Modulation of the Expression of Immune-related Gene in Atlantic and Coho Salmon during Infestation with the Sea lice Caligus rogercresseyi

 , ,
, ,
Abstract
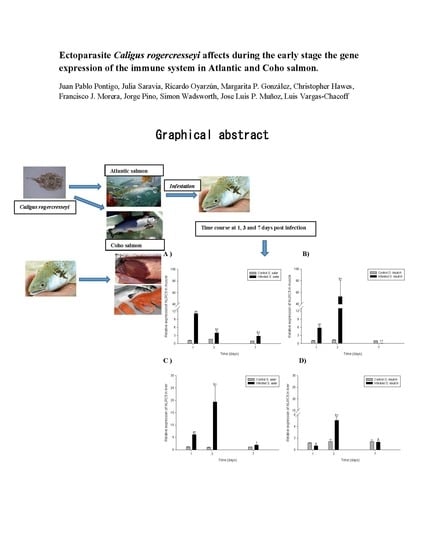
:
1. Introduction
2. Results
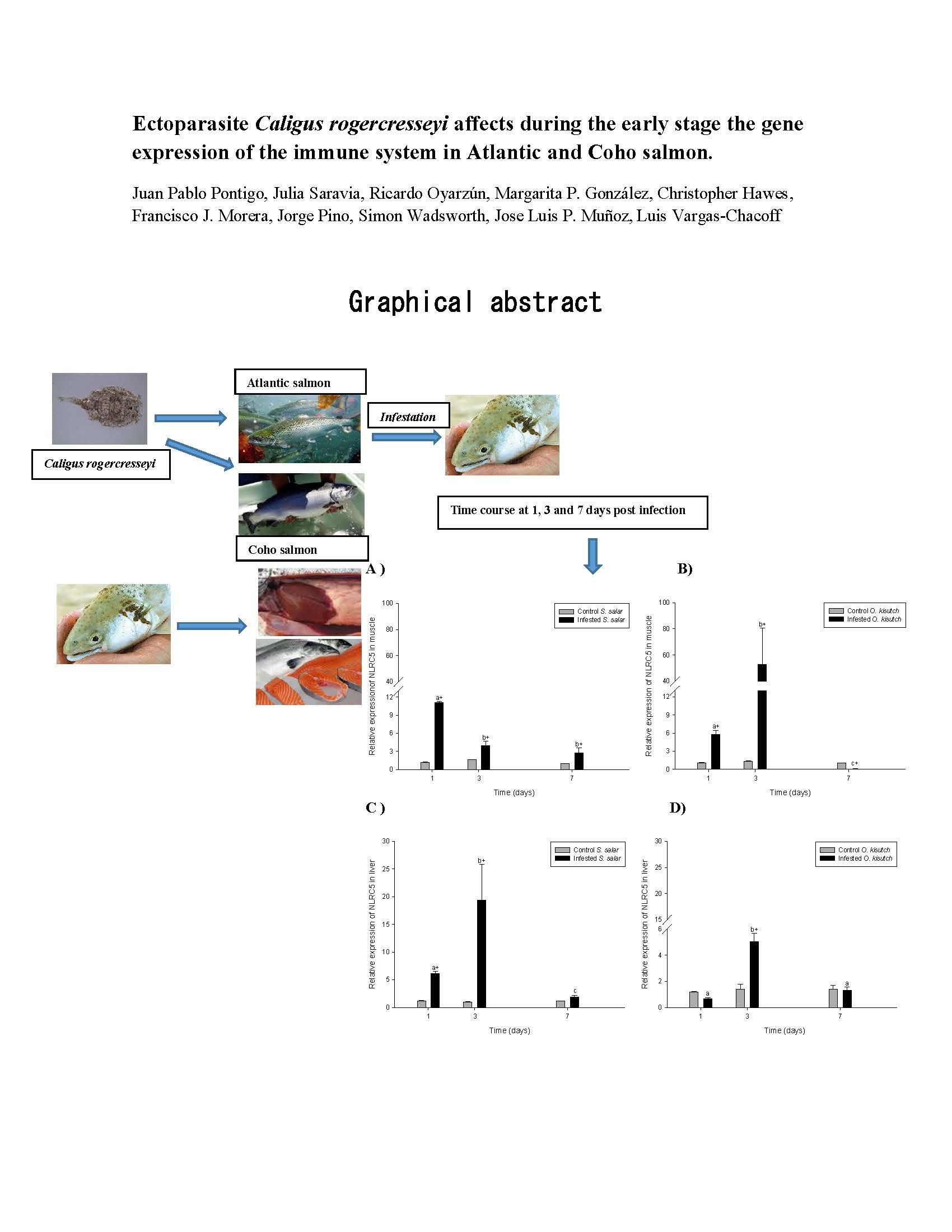
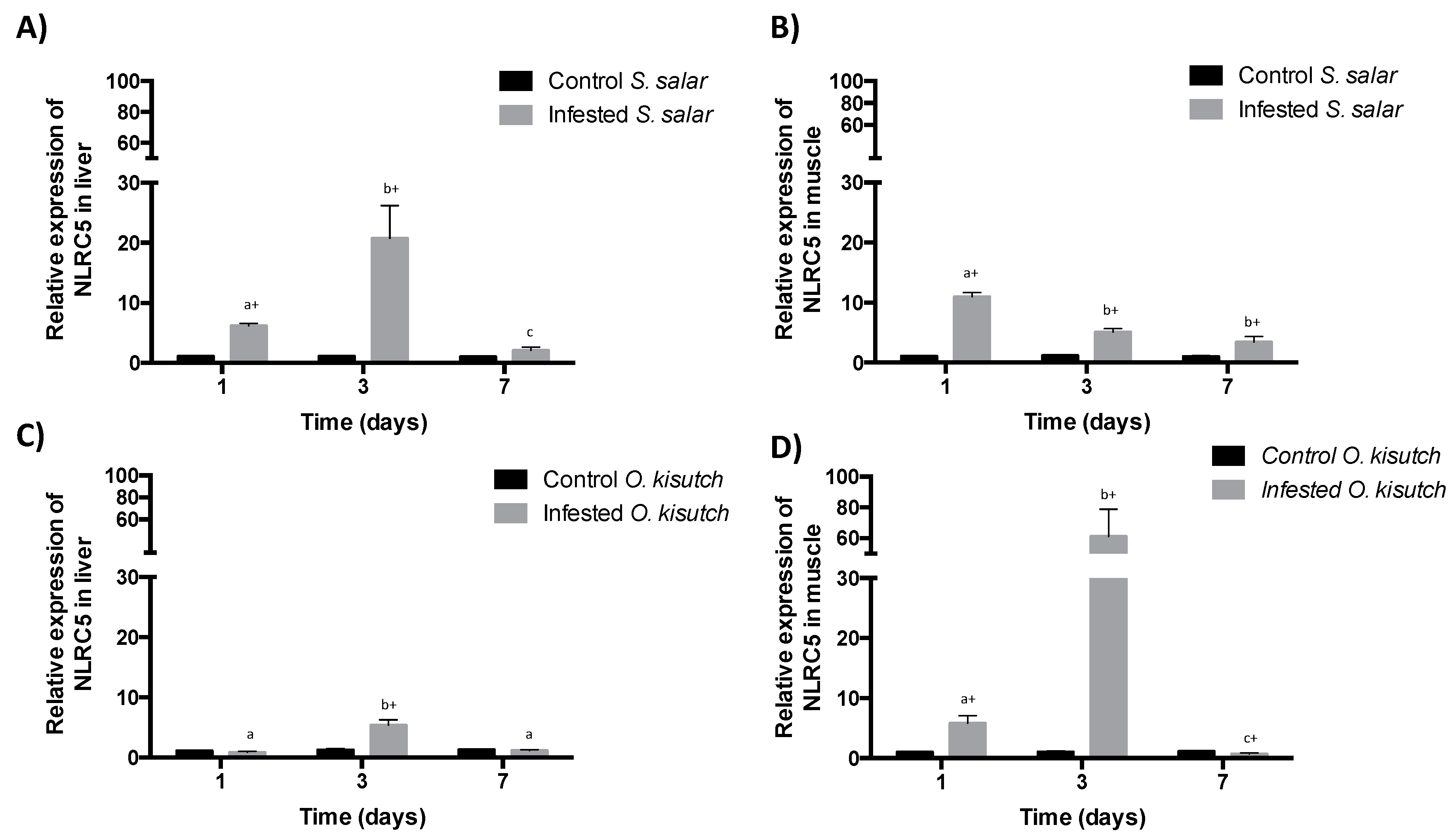
2.1. Analysis of NLRC5 Transcript Expression in the Muscle and Liver of Atlantic and Coho Salmon Infested with C. rogercresseyi
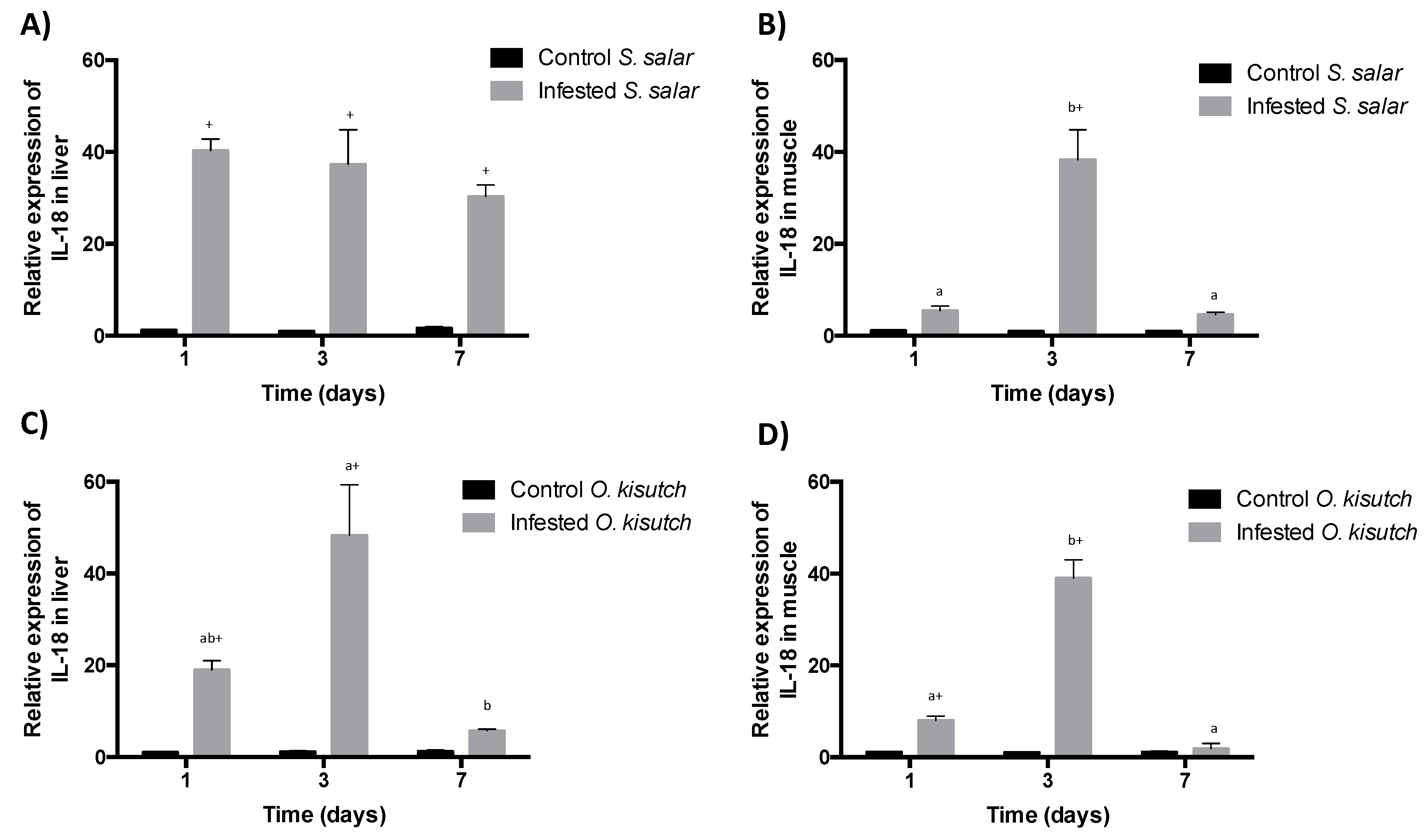
2.2. Analysis of IL-18 Transcript Expression in the Muscle and Liver of Atlantic and Coho Salmon Infested with C. rogercresseyi
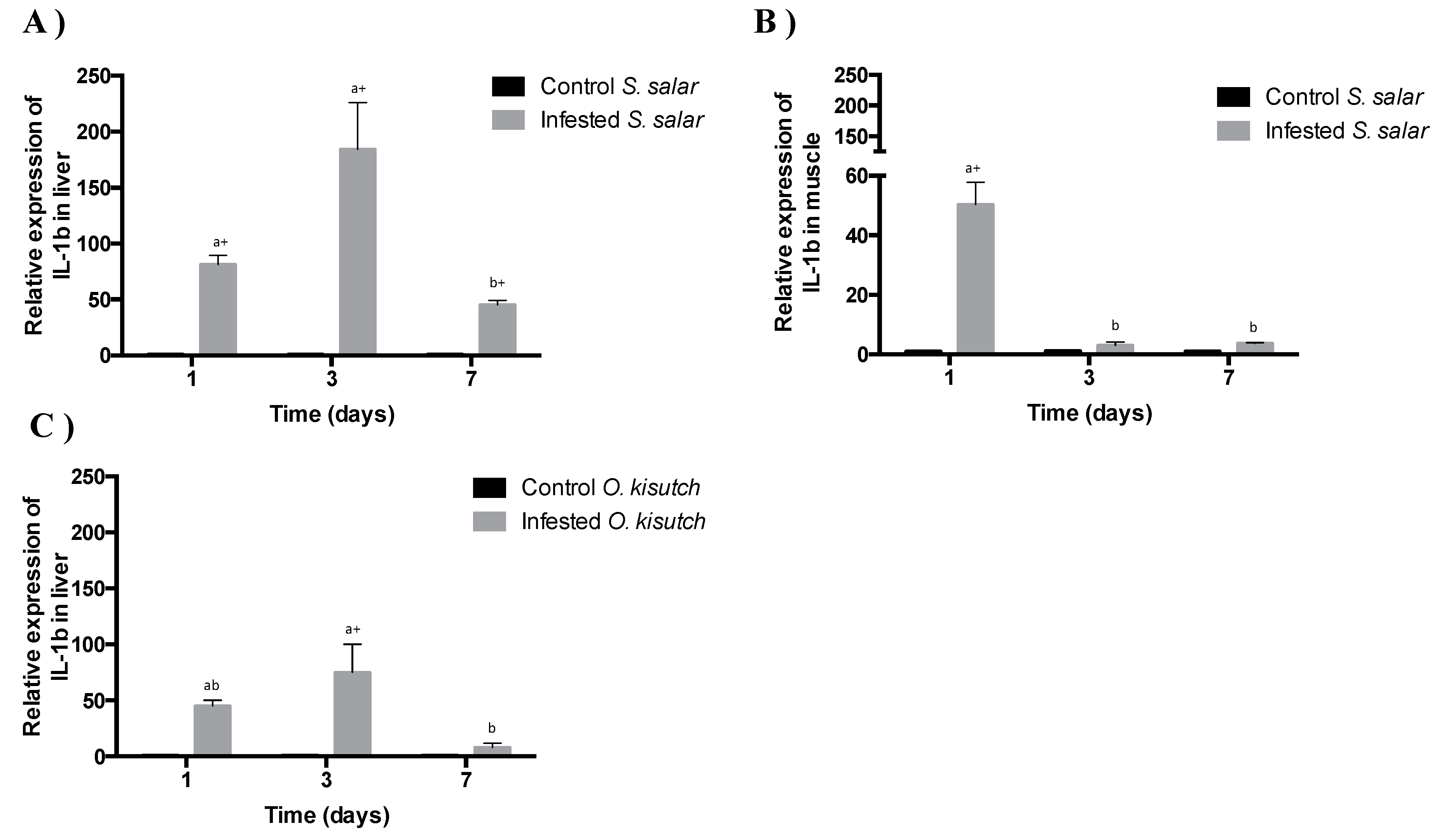
2.3. Analysis of IL-1β Transcript Expression in the Muscle and Liver of Atlantic and Coho Salmon Infested with C. rogercresseyi
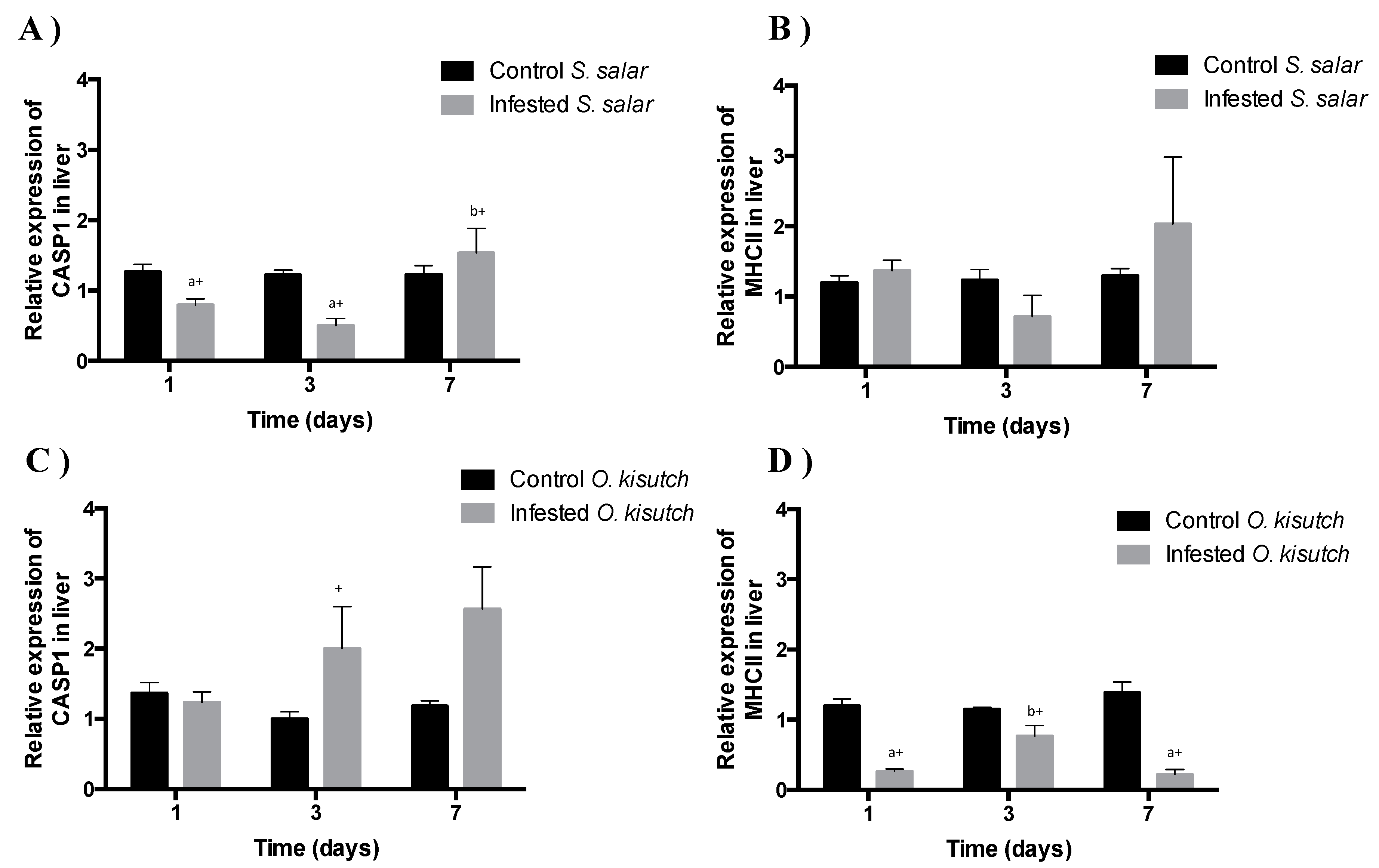
2.4. Analysis of Caspase-1 Transcript Expression in Liver of Atlantic and Coho salmon Infested with C. rogercresseyi
2.5. Analysis of MHCII Transcript Expression in the Muscle and Liver of Atlantic and Coho Salmon Infested with C. rogercresseyi
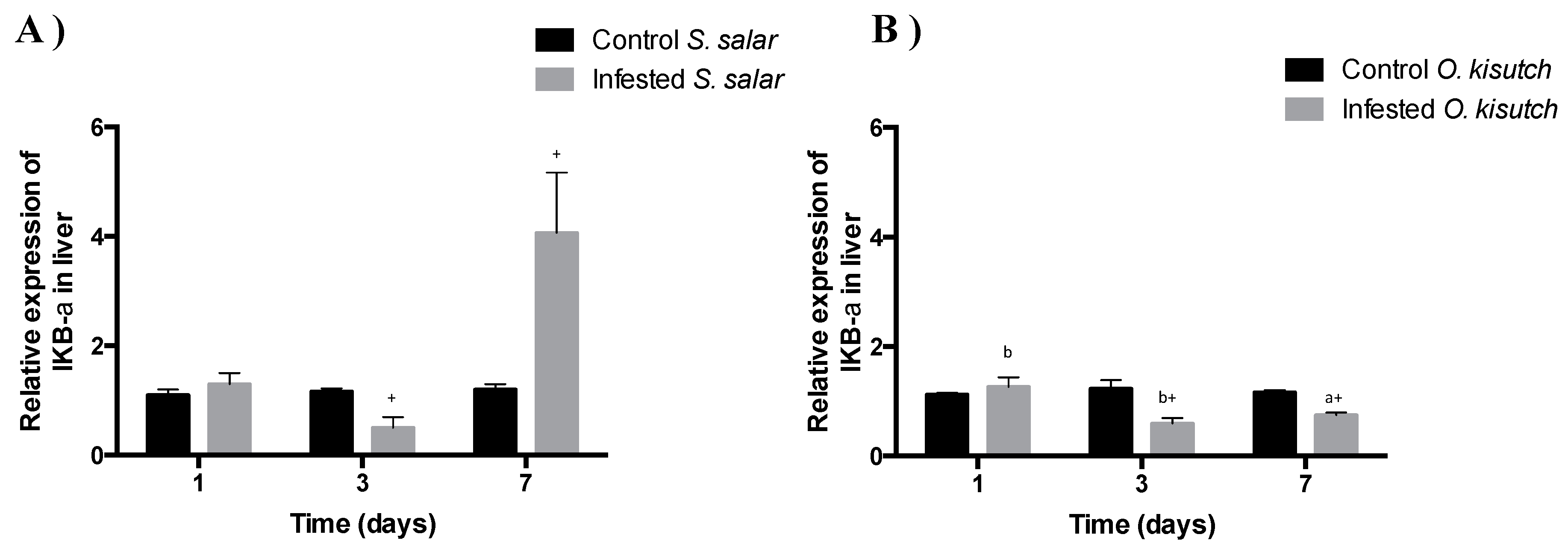
2.6. Analysis of I-kappa-B-alpha Transcript Expression in the Muscle and Liver of Atlantic and Coho Salmon Infested with C. rogercresseyi
3. Discussion
4. Materials and Methods
4.1. Fish and Experimental Design
4.2. Experimental Conditions
4.3. Sampling Procedure
4.4. Gene Expression Analyses
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rebl, A.; Goldammer, T.; Seyfert, H.M. Toll-like Receptor Signaling in Bony Fish. Vet. Immunol. Immunopathol. 2010, 134, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Vera-Bizama, F.; Valenzuela-Muñoz, V.; Gonçalves, A.T.; Marambio, J.P.; Hawes, C.; Wadsworth, S.; Gallardo-Escárate, C. Transcription expression of immune-related genes from Caligus rogercresseyi evidences host-dependent patterns on Atlantic and coho salmon. Fish Shellfish Immunol. 