
Antioxidant Effect via Bioconversion of Isoflavonoid in Astragalus membranaceus Fermented by Lactiplantibacillus plantarum MG5276 In Vitro and In Vivo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Fermentation of Astragalus Membranaceus Extract
2.2. High Perfomance Liquid Chromatography (HPLC) Analysis
2.2.1. Pre-treatment of Samples for Analysis
2.2.2. Analysis of Isoflavonoids by HPLC
2.3. Cell Culture
2.4. Cell Viability
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Animal Treatments
2.7. Biochemical Parameters of Serum and Liver Tissues
2.8. Statistical Analysis
3. Results and Discussion
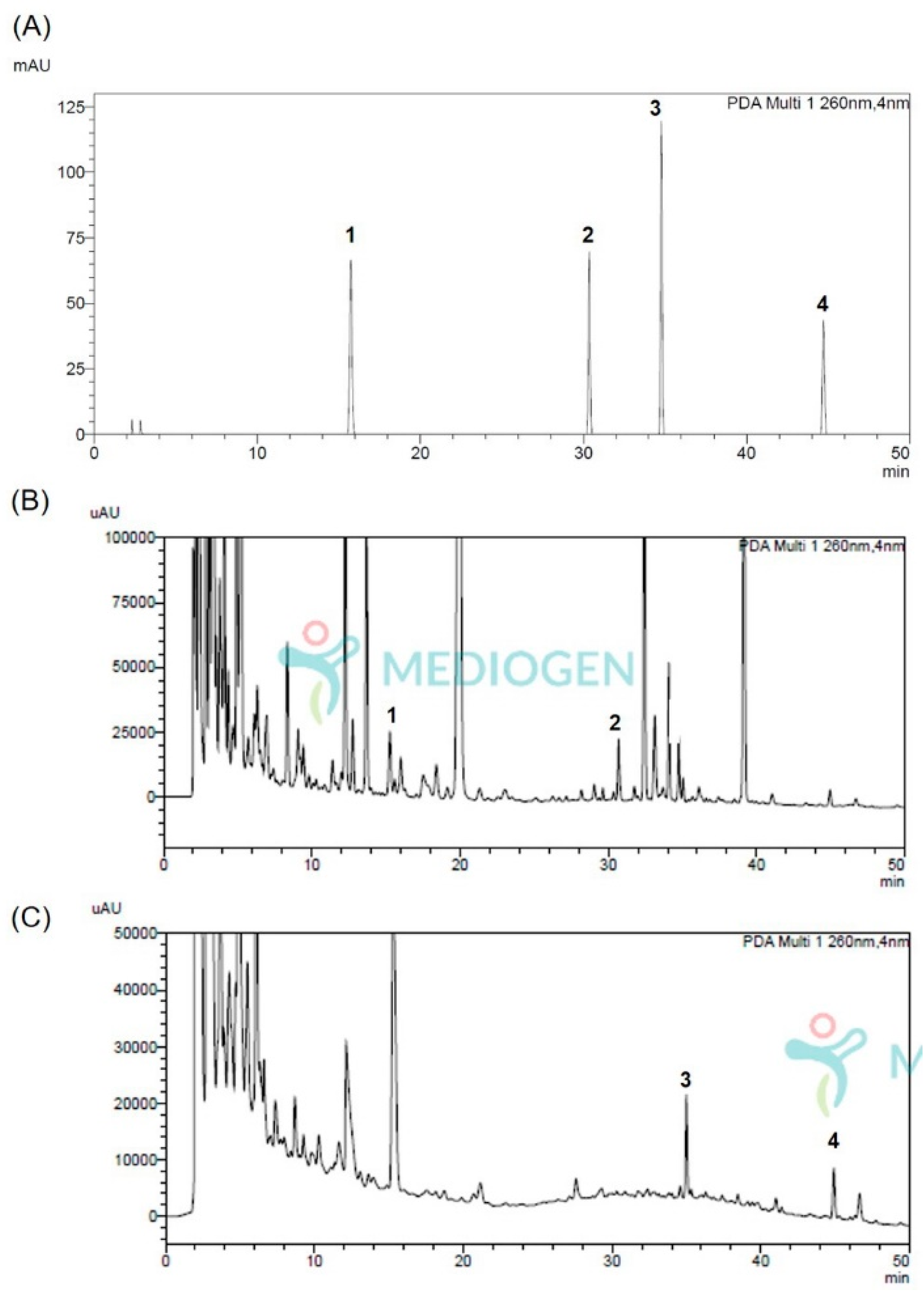
3.1. Analysis Method Vaildation of Isoflavonoids Aglycones and Its Glycosides
3.2. Quantification of Isoflavonoids Aglycones and Its Glycosides Changed through the Fermentation Process of Astragalus membranaceus Extracts
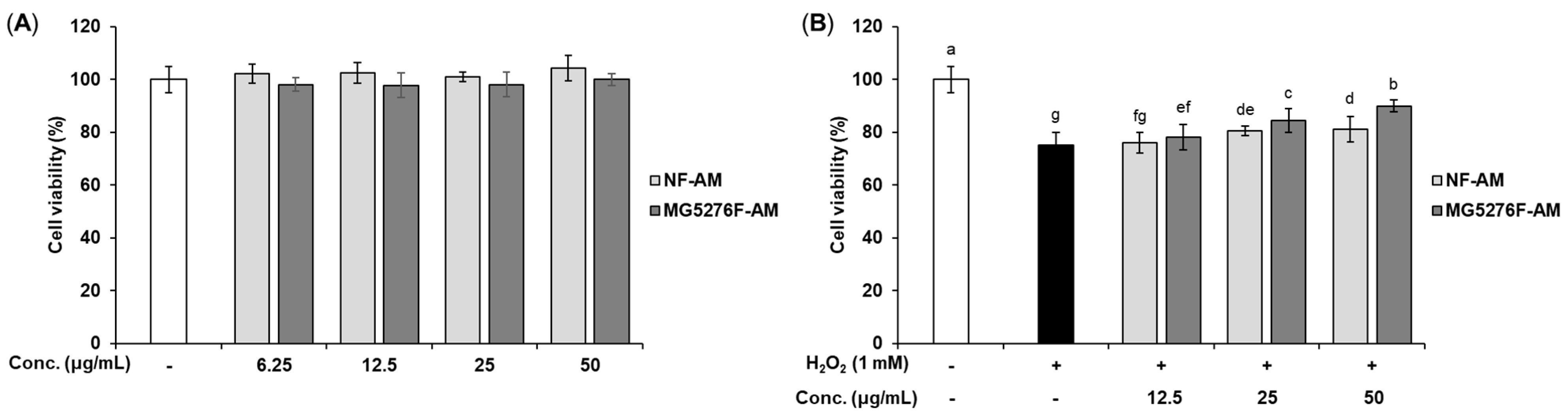
3.3. Cytoprotective Effect of NF-AM and MG5276F-AM in H2O2-Induced HepG2 Cells
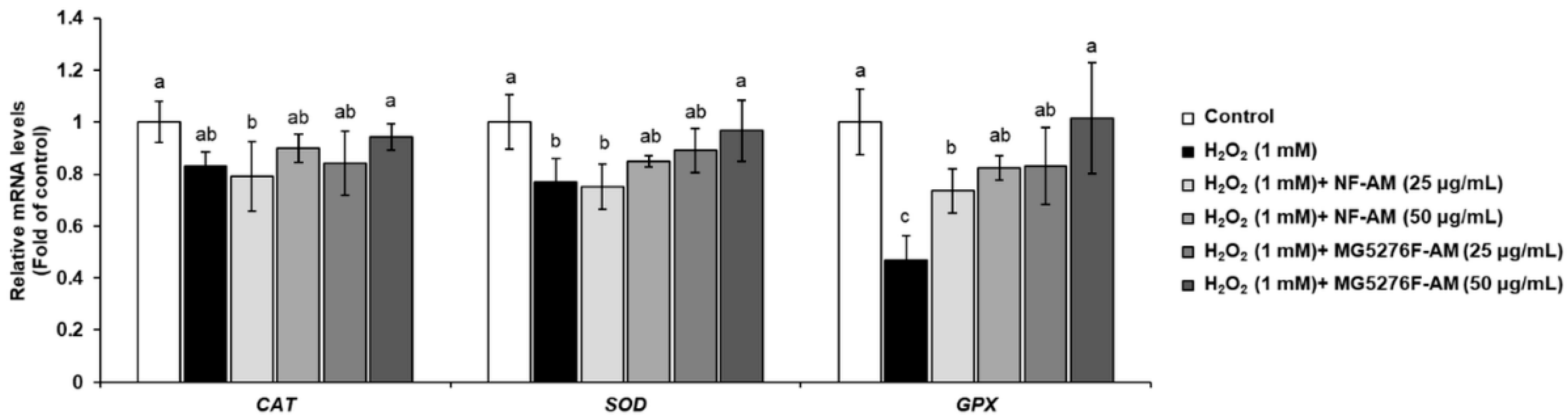
3.4. Effect of NF-AM and MG5276F-AM on Antioxidant Enzymes in H2O2-Induced HepG2 Cells
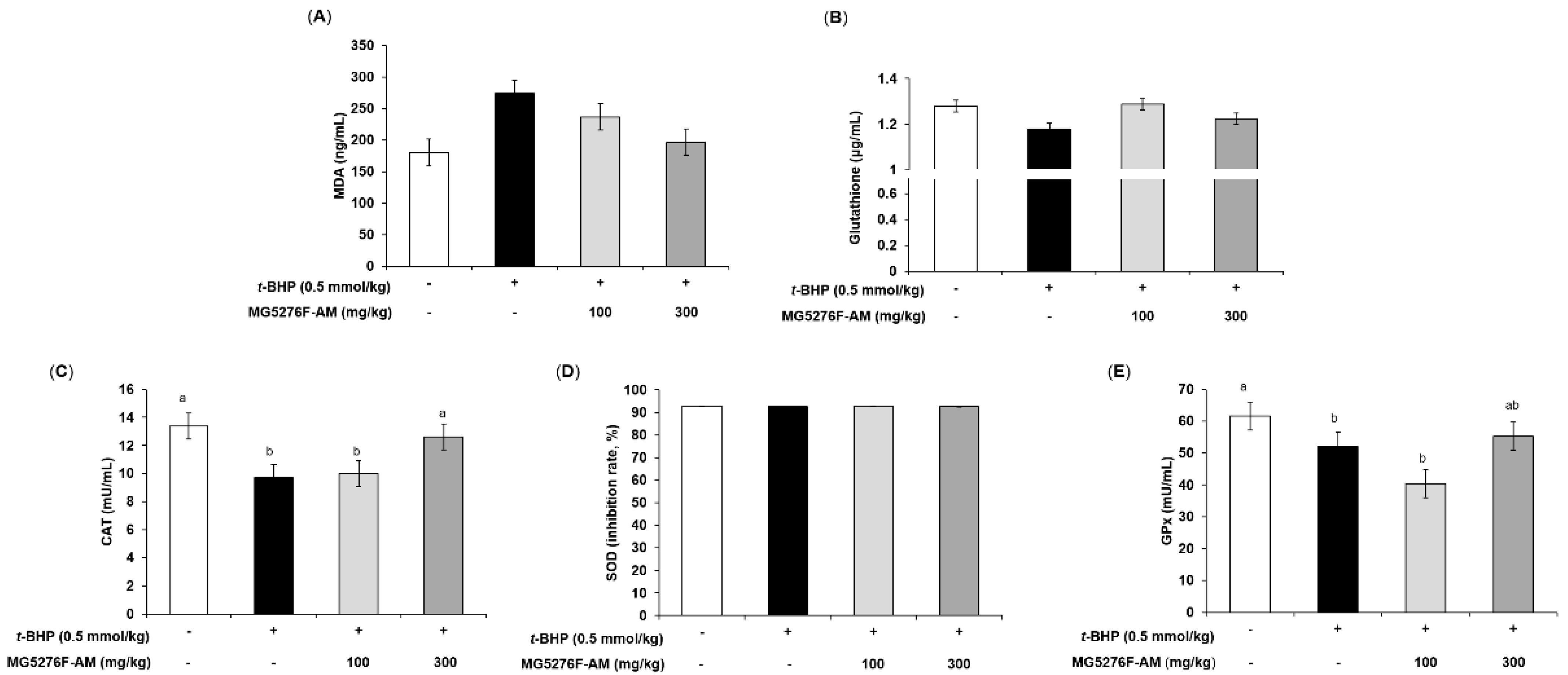
3.5. Effect of NF-AM and MG5276F-AM on AST and ALT in t-BHP-Injected Mice
3.6. Effect of NF-AM and MG5276F-AM on Antioxidant Parameters in t-BHP-Injected Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, I.; Woodside, J. