
Lipids and Fatty Acids of the White Sea Herring Clupea pallasi marisalbi Berg (Clupeiformes, Clupeidae) from Different Habitats of the White Sea

 ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
3.1. Total Lipids and Their Classes
3.2. Membrane Lipids
3.3. Fatty Acids
4. Materials and Methods
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kuznetsov, V.V. The White Sea and Its Biological Features of Flora and Fauna; Academy of USSR: Moscow, Russia, 1960; pp. 57–298. [Google Scholar]
- Lapin, Y.E. White Sea herring as a Biological Entity. Laws of Fish Population Dynamics of the White Sea and Its Basin; Nauka: Moscow, Russia, 1966; pp. 5–28. [Google Scholar]
- Lapin, Y.E.; Pohilyuk, V.V. Spatial dynamics generations of the White Sea herring Clupea pallasi marisalbi Berg. J. Ichthyol. 1993, 33, 367–371. [Google Scholar]
- Galkina, L.A. The Origin and the Locality of the White Sea Herring. VIII Session of the Scientific Council on the Issue of “Biological Resources of the White Sea and the Inland Waters of the European North” (Abstracts); KarRC RAS: Petrozavodsk, Russia, 1969; pp. 207–208. [Google Scholar]
- Dushkina, L.A. The pigmentation of the larvae of sea herring Clupea in the early stages of development in relation to population dynamics. In Biology of the White Sea Herring; Elsevier Applied Science: London, UK, 1975; pp. 227–254. [Google Scholar]
- Muhomedyarov, F.B. Migration ways of the small White Sea herring based on their labeling. In Biology White Sea Herring; Elsevier Applied Science: London, UK, 1975; pp. 109–125. [Google Scholar]
- Layus, D.L. Population Structure of the White Sea Herring: Data of Karyological Analysis. The Results of Study of the White Sea Herring; ZIN RAS: St. Petersburg, Russia, 1990; pp. 113–125. [Google Scholar]
- Vekhov, V.N. Current status of eelgrass in the White Sea. In The White Sea. Biological Resources and Problems of Their Rational Use. Part 1; ZIN RAS: St. Petersburg, Russia, 1995; pp. 176–187. [Google Scholar]
- Alimov, A.F. Production Hydrobiology and functioning of ecosystems. In New Ideas in Oceanology; Nauka: Moscow, Russia, 2004; Volume 1, pp. 264–279. [Google Scholar]
- Shatunovskiy, M.I. Ecological Features of Metabolism of Marine Fish; Nauka: Moscow, Russia, 1980; p. 288. [Google Scholar]
- Gershanowitsch, A.D.; Lapin, V.I.; Shatunovskij, M.I. Specific features of lipid metabolism in fish. Usp. Sovrem. Biol. 1991, 3, 207–219. [Google Scholar]
- Pavlov, D.S.; Nefedova, Z.A.; Veselov, A.E.; Nemova, N.N.; Ruokolainen, T.R.; Vasileva, O.B.; Ripatti, P.O. Age dynamics of lipid status of juveniles of Atlantic salmon (Salmo salar L.) from the Varzuga river. J. Ichthyol. 2009, 49, 1073–1080. [Google Scholar] [CrossRef]
- Sidorov, V.S. Ecological Biochemistry of Fishes; Nauka: Leningrad, Russia, 1983; p. 240. [Google Scholar]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: New York, NY, USA, 2002; p. 466. [Google Scholar]
- Iverson, S.J. Tracing aquatic food webs using fatty acids: From qualitative indicators to quantitative determination. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: New York, NY, USA, 2009; pp. 281–309. [Google Scholar]
- Iljash, L.B.; Zitina, L.S.; Fedorov, V.D. Phytoplankton of the White Sea; Janus-K: Moscow, Russia, 2003; pp. 48–154. [Google Scholar]
- Primakov, I.M. Morphometry and oceanographic characteristics. In Biological Resources of the White Sea: Study and Use. The Fauna of the Seas; ZIN RAS: St. Petersburg, Russia, 2012; Volume 69, pp. 7–13. [Google Scholar]