2015, 47, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Gallardo-Escárate, C. TLR and IMD Signaling Pathways from Caligus rogercresseyi (Crustacea: Copepoda): In Silico Gene Expression and SNPs Discovery. Fish Shellfish Immunol. 2014, 36, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.; Ting, P. Inflammasomes: Mechanism of Action, Role in Disease and Therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Ruegg, A.; Werner, S.; Beer, H.D. Active caspase-1 is a regulator of unconventional protein secretion. Cell 2008, 132, 818–831. [Google Scholar] [CrossRef]
- Beckley, D.; Reid, R.; Huang, M.; Willingham, S.; Brian, J.; Brickey, J.; Barker, B.; Kwan, M.; Taxman, D.; Accavitti-loper, A.; et al. NLRC5-Dependent Activation of the Inflammasome. J. Immunol. 2013, 186, 1333–1337. [Google Scholar]
- Pontigo, J.P.; Agüero, M.; Sanchez, P.; Oyarzún, R.; Vargas-Lagos, C.; Mancilla, J.; Kossmann, H.; Morera, F.; Yáñez, A.; Vargas-Chacoff, L. Identification and Expressional Analysis of NLRC5 Inflammasome Gene in Smolting Atlantic Salmon (Salmo salar). Fish Shellfish Immunol. 2016, 58, 259–265. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-kappa B and Rel proteins: Evolutionarily conserved mediators of immune responses. Annu. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef]
- Angosto, D.; López-Castejón, G.; López-Muñoz, A.; Sepulcre, M.P.; Arizcun, M.; Meseguer, J.; Mulero, V. Evolution of inflammasome functions in vertebrates: Inflammasome and caspase-1 trigger fish macrophage cell death but are dispensable for the processing of IL-1β. Innate Immun. 2012, 18, 815–824. [Google Scholar] [CrossRef]