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Yashin, A.; Yashin, Y.; Xia, X.; Nemzer, B. Antioxidant activity of spices and their impact on human health: A review. Antioxidants 2017, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, N.; Santiago, A.; Alías, J.C. Quantification of the antioxidant activity of plant extracts: Analysis of sensitivity and hierarchization based on the method used. Antioxidants 2020, 9, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matés, J.M.; Pérez-Gómez, C.; De Castro, I.N. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Adwas, A.A.; Elsayed, A.; Azab, A.; Quwaydir, F. Oxidative stress and antioxidant mechanisms in human body. J. Appl. Biotechnol. 2019, 6, 43–47. [Google Scholar]
- Kim, J.G.; Dong, X.; Park, S.H.; Bayazid, A.B.; Jeoung, S.A.; Lim, B.O. Bioconversion of black rice and blueberry regulate immunity system through regulation of MAPKs, NF-kB in RAW264. 7 macrophage cells. Food Agr. Immunol. 2021, 32, 471–481. [Google Scholar] [CrossRef]
- Czekała, W. Concept of IN-OIL project based on bioconversion of by-products from food processing industry. J. Ecol. Eng. 2017, 18, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Eom, S.J.; Kim, K.-T.; Paik, H.-D. Microbial bioconversion of ginsenosides in Panax ginseng and their improved bioactivities. Food Rev. Int. 2018, 34, 698–712. [Google Scholar] [CrossRef]
- Lee, N.-K.; Paik, H.-D. Bioconversion using lactic acid bacteria: Ginsenosides, GABA, and phenolic compounds. J. Microbiol. Biotechnol. 2017, 27, 869–877. [Google Scholar] [CrossRef]
