Single and Multiple Inoculum of Lactiplantibacillus plantarum Strains in Table Olive Lab-Scale Fermentations

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Media, and Growth Conditions
- L. plantarum B1, isolated from olive brines [10].
2.2. Probiotic Characterization of the Starter Cultures
2.2.1. Growth at Acidic pH
2.2.2. Bile Salt Hydrolase (BSH) Activity
2.2.3. Antibiotic Susceptibility
2.2.4. Exopolysaccharide (EPS) Production
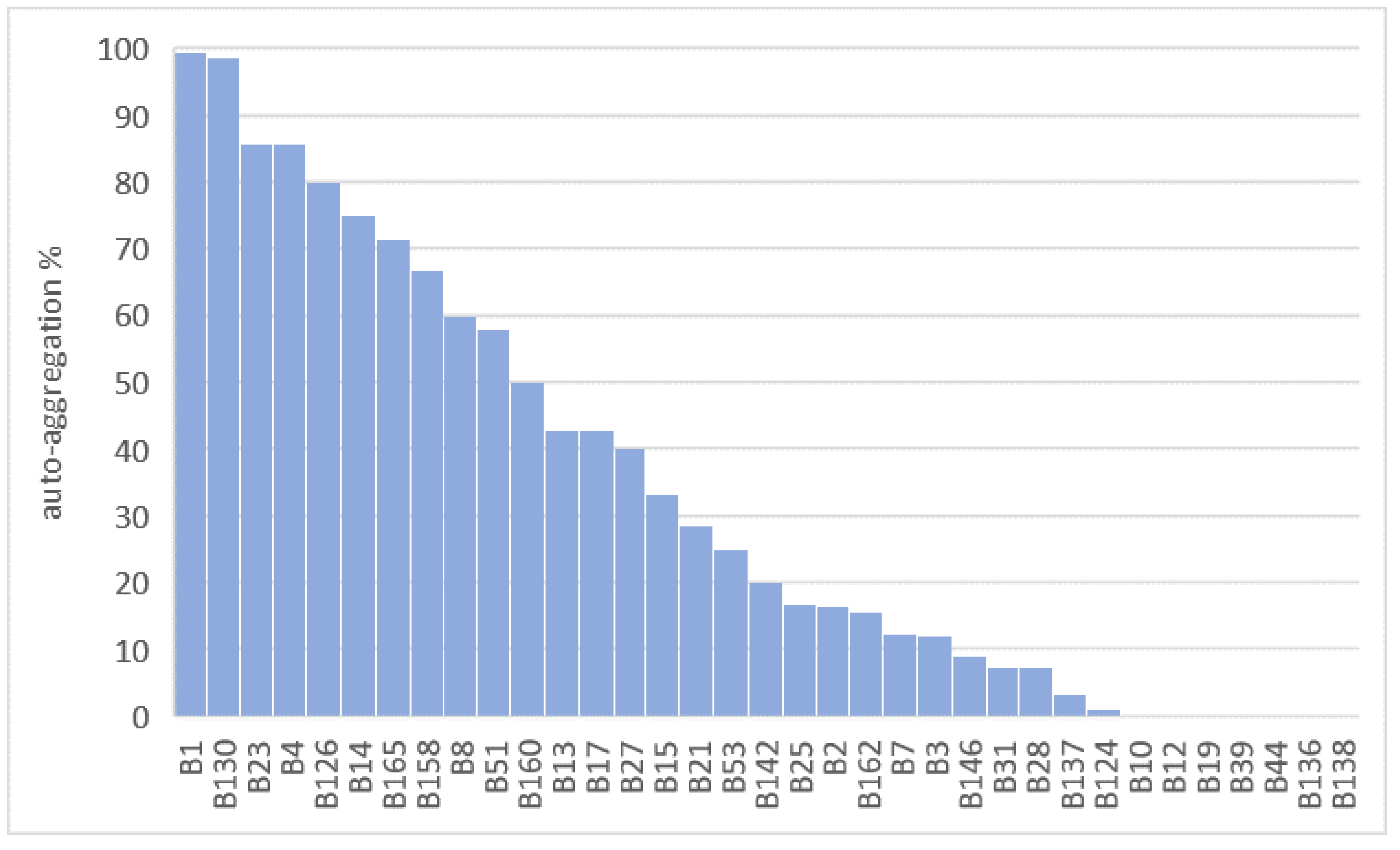
2.2.5. Auto-Aggregation Assay
2.3. Table Olive Processing
2.3.1. Raw Olives and Microorganisms
2.3.2. Preparation of the Inocula
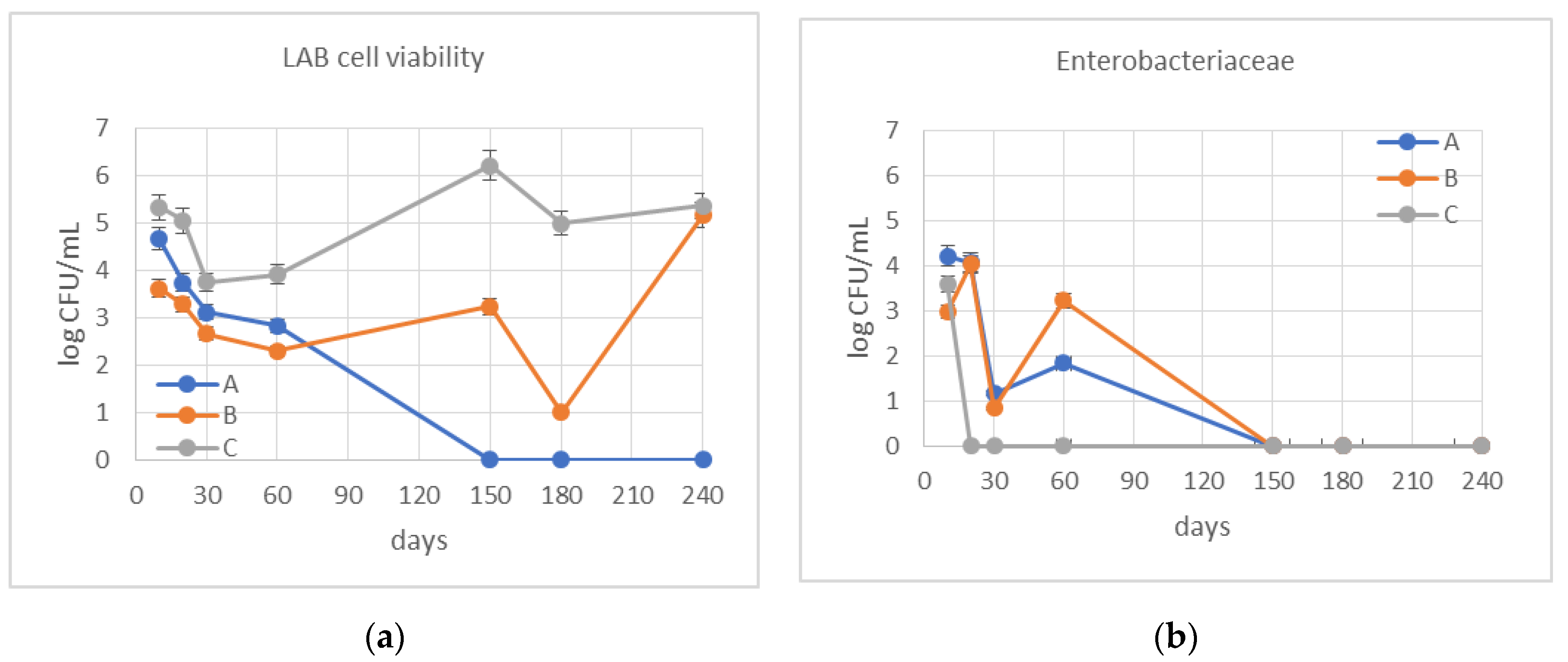
2.3.3. Determination of the Cell Viability during the Fermentation
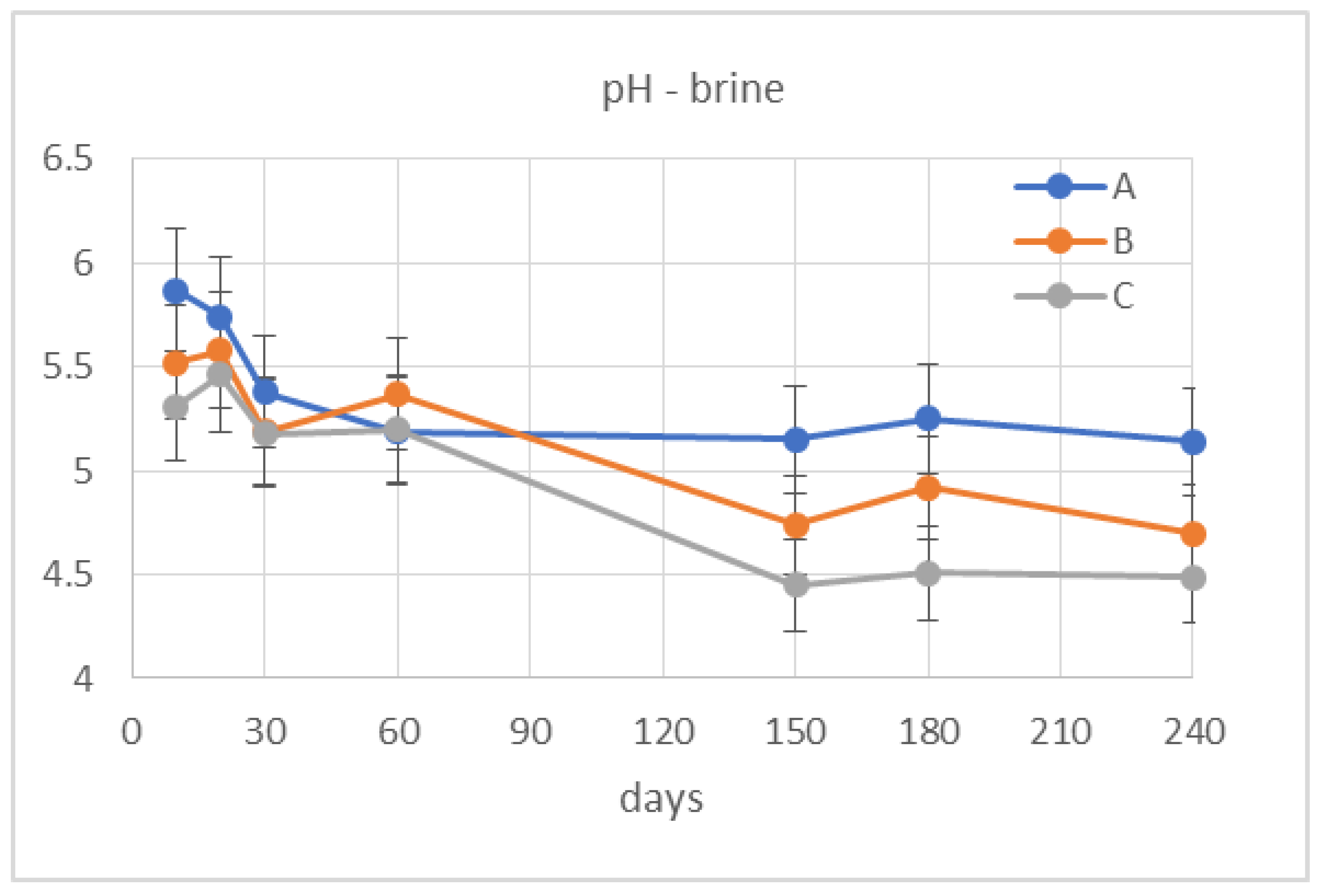
2.3.4. Evaluation of the Physicochemical Properties of the Fermentation Brines
2.3.5. Evaluation of the Physicochemical Properties of the Olive Juice
2.4. Biophenols Composition by HPLC
3. Results
3.1. Probiotic Characterization of Starter Cultures
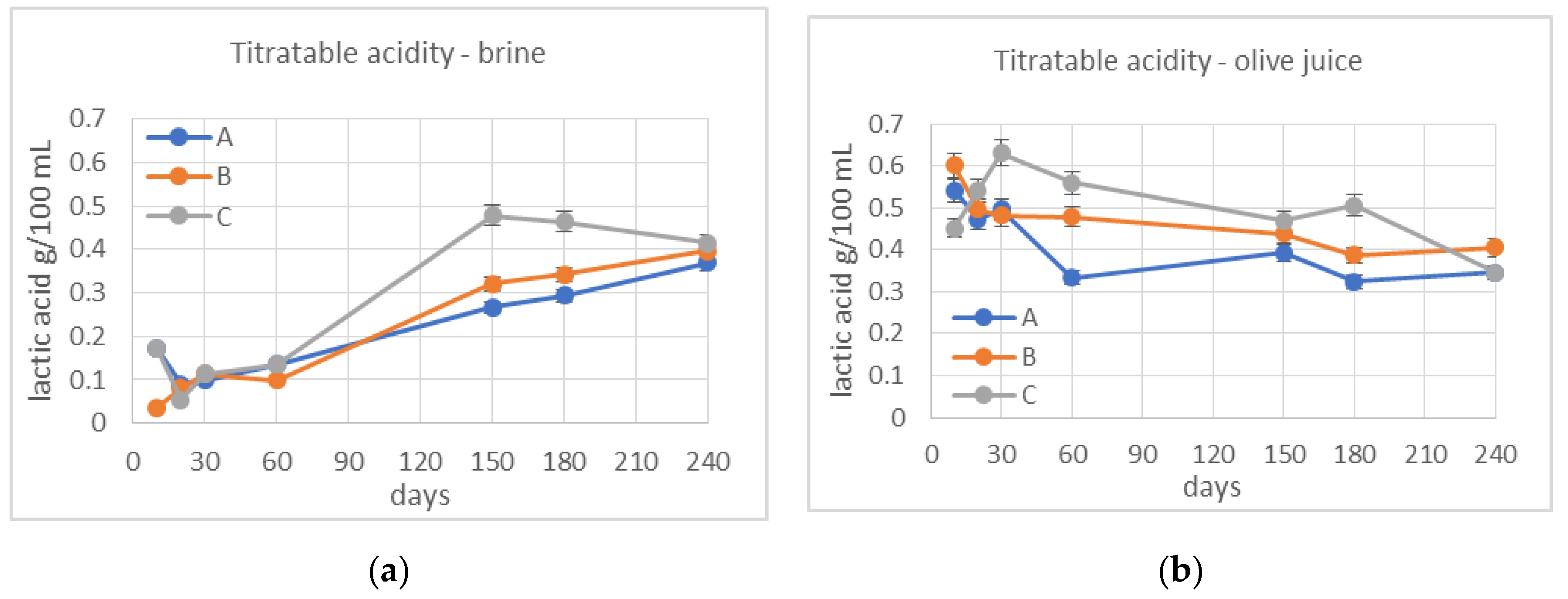
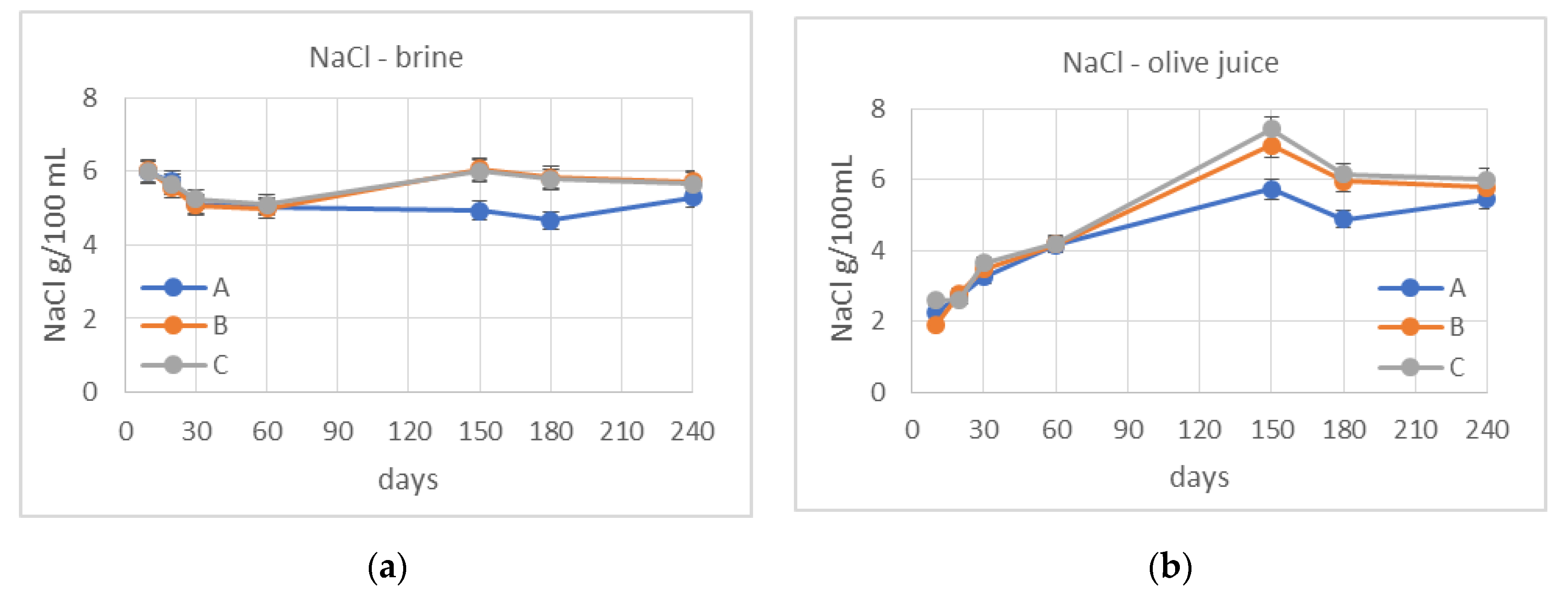
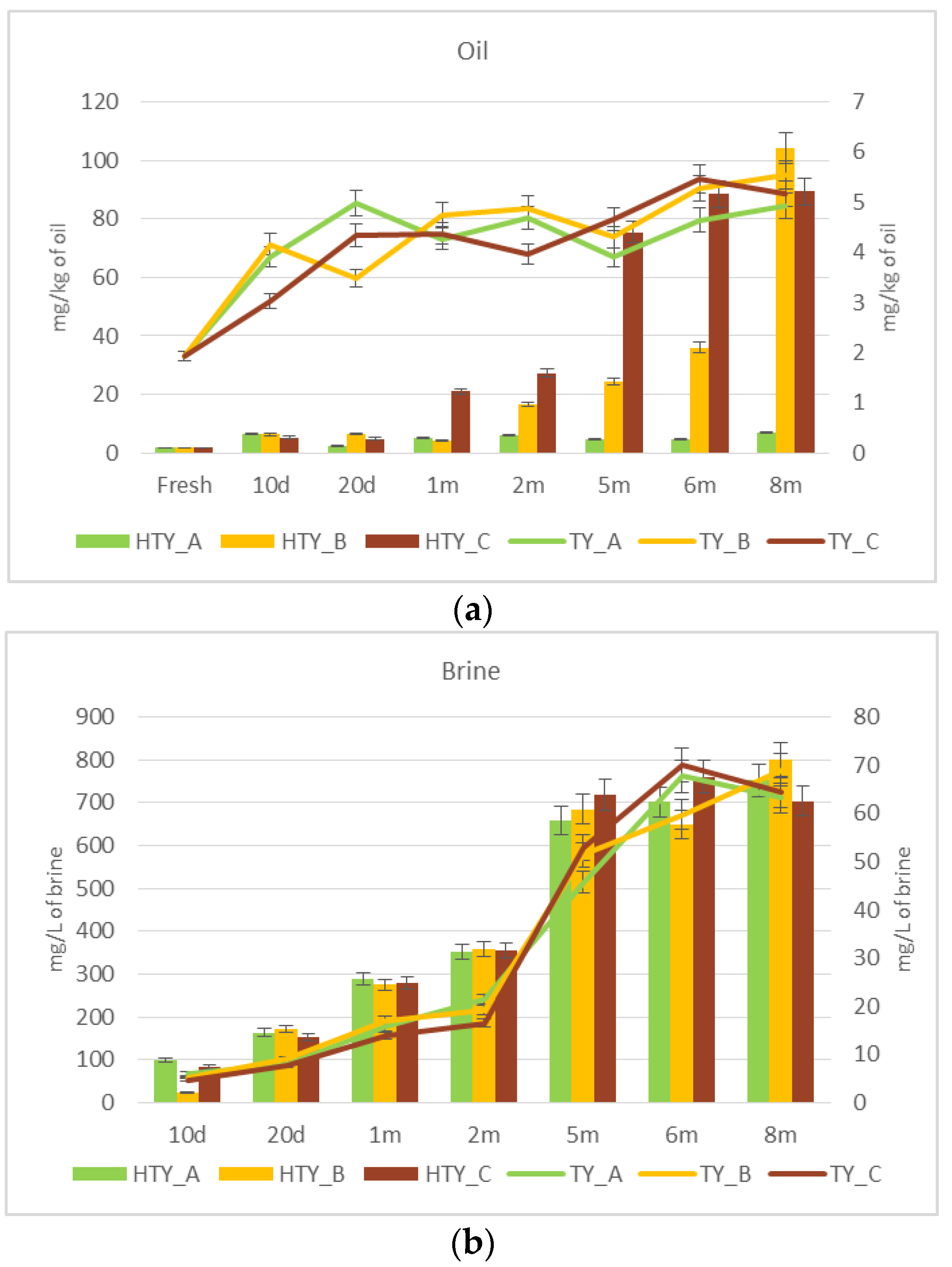
3.2. Monitoring of Fermentation
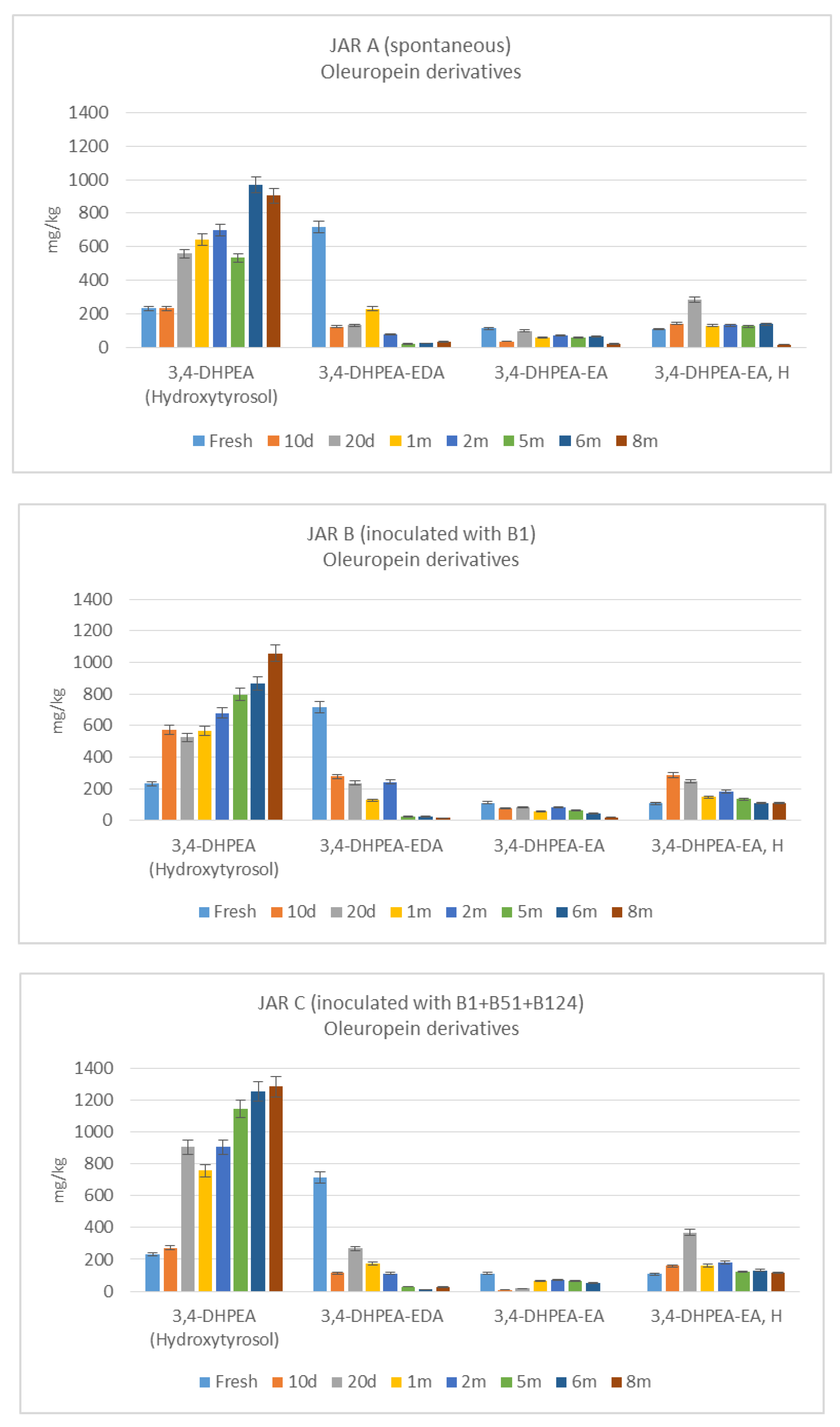
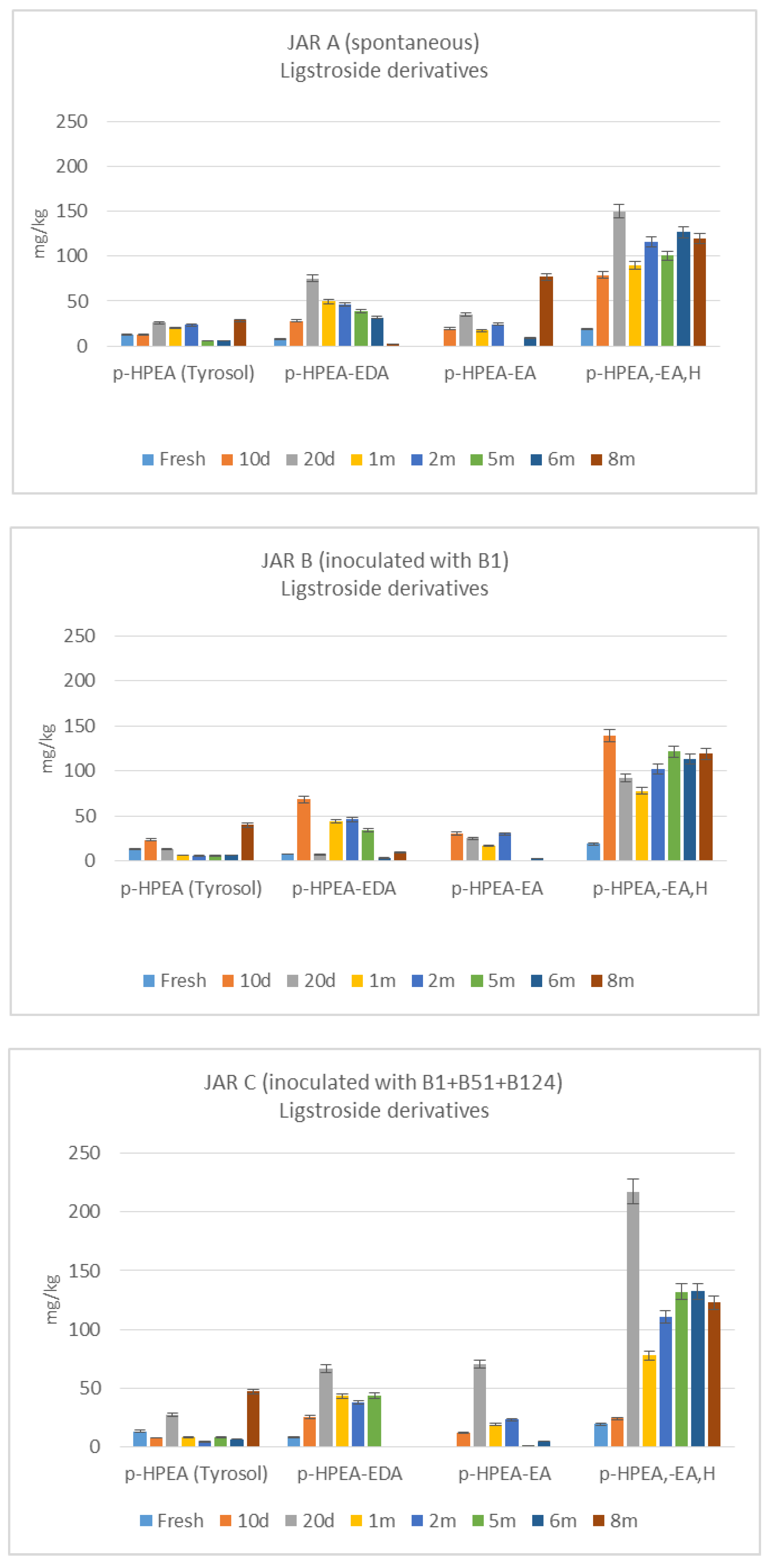
3.3. De-Bittering Process
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Perpetuini, G.; Prete, R.; Garcia-Gonzales, N.; Khairul Alam, M.; Corsetti, A. Table olives more than a fermented food. Foods 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagou, E.Z. Greek table olives processing. In Handbook of Vegetable Preservation and Processing, 2nd ed.; Hui, Y.H., Özgül Evranuz, E., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 547–550. [Google Scholar]
- Lavermicocca, P.; Rossi, M.; Russo, F.; Srirajaskanthan, R. Table olives: A carrier for delivering probiotic bacteria to humans. In Olives and Olive Oil in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press: London, UK, 2010; pp. 735–743. [Google Scholar]