- Sapozhnikov, V.V.; Arzhanova, N.V.; Mordasova, N.V. Hydrochemical bases productivity. In Biological Resources of the White Sea: Study and Use. The Fauna of the Seas; ZIN RAS: St. Petersburg, Russia, 2012; Volume 69, pp. 13–34. [Google Scholar]
- Troshkov, V.A. Zooplankton. In Biological Resources of the White Sea: Study and Use. The Fauna of the Seas; ZIN RAS: St. Petersburg, Russia, 2012; Volume 69, pp. 67–87. [Google Scholar]
- Karagezyan, K. The Phospholipids and Their Role in the Life of the Organism; Yerevan State University: Yerevan, Armenia, 1977; pp. 25–29. [Google Scholar]
- Nefedova, Z.A.; Ruokolainen, T.R.; Vasilyev, O.B.; Nemova, N.N.; Kashulin, N.A. Some lipid metabolism of different age females of Coregonus albula from the lake. Syamozero (Karelia) and Lake. Kovdor (Murmansk region) during feeding. J. Ichthyol. 2005, 45, 689–694. [Google Scholar]
- Murzina, S.A. The Role of Lipids and Their Fatty Acid Components in the Biochemical Adaptations of the Daubed Shanny Leptoclinus maculatus F. from the North-Western Coastal Waters of Spitsbergen. Ph.D. Thesis, Karelian State Pedagogical Academy, Petrozavodsk, Russia, 2010; p. 184. [Google Scholar]
- Dyatlovitskaya, E.V.; Bezuglov, V.V. Lipids like bioeffectors. Introduction. Biochemistry 1998, 63, 67–74. [Google Scholar]
- Kolomiytseva, I.K.; Perepelkina, N.I.; Patrushev, I.V.; Popov, V.I. The role of lipids in the endoplasmic reticulum and assembly dictyosomes of the neuronal cells of the cells of the cerebral cortex Yakut gopher Citellus undulates during hibernation. Biochemistry 2003, 68, 954–967. [Google Scholar]
- Christie, W.W. Available online: http://lipidlibrary.aocs.org/Primer/content.cfm?ItemNumber=39292&navItemNumber=19204 (accessed on 15 May 2016).
- Lopuhin, Y.M.; Archakov, A.I.; Vladimirov, Y.A.; Kogan, E.M. Cholesterinoz; Medicine: Moscow, Russia, 1985; p. 350. [Google Scholar]
- Nemova, N.N.; Nefedova, Z.A.; Murzina, S.A.; Veselov, A.E.; Ripatti, P.O.; Pavlov, D.S. The effect of environmental conditions on the dynamics of fatty acids in juveniles of the Atlantic salmon (Salmo salar L.). Russ. J. Ecol. 2015, 46, 267–271. [Google Scholar] [CrossRef]
- Yanes-Roca, C.; Rhody, N.; Nystrom, M.; Main, K.L. Effects of fatty acid composition and spawning season patterns on egg quality and larval survival in common snook (Centropomus undecimalis). Aquaculture 2009, 287, 335–340. [Google Scholar] [CrossRef]
- Kreps, E.M. Lipids of Cell Membrane. Evolution of Brain Lipids; Nauka: Leningrad, Russia, 1981; p. 339. [Google Scholar]
- Viso, A.C.; Marty, J.C. Fatty acids from 28 marine microalgae. Phytochemistry 1993, 34, 1521–1533. [Google Scholar] [CrossRef]
- Viron, C.; Saunois, A.; Andre, P.; Perly, B.; Laffose, M. Isolation and identification of unsaturated fatty acid methyl esters from marinew micro-algae. Anal. Chim. Acta 2000, 409, 257–266. [Google Scholar] [CrossRef]
- Graeve, M.; Lundberg, M.; Boer, M.; Kattner, G.; Hop, H.; Falk-Petersen, S. The fate of dietary lipids in the Arctic ctenophore Mertensia ovum (Fabricius 1780). Mar. Biol. 2008, 153, 643–651. [Google Scholar] [CrossRef] [Green Version]
- Berger, V.J. Production Potential of the White Sea; ZIN RAS: St. Petersburg, Russia, 2007; pp. 66–131. [Google Scholar]
- Falk-Petersen, S.; Hopkins, C.C.E.; Sargent, J.R. Trophic relationships in the pelagic, arctic food web. In Proceedings of the 24th European Marine Biology Symposium, Trondheim, Norway, 12–16 May 1990; pp. 315–333.