- Lee, Y.; Oh, J.; Jeong, Y.-S. Lactobacillus plantarum-mediated conversion of flavonoid glycosides into flavonols, quercetin, and kaempferol in Cudrania tricuspidata leaves. Food Sci. Biotechnol. 2015, 24, 1817–1821. [Google Scholar] [CrossRef]
- Jung, J.; Jang, H.J.; Eom, S.J.; Choi, N.S.; Lee, N.-K.; Paik, H.-D. Fermentation of red ginseng extract by the probiotic Lactobacillus plantarum KCCM 11613P: Ginsenoside conversion and antioxidant effects. J. Ginseng Res. 2019, 43, 20–26. [Google Scholar] [CrossRef]
- Kim, B.-H.; Jang, J.-O.; Lee, J.-H.; Park, Y.; Kim, J.-G.; Yoon, Y.-C.; Jeong, S.J.; Kwon, G.-S.; Lee, J.-B. Increased anti-oxidative activity and whitening effects of a Saposhnikovia extract following bioconversion fermentation using Lactobacillus plantarum BHN-LAB 33. J. Life Sci. 2019, 29, 1208–1217. [Google Scholar]
- Park, J.-Y. Protective Activity of Roasted Astragalus membranaceus against UVB-Induced Damage in Human Dermal Fibroblasts and Artificial Skin Tissues. Master’s Thesis, Chungbuk National University, Cheongju, Korea, 2020. [Google Scholar]
- Li, Y.; Guo, S.; Zhu, Y.; Yan, H.; Qian, D.-W.; Wang, H.-Q.; Yu, J.-Q.; Duan, J.-A. Comparative analysis of twenty-five compounds in different parts of Astragalus membranaceus var. mongholicus and Astragalus membranaceus by UPLC-MS/MS. J. Pharm. Anal. 2019, 9, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Park, H.M.; Lee, J.Y.; Kim, M.Y.; Kang, C.