- Botta, C.; Langerholc, T.; Cencic, A.; Cocolin, L. In vitro selection and characterization of new probiotic candidates from table olive microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro test. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Gallego, J.; Arroyo-Lopez, F.N.; Rantsiou, K.; Jimenez-Diaz, R.; Garrido-Fernandez, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Guantario, B.; Zinno, P.; Schifano, E.; Rosselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In vitro and in vivo selection of potentially probiotic Lactobacilli from Nocellara del Belice table olives. Front. Microbiol. 2018, 9, 595. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, T.; Ramalhosa, E.; Nunes, L.; Pereira, J.A.; Colla, E.; Pereira, E.L. Probiotic potential of indigenous yeasts isolated during the fermentation of table olives from Northeast of Portugal. Innov. Food Sci. Emerg. Technol. 2017, 44, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Bonatsou, S.; Karamouza, M.; Zoumpopoulou, G.; Mavrogonatou, E.; Kletsas, D.; Papadimitriou, K.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z. Evaluating the probiotic potential and technological characteristics of yeasts implicated in cv. Kalamata natural black olive fermentation. Int. J. Food Microbiol. 2018, 271, 48–59. [Google Scholar] [CrossRef]
- Ciafardini, G.; Marsilio, V.; Lanza, B.; Pozzi, N. Hydrolysis of oleuropein by Lactobacillus plantarum strains associated with olive fermentation. Appl. Environ. Microbiol. 1994, 60, 4142–4147. [Google Scholar] [CrossRef] [Green Version]
- Zago, M.; Lanza, B.; Rossetti, L.; Muzzalupo, I.; Carminati, D.; Giraffa, G. Selection of Lactobacillus plantarum strains to use as starters in fermented table olives: Oleuropeinase activity and phage sensitivity. Food Microbiol. 2013, 34, 81–87. [Google Scholar] [CrossRef]
- Pino, A.; Vaccalluzzo, A.; Solieri, L.; Romeo, F.V.; Todaro, A.; Caggia, C.; Arroyo-López, F.N.; Bautista-Gallego, J.; Randazzo, C.L. Effect of sequential inoculum of beta-glucosidase positive and probiotic strains on brine fermentation to obtain low salt Sicilian table olives. Front. Microbiol. 2019. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Cathcart, D.P.; Warner, P.J.; Jimenez-Diaz, R. Use of Lactobacillus plantarum LPCO10, a bacteriocin producer, as a starter culture in Spanish-Style green olive fermentations. Appl. Environ. Microbiol. 1994, 60, 2059–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, M.D.; Delgado, A.M.; Catulo, L.; Quintans, F.; Perese, C. The role of bacteriocin producers in table-olive fermentation. In Proceedings of the 5th International Symposium on Olive Growing, Izmir, Turkey, 27 September–2 October 2004. [Google Scholar]