- Ogura, Y.; Sutterwala, F.; Flavell, R. The inflammasome: First line of the immune response to cell stress. Cell 2006, 126, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Biswas, G.; Korenaga, H.; Nagamine, R.; Takayama, H.; Kawahara, S.; Takeda, S.; Kikuchi, Y.; Dashnyam, B.; Kono, T.; Sakai, M. Cytokine Responses in the Japanese Puffer Fish (Takifugu rubripes) Head Kidney Cells Induced with Heat-Killed Probiotics Isolated from the Mongolian Dairy Products. Fish Shellfish Immunol. 2013, 34, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Woo, P. Selected Parasitosis in Cultured and Wild Fish. Vet. Parasitol. 2009, 163, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Hamilton-West, C.; Arriagada, G.; Yatabe, T.; Valdes, P.; Herve-Claude, L.P.; Urcelay, S. Epidemiological description of the sea lice (Caligus rogercresseyi) situation in southern Chile in August 2007. Prev. Vet. Med. 2012, 104, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Bjørn, P.A.; Finstad, B.; Kristoffersen, R. Salmon lice infection of wild sea trout and Arctic char in marine and freshwaters: The effects of salmon farms. Aquac. Res. 2001, 32, 947–962. [Google Scholar] [CrossRef]
- Bravo, S. The Reproductive Output of Sea Lice Caligus rogercresseyi under Controlled Conditions. Exp. Parasitol. 2010, 125, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J. The Global Economic Cost of Sea Lice to the Salmonid Farming Industry. J. Fish Dis. 2009, 32, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Fast, M.D. Fish immune responses to parasitic copepod (namely sea lice) infection. Dev. Comp. Immunol. 2013, 43, 300–312. [Google Scholar] [CrossRef]
- Fast, M.D.; Ross, N.W.; Mustafa, A.; Sims, D.E.; Johnson, S.C.; Conboy, G.A.; Speare, D.J.; Johnson, G.; Burka, J.F. Vulnerability of rainbow trout Oncorhynchus mykiss, Atlantic salmon Salmo salar and coho salmon Oncorhynchus kisutch to experimental infection with sea lice Lepeophtheirus salmonis. Dis. Aquat. Organ. 2002, 52, 57–68. [Google Scholar] [CrossRef]