- Primakov, I.M.; Berger, V.Y. Production of planktonic crustaceans in the White Sea. Mar. Biol. 2007, 33, 356–360. [Google Scholar] [CrossRef]
- Primakov, I.M.; Kuznetsov, E.A. The study, sustainable use and protection of natural resources of the White Sea. Articles XI All-Russian conference with international participation. In Spatial Distribution of Summer Zooplankton of the White Sea and South-Eastern Part of the Barents Sea; ZIN RAS: St. Petersburg, Russia, 2010; pp. 142–143. [Google Scholar]
- Kalchenko, E.I.; Videnskaya, T.L.; Yurjeva, M.I. Feeding and fatty acid content of wild and captured juveniles of dog-salmon during keeping in river fishponds. In Proceedings of the International Conference Modern Problems of Physiology and Biochemistry of Aquatic Organisms, Petrozavodsk, Russia, 6–9 September 2004; KarRC RAS: Petrozavodsk, Russia, 2004; p. 58. [Google Scholar]
- Sergeeva, M.G.; Varfolomeeva, A.T. The Cascade of Arachidonic Acid; Narodnoe Obrazovanie: Miscow, Russia, 2006; pp. 21–38. [Google Scholar]
- Boldyrev, A.A.; Kaivarainen, E.I.; Ilykha, V.A. Biomembranology; KarRC RAS: Petrozavodsk, Russia, 2006; pp. 24–102. [Google Scholar]
- Semushin, A.V.; Sherstkov, V.S.; Rukhlova, V.A. Diversity of fish species in the trawl catches in the southeastern Barents Sea in 1980–2008. J. Ichthyol. 2011, 51, 717–737. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissue. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Walsh, D.E.; Banasik, O.J.; Gilles, K.A. Thin-layer chromatographic separation and colorimetric analysis of barley or malt lipid classes and their fatty acids. J. Chromatogr. 1965, 17, 278–287. [Google Scholar] [CrossRef]