- Rubia-Soria, A.; Abriouel, H.; Lucas, R.; Ben Omar, N.; Martinez-Canamero, M.; Galvez, A. Production of antimicrobial substances by bacteria isolated from fermented table olives. World J. Microbiol. Biotechnol. 2006, 22, 765–768. [Google Scholar] [CrossRef]
- Santos, M.M.; Piccirillo, C.; Castro, P.M.L.; Kalogerakis, N.; Pintado, M.E. Bioconversion of oleuropein to hydroxytyrosol by lactic acid bacteria. World J. Microbiol. Biotechnol. 2012, 28, 2435–2440. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, E.; Brenes, M.; de Castro, A.; Romero, C.; Medina, E. Oleuropein hydrolysis by lactic acid bacteria in natural green olives. LWT Food Sci. Technol. 2017, 78, 165–171. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Arroyo-López, F.N.; Roldán-Reyes, J.C.; Torres-Gallardo, R.; Bautista-Gallego, J.; García-García, P.; Garrido-Fernández, A. Assessing the challenges in the application of potential probiotic lactic acid bacteria in the large-scale fermentation of Spanish-style table olives. Front. Microbiol. 2017, 8, 915. [Google Scholar] [CrossRef] [Green Version]
- Heperkan, D. Microbiota of table olive fermentations and criteria of selection for their use as starters. Front. Microbiol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Paba, A.; Chessa, L.; Daga, E.; Campus, M.; Bulla, M.; Angioni, A.; Sedda, P.; Comunian, R. Do best-selected strains perform table olive fermentation better than undefined biodiverse starters? A comparative study. Foods 2020, 9, E135. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food. 2002. Available online: www.fao.org (accessed on 5 April 2020).
- Parker, E.A.; Roy, T.; D’Adamo, C.R.; Wieland, L.S. Probiotics and gastrointestinal conditions: An overview of evidence from the Cochrane Collaboration. Nutrition 2018, 45, 125–134.e11. [Google Scholar] [CrossRef]
- Sniffen, J.C.; McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Choosing an appropriate probiotic product for your patient: An evidence-based practical guide. PLoS ONE 2018, 13, e0209205. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C.; Srigopalram, S. In vitro importance of probiotic Lactobacillus plantarum related to medical field. Saudi J. Biol. Sci. 2016, 23, S6–S10. [Google Scholar] [CrossRef] [Green Version]