-H.; Hwang, H.S. Anti-oxidative and anti-inflammatory activities of Astragalus membranaceus fermented by Lactiplantibacillus plantarum on LPS-Induced RAW 264.7 cells. Fermentation 2021, 7, 252. [Google Scholar] [CrossRef]
- Xu, X.; Li, F.; Zhang, X.; Li, P.; Zhang, X.; Wu, Z.; Li, D. In vitro synergistic antioxidant activity and identification of antioxidant components from Astragalus membranaceus and Paeonia lactiflora. PLoS ONE 2014, 9, e96780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hu, G.; Lin, H.C.; Hong, S.J.; Deng, Y.H.; Tang, J.Y.; Seto, S.W.; Kwan, Y.W.; Waye, M.M.Y.; Wang, Y.T. Radix astragali extract promotes angiogenesis involving vascular endothelial growth factor receptor-related phosphatidylinositol 3-kinase/Akt-dependent pathway in human endothelial cells. Phytother. Res 2009, 23, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. In Cancer Cell Culture; Humana Press: Totowa, NJ, USA, 2011; pp. 237–245. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, M.; Oh, S.B.; Lee, K.M.; Kim, S.M.; Nho, C.W.; Yoon, W.B.; Kang, K.; Pan, C.H. The protective effect of antioxidant enriched fractions from colored potatoes against hepatotoxic oxidative stress in cultured hepatocytes and mice. J. Food Biochem. 2017, 41, e12315. [Google Scholar] [CrossRef] [Green Version]
- Part 1: Validation of analytical procedures: Text and methodology. In Ich Harmonised Tripartite Guideline; Somatek Inc.: San Diego, CA, USA, 2005; p. 5.