- Sidorov, V.S.; Lisenko, E.I.; Bolgova, O.M.; Nefedova, Z.A. Lipids of Fish. I. Methods of Analyses. The Tissues Specificity of Vebdace Lipids (Coregonus albula L.). In Salmonidae of Karelia. Issue 1 Ecology Parasites Biochemistry; Potapova, O.I., Smirnov, Y.A., Eds.; Karelian Branch of the USSR Academy of Sciences, Institute of Biology: Petrozavodsk, Russia, 1972; pp. 150–161. [Google Scholar]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol determination in serum. A rapid direction method. S. Afr. Med. J. 1974, 48, 250–356. [Google Scholar] [PubMed]
- Arduini, A. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies. J. Lipid Res. 1996, 37, 684–689. [Google Scholar] [PubMed]
- Jamieson, G.R. GLC-identification techniques for longchain unsaturated fatty acids. J. Chromatogr. Sci. 1975, 13, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Korosov, A.V.; Gorbach, V.V. Computer Processing of Biological Data; KarRC RAS: Petrozavodsk, Russia, 2007; p. 76. [Google Scholar]
- Zabetakis, I. Marine Oils and Diseases in Marine Oils (from Sea to Pharmaceuticals); NOVA Science Publishers: New, York, NY, USA, 2015; pp. 83–90. [Google Scholar]
- Morphis, G.; Kyriazopoulou, A.; Nasopoulou, C.; Sioriki, E.; Demopoulos, C.A.; Zabetakis, I. Assessment of the in Vitro antithrombotic properties of sardine (Sardina pilchardus) fillet lipids and cod liver oil. Fishes 2016, 1, 1–15. [Google Scholar] [CrossRef]