- Lanza, B.; Zago, M.; Carminati, D.; Rossetti, L.; Meucci, A.; Marfisi, P.; Russi, F.; Iannucci, E.; Di Serio, M.G.; Giraffa, G. Isolation and preliminary characterization of Lactobacillus plantarum bacteriophages from table olive fermentation. Ann. Microbiol. 2012, 62, 1467–1472. [Google Scholar] [CrossRef]
- Tidona, F.; Zago, M.; Corredig, M.; Locci, F.; Contarini, G.; Giraffa, G.; Carminati, D. Selection of Streptococcus thermophilus strains able to produce exopolysaccharides in milk. Int. J. Dairy Technol. 2016, 69, 569–575. [Google Scholar] [CrossRef]
- CODEX STAN 66-1981 (rev. 2013). Standard for Table Olives. Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B66-1981%252FCXS_066e.pdf (accessed on 25 January 2020).
- Lanza, B.; Di Serio, M.G.; Russi, F.; Iannucci, E.; Giansante, L.; Di Loreto, G.; Di Giacinto, L. Evaluation of the nutritional value of oven-dried table olives (cv. Majatica) processed by the Ferrandina style. Riv. Ital. Sostanze Gr. 2014, 2, 117–127. [Google Scholar]
- IOC. Determination of Biophenols in Olive Oils by HPLC; COI/T.20/Doc No 29/Rev.1 2017; International Olive Council: Madrid, Spain, 2017. [Google Scholar]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Lanza, B.; Ninfali, P. Antioxidants in extra virgin olive oil and table olives: Connections between agriculture and processing for health choices. Antioxidants 2020, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L.; Moschetti, G. Use of selected autochthonous lactic acid bacteria for Spanish-style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef]
- Marsilio, V.; Lanza, B.; Pozzi, N. Progress in table olives debittering: Degradation in vitro of oleuropein and its derivatives by L. plantarum. J. Am. Oil Chem. Soc. 1996, 73, 593–597. [Google Scholar] [CrossRef]
- Marsilio, V.; Lanza, B. Characterization of an oleuropein degrading strain of Lactobacillus plantarum. Combined effect of compounds present in olive fermenting brines (phenols, glucose and NaCl) on bacterial activity. J. Sci. Food Agric. 1998, 76, 520–524. [Google Scholar] [CrossRef]
