York, NY, USA, 1986; ISBN 9780070032125. [Google Scholar]
- Maslova, O.; Stepanov, N.; Senko, O.; Efremenko, E. Production of various organic acids from different renewable sources by immobilized cells in the regimes of separate hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SFF). Bioresour. Technol. 2019, 272, 1–9. [Google Scholar] [CrossRef]
- Wang, Z.; Li, H.; Feng, J.; Zhang, A.; Ying, H.; He, X.; Jiang, M.; Chen, K.; Ouyang, P. Enhanced succinic acid production from polyacrylamide-pretreated cane molasses in microbial electrolysis cells. J. Chem. Technol. Biotechnol. 2018, 93, 855–860. [Google Scholar] [CrossRef]
- Li, Q.; Yang, M.; Wang, D.; Li, W.; Wu, Y.; Zhang, Y.; Xing, J.; Su, Z. Efficient conversion of crop stalk wastes into succinic acid production by Actinobacillus succinogenes. Bioresour. Technol. 2010, 101, 3292–3294. [Google Scholar] [CrossRef]
- Patsalou, M.; Menikea, K.K.; Makri, E.; Vasquez, M.I.; Drouza, C.; Koutinas, M. Development of a citrus peel-based biorefinery strategy for the production of succinic acid. J. Clean. Prod. 2017, 166, 706–716. [Google Scholar] [CrossRef]
- Lee, P.C.; Lee, W.G.; Lee, S.Y.; Chang, H.N. Effects of medium components on the growth of Anaerobiospirillum succiniciproducens and succinic acid production. Process. Biochem. 1999, 35, 49–55. [Google Scholar] [CrossRef]
- Lee, P.C.; Lee, W.G.; Lee, S.Y.; Chang, H.N. Succinic acid production with reduced by-product formation in the fermentation of Anaerobiospirillum succiniciproducens using glycerol as a carbon source. Biotechnol. Bioeng. 2001, 72, 41–48. [Google Scholar] [CrossRef]
- Baniel, A.M.; Eyal, A.M. Citric Acid Extraction. U.S. Patent 5,426,220, 20 June 1995. [Google Scholar]
- Zhang, W.; Yang, Q.; Wu, M.; Liu, H.; Zhou, J.; Dong, W.; Ma, J.; Jiang, M.; Xin, F. Metabolic Regulation of Organic Acid Biosynthesis in Actinobacillus succinogenes. Front. Bioeng. Biotechnol. 2019, 7, 216–226. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Time (h) | Glucose (g/L) | Cellulose Conversion (%) | Xylose (g/L) | Hemicellulose Conversion (%) |
|---|---|---|---|---|
| 24 | 6.2 ± 0.1 | 52 ± 1 | 2.0 ± 0.0 | 53 ± 1 |
| 48 | 6.9 ± 0.1 | 58 ± 1 | 2.4 ± 0.0 | 63 ± 1 |
| 72 | 7.2 ± 0.1 | 61 ± 1 | 2.7 ± 0.1 | 71 ± 2 |
| YSA/Gl (g/g) | YAA/Gl (g/g) | YFA/Gl (g/g) | |
|---|---|---|---|
| MH | 0.64 ± 0.01 | 0.39 ± 0.07 | 0.27 ± 0.05 |
| MH + Citrate | 0.60 ± 0.05 | 0.61 ± 0.01 | 0.30 ± 0.02 |
| SA (g/L) | YSA/Gl (g/g) | YAA/Gl (g/g) | YFA/Gl (g/g) | Yx/Gl (g/g) | pHfin | Glucose Exhaustion | |
|---|---|---|---|---|---|---|---|
| Glucose 10 g/L, MgCO3 10 g/L | 6.3 ± 0.8 | 0.64 ± 0.01 | 0.39 ± 0.07 | 0.27 ± 0.05 | 0.70 ± 0.14 | 6.0 ± 0.1 | Between 11 and 24 h |
| Glucose 20 g/L, MgCO3 20 g/L | 13.4 ± 1.0 | 0.76 ± 0.05 | 0.38 ± 0.06 | 0.24 ± 0.05 | 0.66 ± 0.10 | 6.2 ± 0.2 | Between 11 and 24 h |
| Glucose 50 g/L, MgCO3 50 g/L | 33.9 ± 1.9 | 0.72 ± 0.04 | 0.14 ± 0.01 | 0.10 ± 0.01 | 0.28 ± 0.01 | 6.4 ± 0.1 | Between 55 and 72 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventrone, M.; Schiraldi, C.; Squillaci, G.; Morana, A.; Cimini, D. Chestnut Shells as Waste Material for Succinic Acid Production from Actinobacillus succinogenes 130Z. Fermentation 2020, 6, 105. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040105
Ventrone M, Schiraldi C, Squillaci G, Morana A, Cimini D. Chestnut Shells as Waste Material for Succinic Acid Production from Actinobacillus succinogenes 130Z. Fermentation. 2020; 6(4):105. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040105
Chicago/Turabian StyleVentrone, Michela, Chiara Schiraldi, Giuseppe Squillaci, Alessandra Morana, and Donatella Cimini. 2020. "Chestnut Shells as Waste Material for Succinic Acid Production from Actinobacillus succinogenes 130Z" Fermentation 6, no. 4: 105. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040105