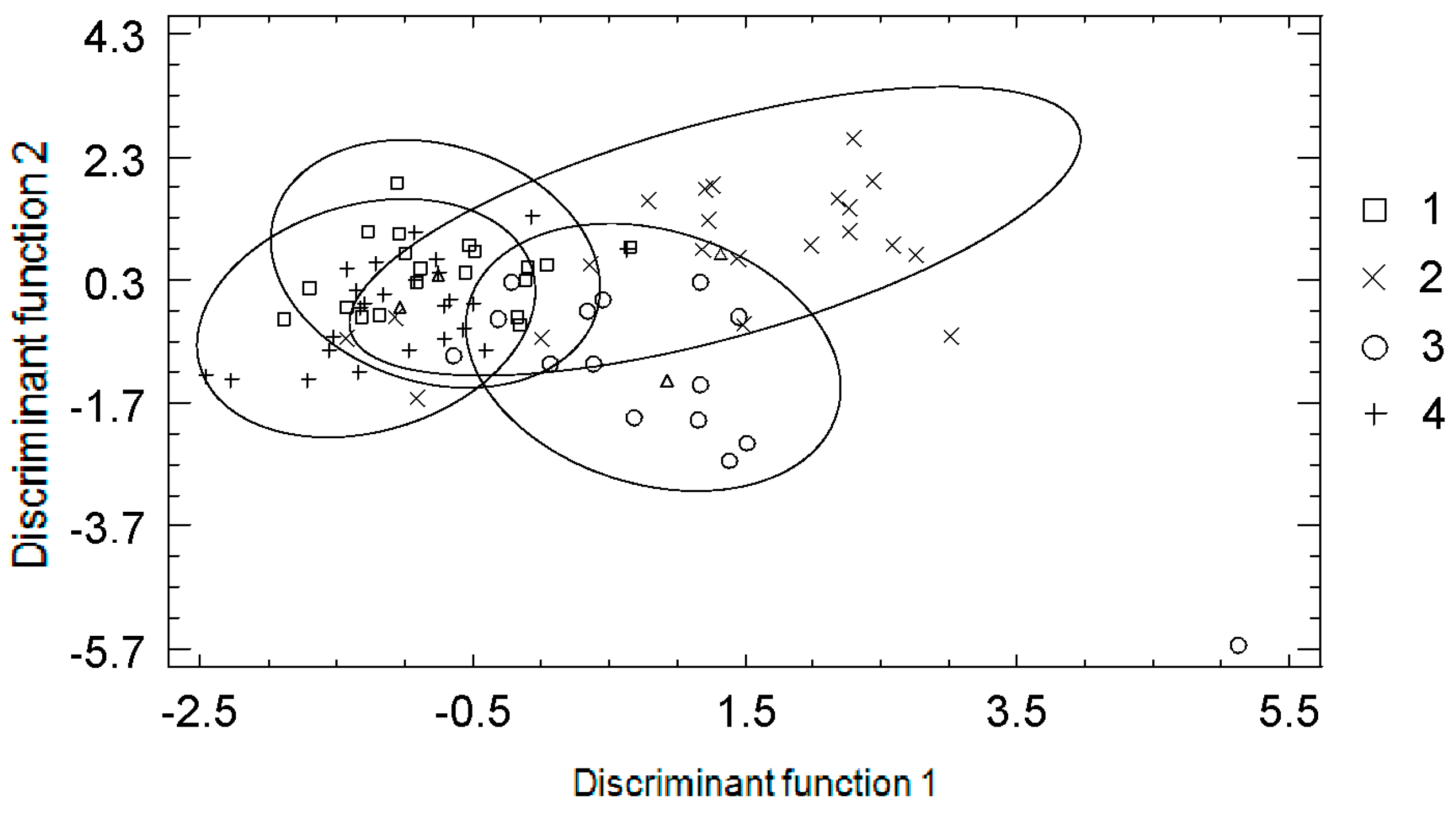{kind=link}
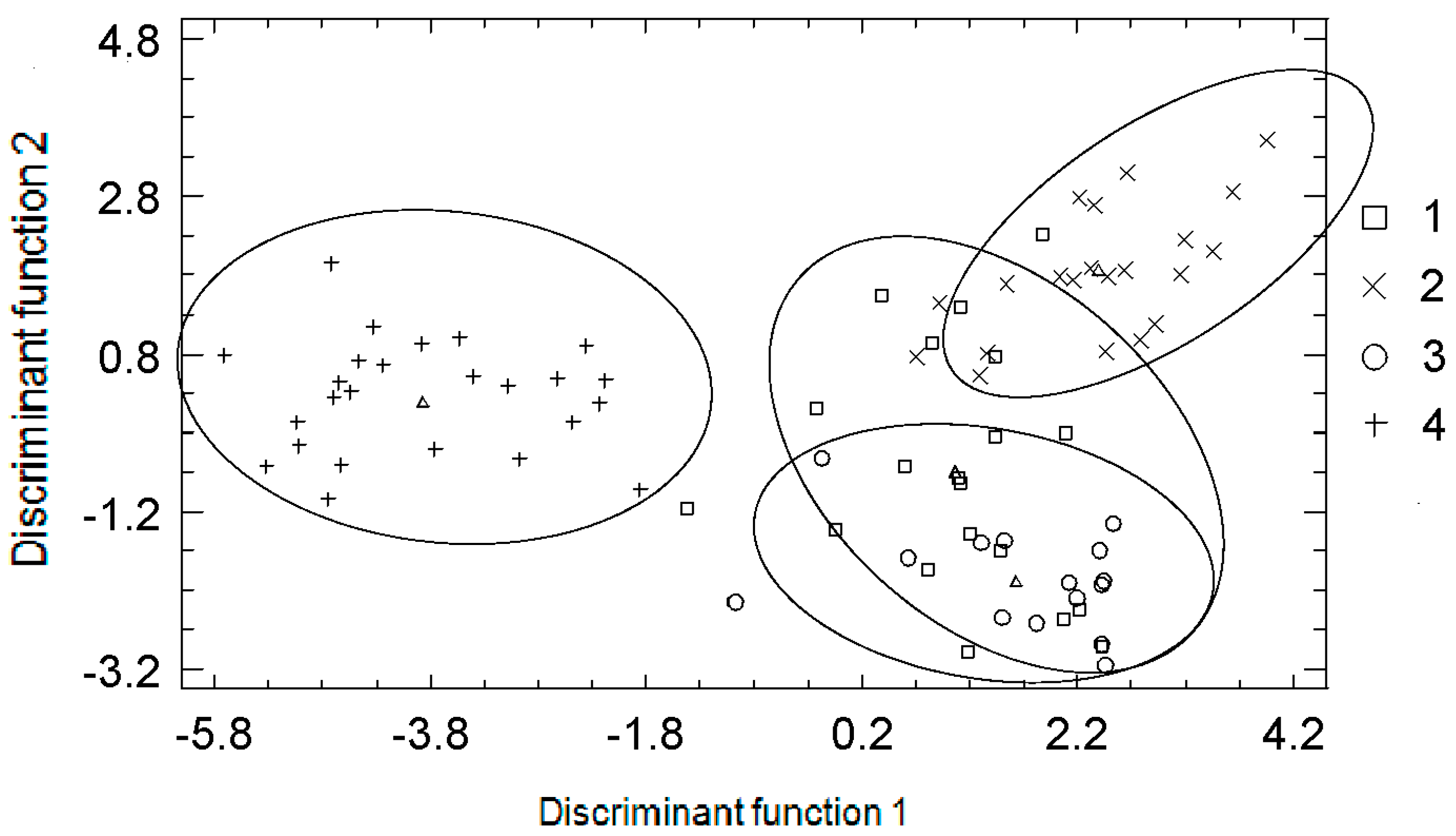{kind=link}
| Sampling Place | Dvina Bay | Kandalaksha Bay | Tersky Shore | Onega Bay | MANOVA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | |||||||||||
| Number of fish | 20 | 21 | 15 | 25 | p | % | F | ||||
| Fish length, cm | 11.9 ± 0.17 2,3,4 (10.5–13.0) | 12.4 ± 0.13 1,3,4 (11.0–13.5) | 11.2 ± 0.22 1,2,4 (9.0–12.5) | 9.98 ± 0.39 1,2,3 (8.0–15.0) | |||||||
| % dry weight | |||||||||||
| TL | 34.5 ± 0.6 2,4 | 41.7 ± 1.4 1,3,4 | 34.7 ± 0.8 2,4 | 31.8 ± 0.8 1,2,3 | 0.7623 | 0.0001 | 12.1 | 19.5 | 0.75 | 8.50 | |
| PLs | 19.1 ± 1.3 4 | 18.4 ± 1.0 4 | 17.2 ± 0.8 4 | 14.3 ± 0.7 1,2,3 | 0.5593 | 0.1608 | 21.4 | 5.9 | 0.93 | 1.78 | |
| TAGs | 14.1 ± 1.2 2 | 19.4 ± 0.9 1,3,4 | 12.2 ± 1.0 2,4 | 15.5 ± 0.9 2,3 | 0.3771 | 0.0076 | 22.0 | 12.6 | 1.10 | 4.39 | |
| EfCHOLs | 0.4 ± 0.1 2,3 | 0.7 ± 0.1 1 | 0.8 ± 0.1 1 | 0.5 ± 0.1 | 0.3845 | 0.0301 | 26.1 | 10.9 | 1.09 | 3.20 | |
| CHOL | 0.9 ± 0.1 2,3,4 | 3.2 ± 0.3 1,3,4 | 4.5 ± 0.9 1,4 | 1.5 ± 0.1 1,2,3 | 0.9976 | 0.0000 | 6.3 | 30.1 | 0.31 | 10.57 | |
| CHOL/PLs | 0.05 ± 0.01 2,3,4 | 0.19 ± 0.03 1,4 | 0.29 ± 0.08 1,4 | 0.11 ± 0.01 1,2,3 | - | - | - | - | - | - | |
| Sampling Place/Parameter | Dvina Bay | Kandalaksha Bay | Tersky Shore | Onega Bay |
|---|---|---|---|---|
| Number of Fish | 20 | 21 | 15 | 25 |
| PLs | 19.1 ± 1.3 4 | 18.4 ± 1.0 4 | 17.2 ± 0.8 4 | 14.3 ± 0.7 1,2,3 |