- Marsilio, V.; Seghetti, L.; Iannucci, E.; Russi, F.; Lanza, B.; Felicioni, M. Use of a lactic acid bacteria starter culture during green olive (Olea europaea L., cv. Ascolana tenera) processing. J. Sci. Food Agric. 2005, 85, 1084–1090. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K. Prebiotic–Probiotic Relationship: The Genetic Fundamentals of Polysaccharides Conversion by Bifidobacterium and Lactobacillus Genera. In Food Bioconversion, 1st ed.; Alexandru Grumezescu, A., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Lavermicocca, P.; Lonigro, S.L.; Visconti, A.; De Angelis, M.; Valerio, F.; Morelli, L. Table Olives Containing Probiotic Microrganisms; Applicant CNR; EP1843664 B1 (8.7.2009); Granted European Patent Office: München, Germany, 2009. [Google Scholar]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; Pinto, C.L.O.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Zabat, M.A.; Sano, W.H.; Wurster, J.I.; Cabral, D.J.; Belenky, P. Microbial Community Analysis of Sauerkraut Fermentation Reveals a Stable and Rapidly Established Community. Foods 2018, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, P.; Emanuilova, M.; Petrov, K. Amylolytic Lactobacillus strains from Bulgarian fermented beverage boza. Z. Naturforsch. C J. Biosci. 2010, 65, 218–224. [Google Scholar] [CrossRef]
- Ruiz-Barba, J.L.; Caballero-Guerrero, B.; Maldonado-Barragán, A.; Jiménez-Díaz, R. Coculture with specific bacteria enhances survival of Lactobacillus plantarum NC8, an autoinducer-regulated bacteriocin producer, in olive fermentations. Food Microbiol. 2010, 27, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, A.; Ben Othman, N.; Chammem, N.; Hamdi, M.; Ferrer, S.; Reguant, C.; Bordons, A.; Rozès, N. Characterization of Lactobacillus isolates from fermented olives and their bacteriocin gene profiles. Food Microbiol. 2011, 28, 1514–1518. [Google Scholar] [CrossRef] [PubMed]
- Pérez Montoro, B.; Benomar, N.; Lavilla Lerma, L.; Castillo Gutiérrez, S.; Gálvez, A.; Abriouel, H. Fermented aloreña table olives as a source of potential probiotic Lactobacillus pentosus strains. Front. Microbiol. 2016, 7, 1583. [Google Scholar]
- Anagnostopoulos, D.A.; Goulas, V.; Xenofontos, E.; Vouras, C.; Nikoloudakis, N.; Tsaltas, D. Benefits of the use of lactic acid bacteria starter in green cracked Cypriot table olives fermentation. Foods 2020, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Kaltsa, A.; Papaliaga, D.; Papaioannou, E.; Kotzekidou, P. Characteristics of oleuropeinolytic strains of Lactobacillus plantarum group and influence on phenolic compounds in table olives elaborated under reduced salt conditions. Food Microbiol. 2015, 48, 58–62. [Google Scholar] [CrossRef]