- Toda, S.; Shirataki, Y. Inhibitory effects of isoflavones in roots of Astragalus membranaceus Bunge (Astragali Radix) on lipid peroxidation by reactive oxygen species. Phytother. Res. 1998, 12, 59–61. [Google Scholar] [CrossRef]
- Ávila, M.; Hidalgo, M.; Sánchez-Moreno, C.; Pelaez, C.; Requena, T.; de Pascual-Teresa, S. Bioconversion of anthocyanin glycosides by Bifidobacteria and Lactobacillus. Food Res. Int. 2009, 42, 1453–1461. [Google Scholar] [CrossRef]
- Georgieva, R.; Iliev, I.; Haertlé, T.; Chobert, J.-M.; Ivanova, I.; Danova, S. Technological properties of candidate probiotic Lactobacillus plantarum strains. Int. Dairy J. 2009, 19, 696–702. [Google Scholar] [CrossRef]
- Yuksekdag, Z.; Cinar Acar, B.; Aslim, B.; Tukenmez, U. β-Glucosidase activity and bioconversion of isoflavone glycosides to aglycones by potential probiotic bacteria. Int. J. Food Prop. 2017, 20, S2878–S2886. [Google Scholar] [CrossRef] [Green Version]
- Rekha, C.; Vijayalakshmi, G. Bioconversion of isoflavone glycosides to aglycones, mineral bioavailability and vitamin B complex in fermented soymilk by probiotic bacteria and yeast. J. Appl. Microbiol. 2010, 109, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-H.; Jang, J.-O.; Lee, J.-H.; Park, Y.-E.; Kim, J.-G.; Yoon, Y.-C.; Jeong, S.J.; Kwon, G.-S.; Lee, J.-B. Bioconversion of glycosides isoflavones to aglycone isoflavones by Lactobacillus rhamnosus BHN-LAB 76 under anaerobic conditions. Korean J. Food Preserv. 2019, 26, 148–156. [Google Scholar] [CrossRef]
- Pan, H.; Fang, C.; Zhou, T.; Wang, Q.; Chen, J. Accumulation of calycosin and its 7-O-β-d-glucoside and related gene expression in seedlings of Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao induced by low temperature stress. Plant Cell Rep. 2007, 26, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.-J.; Jeong, W.-S.; Kim, K.-B. Detoxifying effect of fermented black ginseng on H2O2-induced oxidative stress in HepG2 cells. Int. J. Mol. Med. 2014, 34, 1516–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, H.M.; Kim, Y.J.; Oh, E.J.; Oh, S.H.; Choi, J.Y.; Cho, J.H.; Kim, C.D.; Park, S.H.; Kim, Y.L. Hypoxanthine induces cholesterol accumulation and incites atherosclerosis in apolipoprotein E-deficient mice and cells. J. Cell Mol. Med. 2016, 20, 2160–2172. [Google Scholar] [CrossRef]
- Liu, J.; Tan, F.; Liu, X.; Yi, R.; Zhao, X. Grape skin fermentation by Lactobacillus fermentum CQPC04 has anti-oxidative effects on human embryonic kidney cells and apoptosis-promoting effects on human hepatoma cells. RSC Adv. 2020, 10, 4607–4620. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Liu, Z.J.; Chen, T.; Zhao, D. Pharmaceutical properties of calycosin, the major bioactive isoflavonoid in the dry root extract of Radix astragali. Pharm. Biol. 2014, 52, 1217–1222. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhao, S.; Li, F.; Zhang, B.; Qu, Y.; Sun, T.; Luo, T.; Li, D. Investigation of antioxidant interactions between Radix Astragali and Cimicifuga foetida and identification of synergistic antioxidant compounds. PLoS ONE 2014, 9, e87221. [Google Scholar] [CrossRef]