| PI | 0.52 ± 0.08 | 0.68 ± 0.05 3,4 | 0.38 ± 0.04 2 | 0.43 ± 0.03 2 |
| PS | 0.80 ± 0.15 | 0.79 ± 0.10 4 | 0.64 ± 0.07 | 0.54 ± 0.04 2 |
| PEA | 0.59 ± 0.11 | 0.82 ± 0.07 | 0.85 ± 0.10 | 0.62 ± 0.12 |
| PC | 16.06 ± 1.07 2,3,4 | 13.39 ± 0.78 1,4 | 12.93 ± 0.84 1,4 | 10.11 ± 0.55 1,2,3 |
| LysoPC | 0.55 ± 0.12 2,3,4 | 1.44 ± 0.13 1,3 | 0.25 ± 0.02 1,2,4 | 1.28 ± 0.14 1,3 |
| SFM | 0.15 ± 0.05 2,3,4 | 0.54 ± 0.08 1,4 | 0.95 ± 0.23 1 | 0.95 ± 0.12 1,2 |
| Unidentified | 0.39 ± 0.10 3 | 0.75 ± 0.18 4 | 1.20 ± 0.25 1,4 | 0.41 ± 0.06 2,3 |
| Fatty Acids/Sampling Place | Dvina Bay | Kandalaksha Bay | Tersky Shore | Onega Bay |
|---|---|---|---|---|
| Number of Fish | 20 | 21 | 15 | 25 |
| 14:0 | 10.53 ± 0.35 2,4 | 12.40 ± 0.28 1,3,4 | 11.01 ± 0.20 2,4 | 8.80 ± 0.43 1,2,3 |
| 16:0 | 21.66 ± 0.47 3 | 21.40 ± 0.35 3 | 20.43 ± 0.23 1,2,4 | 22.26 ± 0.62 3 |
| 17:0 | 0.47 ± 0.06 3,4 | 0.37 ± 0.06 4 | 0.31 ± 0.01 1,4 | 0.90 ± 0.02 1,2,3 |
| 18:0 | 2.25 ± 0.06 2,4 | 1.66 ± 0.07 1,3,4 | 2.10 ± 0.01 2,4 | 3.21 ± 0.17 1,2,3 |
| ƩSFA | 37.12 ± 0.80 | 37.23 ± 0.66 3 | 35.23 ± 0.33 2 | 37.26 ± 0.81 |
| 16:1(n-9) | 0.73 ± 0.06 | 0.40 ± 0.04 3,4 | 0.68 ± 0.02 2 | 0.74 ± 0.10 2 |
| 16:1(n-7) | 11.31 ± 0.29 4 | 11.45 ± 0.22 4 | 11.11 ± 0.23 4 | 7.73 ± 0.24 1,2,3 |
| 17:1(n-7) | 0.35 ± 0.04 4 | 0.34 ± 0.04 4 | 0.40 ± 0.03 4 | 0.59 ± 0.03 1,2,3 |
| 18:1(n-9) | 19.80 ± 0.35 | 20.14 ± 0.33 3,4 | 18.86 ± 0.25 2 | 18.40 ± 0.73 2 |
| 18:1(n-7) | 4.05 ± 0.28 2,4 | 3.19 ± 0.26 1 | 3.62 ± 0.06 | 3.39 ± 0.10 1 |
| 18:1(n-5) | 0.82 ± 0.05 2,3,4 | 1.09 ± 0.05 1,4 | 1.07 ± 0.05 1,4 | 0.69 ± 0.03 1,2,3 |