- Iorizzo, M.; Lombardi, S.J.; Macciola, V.; Testa, B.; Lustrato, G.; Lopez, F.; De Leonardis, A. Technological potential of Lactobacillus strains isolated from fermented green olives: In vitro studies with emphasis on oleuropein-degrading capability. Sci. World J. 2016, 2016, 1917592. [Google Scholar] [CrossRef] [Green Version]
- Servili, M.; Minnocci, A.; Veneziani, G.; Taticchi, A.; Urbani, S.; Esposto, S.; Sebastiani, L.; Valmorri, S.; Corsetti, A. Compositional and tissue modifications induced by the natural fermentation process in table olives. J. Agric. Food Chem. 2008, 56, 6389–6396. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Origin (Olea europaea L. cv.) |
|---|---|
| B1 | unknown |
| B2 | unknown |
| B3 | I-77 |
| B4 | Carolea |
| B7 | unknown |
| B8 | unknown |
| B10 | Ascolana tenera |
| B12 | Picholine |
| B13 | Picholine |
| B14 | Nocellara del Belice |
| B15 | Picholine |
| B17 | Leccino |
| B19 | Sant’Agostino |
| B21 | Sant’Agostino |
| B23 | Nocellara del Belice |
| B25 | Coratina |
| B27 | Coratina |
| B28 | Coratina |
| B31 | I-77 |
| B39 | Grossa di Cassano |
| B44 | Santa Caterina |
| B51 | Cucco |
| B53 | Cucco |
| B124 | Sant’Agostino |
| B126 | Nocellara del Belice |
| B130 | Ascolana tenera |
| B136 | Nocellara del Belice |
| B137 | Nocellara etnea |
| B138 | Bella di Cerignola |
| B142 | Intosso |
| B146 | Ascolana tenera |
| B158 | Sant’Agostino |
| B160 | Bella di Cerignola |
| B162 | Nocellara del Belice |
| B165 | Nocellara etnea |
| Strain | Growth at Acidic pH | Bile Salt Hydrolysis | Antibiotic Susceptibility (MIC *) | Exopolysaccharides Production | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH 2.0 | TDCA | GDCA | Clindamycin | Erythromycin | Gentamicin | Tetracycline | Chloramphenicol | Ampicillin | Kanamycin | Colony Mucosity | Culture Viscosity (mPa.s) | Visualization of EPSs by SEM | |
| B1 | +++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.11 | absent |
| B2 | +++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.06 | absent |
| B3 | + | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.06 | absent |