- Sekhar, S.; Sampath-Kumara, K.K.; Niranjana, S.R.; Prakash, H.S. Attenuation of reactive oxygen/nitrogen species with suppression of inducible nitric oxide synthase expression in RAW 264.7 macrophages by bark extract of Buchanania lanzan. Pharmacogn. Mag. 2015, 11, 283. [Google Scholar] [PubMed] [Green Version]
- Oh, J.M.; Jung, Y.S.; Jeon, B.S.; Yoon, B.I.; Lee, K.S.; Kim, B.H.; Oh, S.J.; Kim, S.K. Evaluation of hepatotoxicity and oxidative stress in rats treated with tert-butyl hydroperoxide. Food Chem. Toxicol. 2012, 50, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Martín, C.; Martínez, R.; Navarro, R.; Ruiz-Sanz, J.I.; Lacort, M.; Ruiz-Larrea, M.B. tert-Butyl hydroperoxide-induced lipid signaling in hepatocytes: Involvement of glutathione and free radicals. Biochem. Pharmacol. 2001, 62, 705–712. [Google Scholar] [CrossRef]
- Kim, E.Y.; Hong, K.-B.; Suh, H.J.; Choi, H.-S. Protective effects of germinated and fermented soybean extract against tert-butyl hydroperoxide-induced hepatotoxicity in HepG2 cells and in rats. Food Funct. 2015, 6, 3512–3521. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Primer Sequence (5′→3′) | Product Size (bp) | |
|---|---|---|---|
| CAT | Forward | GAACTGTCCCTACCGTGCTCGA | 156 |
| Reverse | CCAGAATATTGGATGCTGTGCTCCAGG | ||
| SOD | Forward | AATGGACCAGTGAAGGTGTGGGG | 186 |
| Reverse | CACATTGCCCAAGTCTCCAACATGC | ||
| GPX | Forward | CGGCCCAGTCGGTGTATGC | 122 |
| Reverse | CGTGGTGCCTCAGAGGGAC | ||
| GAPDH | Forward | ACCCACTCCTCCACCTTTG | 178 |
| Reverse | CTCTTGTGCTCTTGCTGGG | ||
| Standards 1 | Rt (min) | Calibration Curve 2 | R 2,3 | LOD (μg/mL) | LOQ (μg/mL) |
|---|---|---|---|---|---|
| Calycosin-G | 15.74 | Y = 57,122X – 19,253 | 1.00 | 1.66 | 5.02 |
| Formononetin-G | 30.36 | Y = 46,662X – 9324 | 1.00 | 2.47 | 7.49 |
| Calycosin | 34.76 | Y = 55,047X – 22,903 | 1.00 | 2.93 | 8.87 |
| Formononetin | 44.71 | Y = 52,371X – 22,302 | 1.00 | 3.43 | 10.40 |
| Sample 1 | Content (mg/100 g Dried Weight) | |||
|---|---|---|---|---|
| Calycosin | Formononetin | Calycosin-G | Formononetin-G | |
| NF-AM | n.d. 2 | n.d. | 0.91 ± 0.01 | 0.58 ± 0.00 |
| MG5276F-AM | 0.88 ± 0.02 | 0.87 ± 0.00 | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.Y.; Park, H.M.; Kang, C.-H. Antioxidant Effect via Bioconversion of Isoflavonoid in Astragalus membranaceus Fermented by Lactiplantibacillus plantarum MG5276 In Vitro and In Vivo. Fermentation 2022, 8, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010034
Lee JY, Park HM, Kang C-H. Antioxidant Effect via Bioconversion of Isoflavonoid in Astragalus membranaceus Fermented by Lactiplantibacillus plantarum MG5276 In Vitro and In Vivo. Fermentation. 2022; 8(1):34. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010034
Chicago/Turabian StyleLee, Ji Yeon, Hye Min Park, and Chang-Ho Kang. 2022. "Antioxidant Effect via Bioconversion of Isoflavonoid in Astragalus membranaceus Fermented by Lactiplantibacillus plantarum MG5276 In Vitro and In Vivo" Fermentation 8, no. 1: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010034