- Firth, K.; Johnson, S.; Ross, N. Characterization of Proteases in the Skin Mucus of Atlantic Salmon (Salmo salar) Infected with the Salmon Louse (Lepeophtheirus salmonis) and in Whole-Body Louse Homogenate (Salmo salar) Infected with the Salmon Louse (Lepeophtheirus salmonis). J. Parasitol. 2000, 86, 1199–1205. [Google Scholar] [CrossRef]
- Braden, L.; Koop, M.B.F.; Simon, R.M.J. Signatures of Resistance to Lepeophtheirus salmonis Include a T H 2-Type Response at the Louse-Salmon Interface. Dev. Comp. Immunol. 2015, 48, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Comparative immunity of Salmo salar and Oncorhynchus kisutch during infestation with the sea louse Caligus rogercresseyi: An enrichment transcriptome analysis. Fish Shellfish Immunol. 2016, 59, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Uncovering iron regulation with species-specific transcriptome patterns in Atlantic and coho salmon during a Caligus rogercresseyi infestation. J. Fish Dis. 2017, 40, 1169–1184. [Google Scholar] [CrossRef] [PubMed]
- Whyte, S.K. The Innate Immune Response of Finfish -A Review. Fish Shellfish Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Nie, L.; Zhu, G.; Xiang, L.; Shao, J. Advances in Research of Fish Immune-Relevant Genes: A Comparative Overview of Innate and Adaptive Immunity in Teleosts. Dev. Comp. Immunol. 2013, 39, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Sakai, M. Genomics of Fish Cytokines. Comp. Biochem. Physiol. D. 2006, 1, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Huising, M.; Stet, R.; Savelkoul, H.; Verburg-van Kemenade, B. The molecular evolution of the interleukin-1 family of cytokines; IL-18 in teleost fish. Dev. Comp. Immunol. 2004, 28, 395–413. [Google Scholar] [CrossRef] [PubMed]