| B4 | ++ | − | − | 1 (S) | 0.5 (S) | 32 (R) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.18 | absent |
| B7 | ++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.21 | absent |
| B8 | ++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.09 | absent |
| B10 | ++ | − | − | 1 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.18 | absent |
| B12 | ++ | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.09 | absent |
| B13 | ++ | − | + | 1 (S) | 0.25 (S) | 32 (R) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.2 | absent |
| B14 | ++ | − | + | 1 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.1 | absent |
| B15 | − | − | − | 1 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.2 | absent |
| B17 | +++ | − | + | 2 (S) | 0.25 (S) | 32 (R) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.22 | absent |
| B19 | ++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (S) | 256 (S) | +/− | 1.3 | absent |
| B21 | +++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 4 (R) | 256 (R) | absent | 1.09 | absent |
| B23 | ++ | − | − | 1 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 4 (R) | 256 (R) | absent | 1.16 | absent |
| B25 | + | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 4 (R) | 256 (R) | absent | 1.09 | absent |
| B27 | ++ | − | + | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.08 | absent |
| B28 | + | − | − | 4 (R) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 512 (R) | +/− | 1.3 | absent |
| B31 | − | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.2 | absent |
| B39 | ++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.2 | absent |
| B44 | ++ | − | + | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.3 | absent |
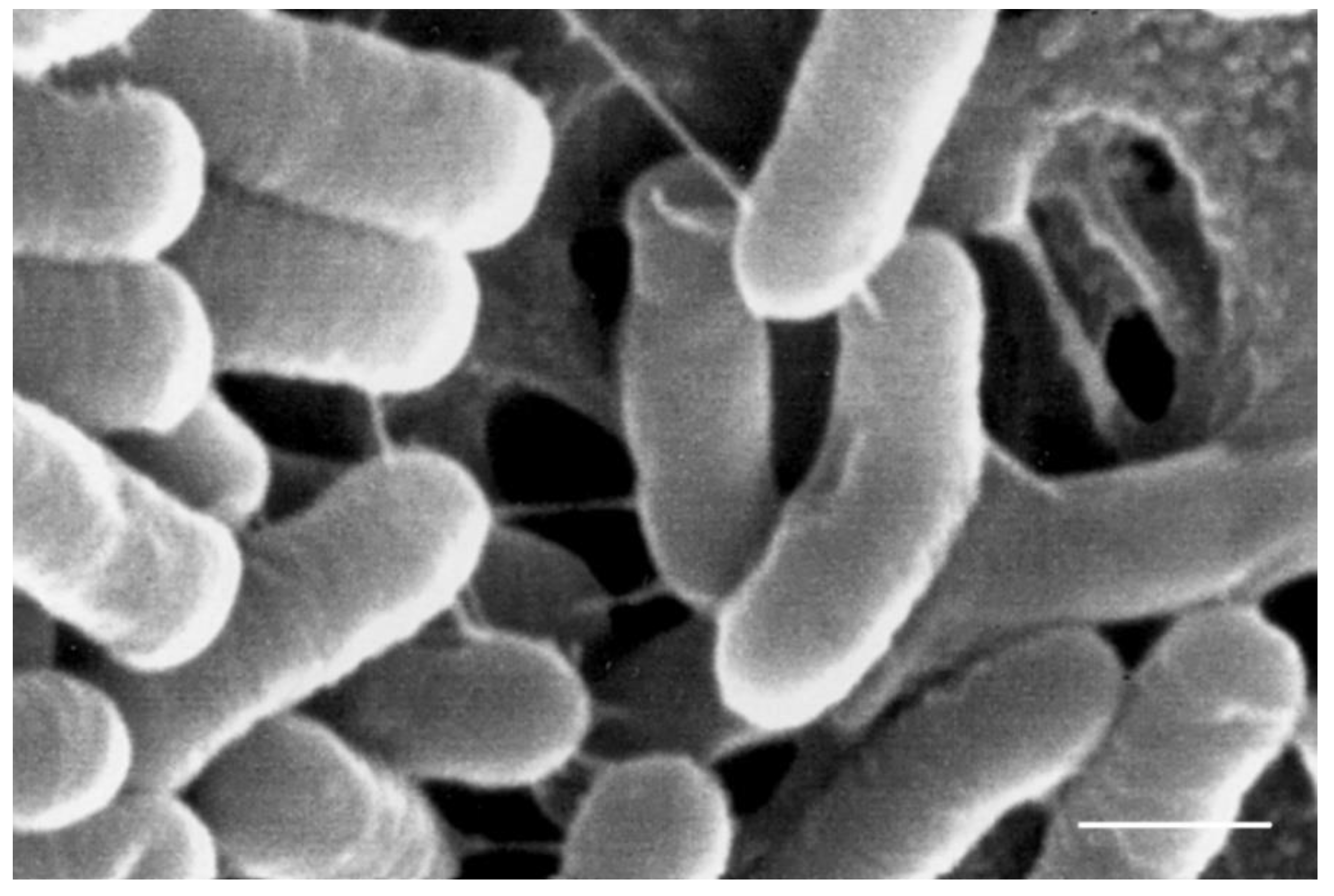
| B51 | ++ | ++ | ++ | 0.25 (S) | 0.5 (S) | 32 (R) | 32 (S) | 8 (S) | 16 (R) | 512 (R) | absent | 1.12 | [see Figure 1] |
| B53 | ++ | − | + | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.1 | absent |
| B124 | +++ | − | − | 2 (S) | 1 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.03 | absent |
| B126 | +++ | − | + | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.24 | absent |
| B130 | +++ | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.21 | absent |