| 20:1(n-11) | 0.24 ± 0.05 | 0.17 ± 0.02 | 0.26 ± 0.06 | 0.58 ± 0.24 |
| 20:1(n-9) | 1.46 ± 0.12 2,3 | 2.10 ± 0.21 1 | 2.50 ± 0.22 1 | 1.93 ± 0.56 |
| 22:1(n-9) | 0.32 ± 0.02 | 0.32 ± 0.02 | 0.37 ± 0.02 | 0.79 ± 0.42 |
| 22:1(n-7) | 0.23 ± 0.01 3,4 | 0.21 ± 0.01 4 | 0.20 ± 0.01 1,4 | 0.43 ± 0.03 1,2,3 |
| 24:1(n-9) | 0.22 ± 0.08 4 | 0.27 ± 0.08 3,4 | 0.04 ± 0.00 2,4 | 1.18 ± 0.04 1,2,3 |
| ƩMUFA | 41.40 ± 0.75 | 42.06 ± 0.51 | 41.77 ± 0.49 | 38.81 ± 1.57 |
| 16:3(n-6) | 0.35 ± 0.03 2,4 | 0.21 ± 0.03 1,3,4 | 0.37 ± 0.01 2,4 | 0.01 ± 0.00 1,2,3 |
| 18:2(n-6) | 1.31 ± 0.05 3,4 | 1.21 ± 0.02 3,4 | 1.50 ± 0.06 1,2,4 | 1.06 ± 0.05 1,2,3 |
| 18:3(n-6) | 0.15 ± 0.01 2,3 | 0.11 ± 0.01 1,3,4 | 0.09 ± 0.01 1,2,4 | 0.15 ± 0.01 2,3 |
| 20:4(n-6) | 0.28 ± 0.02 3 | 0.30 ± 0.01 3 | 0.37 ± 0.01 1,2,4 | 0.29 ± 0.02 3 |
| Ʃ(n-6) PUFA | 3.52 ± 0.16 2,4 | 2.70 ± 0.07 1,3 | 3.50 ± 0.11 2,4 | 2.58 ± 0.09 1,3 |
| 18:3(n-3) | 0.76 ± 0.06 2 | 0.92 ± 0.03 1,3 | 0.74 ± 0.03 2,4 | 0.93 ± 0.07 3 |
| 20:5(n-3) | 6.51 ± 0.52 | 6.61 ± 0.31 | 6.78 ± 0.38 | 7.38 ± 0.47 |
| 22:6(n-3) | 5.25 ± 0.45 3,4 | 5.11 ± 0.27 3,4 | 6.53 ± 0.18 1,2 | 7.01 ± 0.65 1,2 |
| Ʃ(n-3) PUFA | 15.12 ± 1.15 4 | 15.84 ± 0.68 4 | 17.06 ± 0.49 | 19.19 ± 1.35 1,2 |
| ƩPUFA | 21.48 ± 1.38 | 20.71 ± 0.74 3 | 23.0 ± 0.50 2 | 23.92 ± 1.48 |
| (n-3)/(n-6) | 4.29 ± 0.02 2,4 | 5.86 ± 0.01 1,3,4 | 4.87 ± 0.01 2,4 | 7.44 ± 0.01 1,2,3 |
| 18:3(n-3)/18:2(n-6) | 0.58 ± 0.04 2,4 | 0.76 ± 0.02 1,3,4 | 0.50 ± 0.02 2,4 | 0.89 ± 0.05 1,2,3 |