- Akira, S. The Role of IL-18 in Innate Immunity. Curr. Opin. Immunol. 2000, 12, 59–63. [Google Scholar] [CrossRef]
- Biet, F.; Locht, C.; Kremer, L. Immunoregulatory Functions of Interleukin 18 and Its Role in Defense against Bacterial Pathogens. J. Mol. Med. 2002, 80, 147–162. [Google Scholar] [CrossRef]
- Gracie, J.A.; Robertson, S.E.; McInnes, I.B. Interleukin-18. J. Leukoc. Biol. 2003, 73, 213–224. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 is a unique cytokine that stimulates both Th1 and Th2 responses depending on its cytokine milieu. Cytokine Growth Factor Rev. 2001, 12, 53–72. [Google Scholar] [CrossRef]
- Biswas, G.; Bilen, S.; Kono, T.; Sakai, M.; Hikima, J. Inflammatory Immune Response by Lipopolysaccharide-Responsive Nucleotide Binding Oligomerization Domain (NOD) -like Receptors in the Japanese Puffer Fish (Takifugu rubripes). Dev. Comp. Immunol. 2016, 55, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Ramírez-Cepeda, F.; Santana, P.; Torres, E.; Cortés, J.; Guzmán, F.; Schmitt, P.; Mercado, L. Insights into the diversity of NOD-like receptors: Identification and expression analysis of NLRC3, NLRC5 and NLRX1 in rainbow trout. Mol. Immunol. 2017, 87, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Luheshi, N.M.; Compan, V.; High, S.; Whitehead, R.C.; Flitsch, S.; Kirov, A.; Prudovsky, I.; Swanton, E.; Brough, D. Deubiquitinases regulate the activity of caspase-1 and interleukin-1beta secretion via assembly of the inflammasome. J. Biol. Chem. 2013, 288, 2721–2733. [Google Scholar] [CrossRef] [PubMed]