| B136 | +++ | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 4 (S) | 8 (R) | 256 (R) | absent | 1.07 | absent |
| B137 | +++ | − | − | 2 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 4 (R) | 256 (R) | +/− | 1.12 | absent |
| B138 | +++ | − | + | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.17 | absent |
| B142 | +++ | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.17 | absent |
| B146 | ++ | − | + | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.17 | absent |
| B158 | ++ | − | + | 0.5 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.16 | absent |
| B160 | ++ | − | + | 1 (S) | 0.25 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.18 | absent |
| B162 | +++ | − | − | 1 (S) | 0.25 (S) | 17 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | absent | 1.52 | absent |
| B165 | +++ | − | − | 2 (S) | 0.5 (S) | 16 (S) | 32 (S) | 8 (S) | 8 (R) | 256 (R) | +/− | 1.15 | absent |
| Lp 790 | + | ++ | ++ | 0.25 (S) | 0.25 (S) | 16 (S) | 32 (S) | 4 (S) | 8 (R) | 64 (S) | absent | ||
| Lb 270 | + | 1.29 | absent | ||||||||||
| Flesh | Oil | Brine | ||||
|---|---|---|---|---|---|---|
| HTY | TY | HTY | TY | HTY | TY | |
| Jar A | 3.9 | 2.2 | 3.8 | 2.5 | 7.7 | 10.5 |
| Jar B | 4.6 | 3.1 | 57.8 | 2.9 | 33.3 | 13.3 |
| Jar C | 5.5 | 3.6 | 49.4 | 2.7 | 8.4 | 13.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanza, B.; Zago, M.; Di Marco, S.; Di Loreto, G.; Cellini, M.; Tidona, F.; Bonvini, B.; Bacceli, M.; Simone, N. Single and Multiple Inoculum of Lactiplantibacillus plantarum Strains in Table Olive Lab-Scale Fermentations. Fermentation 2020, 6, 126. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040126
Lanza B, Zago M, Di Marco S, Di Loreto G, Cellini M, Tidona F, Bonvini B, Bacceli M, Simone N. Single and Multiple Inoculum of Lactiplantibacillus plantarum Strains in Table Olive Lab-Scale Fermentations. Fermentation. 2020; 6(4):126. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040126
Chicago/Turabian StyleLanza, Barbara, Miriam Zago, Sara Di Marco, Giuseppina Di Loreto, Martina Cellini, Flavio Tidona, Barbara Bonvini, Martina Bacceli, and Nicola Simone. 2020. "Single and Multiple Inoculum of Lactiplantibacillus plantarum Strains in Table Olive Lab-Scale Fermentations" Fermentation 6, no. 4: 126. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040126