| 16:0/18:1(n-9) | 1.10 ± 0.31 4 | 1.07 ± 0.27 4 | 1.08 ± 0.32 4 | 1.24 ± 0.05 1,2,3 |
| Characteristic/Sampling | Dvina Bay | Kandalaksha Bay | Tarsky Shore | Onega Bay |
|---|---|---|---|---|
| Coordinates | 64°57′ 38°23′ | 67°02′ 32°23′ | 66°10′ 37°03′ | 64°59′ 36°37′ |
| Depth, m | 50.0 | 25.0 | 38.0 | 38.0 |
| Water temperature, °С | 6.5 | 2.9 | 5.9 | 6.7 |
| Salinity, ‰ | 26.5 | 27.1 | 27.0 | 26.4 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murzina, S.A.; Nefedova, Z.A.; Pekkoeva, S.N.; Ruokolainen, T.R.; Ripatti, P.O.; Semushin, A.V.; Nemova, N.N. Lipids and Fatty Acids of the White Sea Herring Clupea pallasi marisalbi Berg (Clupeiformes, Clupeidae) from Different Habitats of the White Sea. Fishes 2016, 1, 65-76. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010065
Murzina SA, Nefedova ZA, Pekkoeva SN, Ruokolainen TR, Ripatti PO, Semushin AV, Nemova NN. Lipids and Fatty Acids of the White Sea Herring Clupea pallasi marisalbi Berg (Clupeiformes, Clupeidae) from Different Habitats of the White Sea. Fishes. 2016; 1(1):65-76. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010065
Chicago/Turabian StyleMurzina, Svetlana A., Zinaida A. Nefedova, Svetlana N. Pekkoeva, Tatiana R. Ruokolainen, Pauli O. Ripatti, Andrey V. Semushin, and Nina N. Nemova. 2016. "Lipids and Fatty Acids of the White Sea Herring Clupea pallasi marisalbi Berg (Clupeiformes, Clupeidae) from Different Habitats of the White Sea" Fishes 1, no. 1: 65-76. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010065