- Ogryzko, N.; Renshaw, S.; Wilson, H. The IL-1 family in fish: Swimming through the muddy waters of inflammasome evolution. Dev. Comp. Immunol. 2014, 46, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Chacoff, L.; Muñoz, J.L.P.; Hawes, C.; Oyarzún, R.; Pontigo, J.P.; Saravia, J.; González, M.P.; Mardones, O.; Labbé, B.S.; Morera, F.J.; et al. Ectoparasite Caligus rogercresseyi modifies the lactate response in Atlantic salmon (Salmo salar) and Coho salmon (Oncorhynchus kisutch). Vet. Parasitol. 2017, 243, 6–11. [Google Scholar] [CrossRef]
- Vargas-Chacoff, L.; Muñoz, J.L.; Hawes, C.; Oyarzún, R.; Pontigo, J.P.; Saravia, J.; Gonzalez, M.P.; Morera, F.; Labbé, B.; Bertran, C.; et al. Atlantic salmon (Salmo salar) and Coho Salmon (Oncorhynchus kisutch) Display Differential Metabolic Changes in Response to Infestation by the Ectoparasite Caligus rogercresseyi. Aquaculture 2016, 464, 469–479. [Google Scholar] [CrossRef]
- Neerincx, A.; Rodriguez, G.M.; Steimle, V.; Kufer, T.A. NLRC5 Controls Basal MHC Class I Gene Expression in an MHC Enhanceosome-Dependent Manner. J. Immunol. 2012, 188, 4940–4950. [Google Scholar] [CrossRef]
- Skugor, S.; Glover, K.A.; Nilsen, F.; Krasnov, A. Local and Systemic Gene Expression Responses of Atlantic Salmon (Salmo salar L.) to Infection with the Salmon Louse (Lepeophtheirus salmonis). BMC Genom. 2008, 9, 498. [Google Scholar] [CrossRef]
- Tadiso, T.M.; Krasnov, A.; Skugor, S.; Afanasyev, S.; Hordvik, I.; Nilsen, F. Gene expression analyses of immune responses in Atlantic salmon during early stages of infection by salmon louse (Lepeophtheirus salmonis) revealed bi-phasic responses coinciding with the copepod-chalimus transition. BMC Genom. 2011, 12, 141. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Acuña, G.; Gonçalves, A.T.; Valenzuela-Muñoz, V.; Pino-Marambio, J.; Wadsworth, S.; Gallardo-Escárate, C. Transcriptome immunomodulation of in-feed additives in Atlantic Salmon Salmo salar infested with sea lice Caligus rogercresseyi. Fish Shellfish Immunol. 2015, 47, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Q.; Lu, Y.; Chen, S.; Li, Q.; Sha, Z. Expression Profiles of NODs in Channel Catfish (Ictalurus punctatus) after Infection with Edwardsiella tarda, Aeromonas hydrophila, Streptococcus iniae and Channel Catfish Hemorrhage reovirus. Fish Shellfish Immunol. 2012, 33, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, B.J.G.; Koczka, K.W.; Yasuike, M.; Jantzen, S.G.; Yazawa, R.; Koop, B.F.; Jones, S.R. Comparative transcriptomics of Atlantic Salmo salar, chum Oncorhynchus keta and pink salmon O. gorbuscha during infections with salmon lice Lepeophtheirus salmonis. BMC Genom. 2014, 15, 200. [Google Scholar] [CrossRef] [PubMed]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Barton, B. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- González, M.P.; Marín, S.L.; Vargas-Chacoff, L. Effects of Caligus rogercresseyi (Boxshall and Bravo, 2000) Infestation on Physiological Response of Host Salmo salar (Linnaeus 1758): Establishing Physiological Thresholds. Aquaculture 2015, 438, 47–54. [Google Scholar] [CrossRef]
- Gonzalez, L.; Carvajal, J. Life cycle of Caligus rogercresseyi, (Copepoda: Caligidae) parasite of Chilean reared salmonids. Aquaculture 2003, 220, 101–117. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Lekanne-Deprez, R.H.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Nucleotide Sequence (5′ ➔ 3′) | GenBank Accession nº | Efficiency Muscle (%) (S.salar/O.Kitsuch) | Efficiency Liver (%) (S.salar/O.Kitsuch) |
|---|---|---|---|---|
| NLRC5—Forward | TCTGTCTACCGTGACCATAAGCCT | XM_014149024.1 | 101.1/104.3 | 96.2/93.2 |
| NLRC5—Reverse | CCCACTCTACCAATGCTGGTCAAT | |||
| IL-18—Forward | GGAGCAACCTTTGCCTGACCAAAT | NM_001141408.1 | 103.1/105.1 | 100.3/95.7 |
| IL-18—Reverse | CTGGTCCATCCTCAAAGCTCAAGT | |||
| Il-1β—Forward | TGGGTGCACGCACATCAACAT | NM_001123582.1 | 95.8 | 103.4/94.2 |
| Il-1β—Reverse | AGGGGCGCTTACCACAATATTGAC | |||
| 18S—Forward | GTCCGGGAAACCAAAGTC | AJ427629.1 | 103.4/103.1 | 103.3/103.2 |
| 18S—Reverse | TTGAGTCAAATTAAGCCGCA | |||
| Caspase-1—Forward | TTGGCACTGAAGAGCAGGAAAGAG | 101.3/92.3 | ||
| Caspase-1—Reverse | GGCCTAAGATCAGCTTGGCAAATG | |||
| MHCII—Forward | GCAGAAGGGTCCAACAAGAG | XM_014133066.1 | 100.2/97.1 | |
| MHCII—Reverse | GCAGACTCATCGATCAGCAA | |||
| IkB-α—Forward | TAGGCCAGCTCTATGTGGCT | XM_014204687.1 | 100.8/104.6 | |
| IkB-α—Reverse | TGAGGAGGAGTGCATGTCTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pontigo, J.P.; Saravia, J.; Oyarzún, R.; González, M.P.; Hawes, C.; Morera, F.J.; Pino, J.; Wadsworth, S.; Muñoz, J.L.P.; Vargas-Chacoff, L. Modulation of the Expression of Immune-related Gene in Atlantic and Coho Salmon during Infestation with the Sea lice Caligus rogercresseyi. Fishes 2019, 4, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030042
Pontigo JP, Saravia J, Oyarzún R, González MP, Hawes C, Morera FJ, Pino J, Wadsworth S, Muñoz JLP, Vargas-Chacoff L. Modulation of the Expression of Immune-related Gene in Atlantic and Coho Salmon during Infestation with the Sea lice Caligus rogercresseyi. Fishes. 2019; 4(3):42. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030042
Chicago/Turabian StylePontigo, Juan Pablo, Julia Saravia, Ricardo Oyarzún, Margarita P. González, Christopher Hawes, Francisco J. Morera, Jorge Pino, Simon Wadsworth, Jose Luis P. Muñoz, and Luis Vargas-Chacoff. 2019. "Modulation of the Expression of Immune-related Gene in Atlantic and Coho Salmon during Infestation with the Sea lice Caligus rogercresseyi" Fishes 4, no. 3: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030042