Zebrafish Models in Neural and Behavioral Toxicology across the Life Stages



Abstract
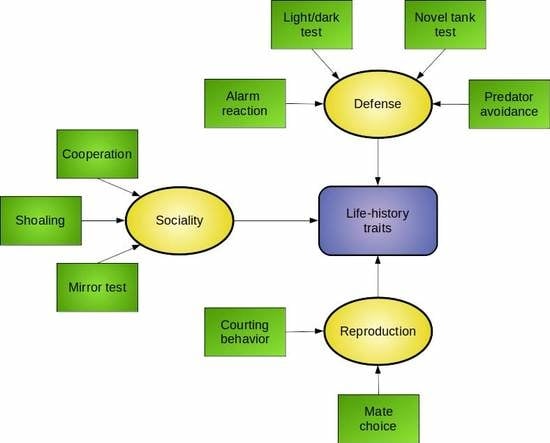
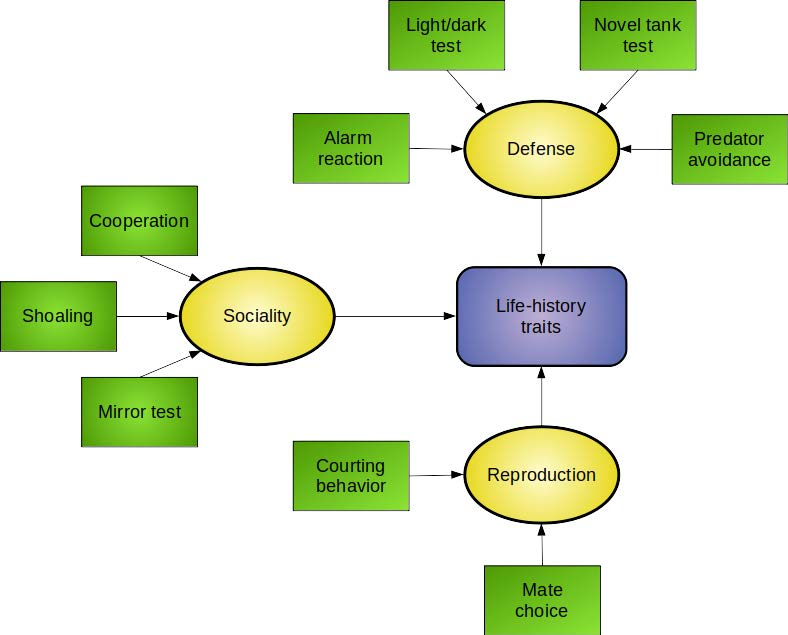
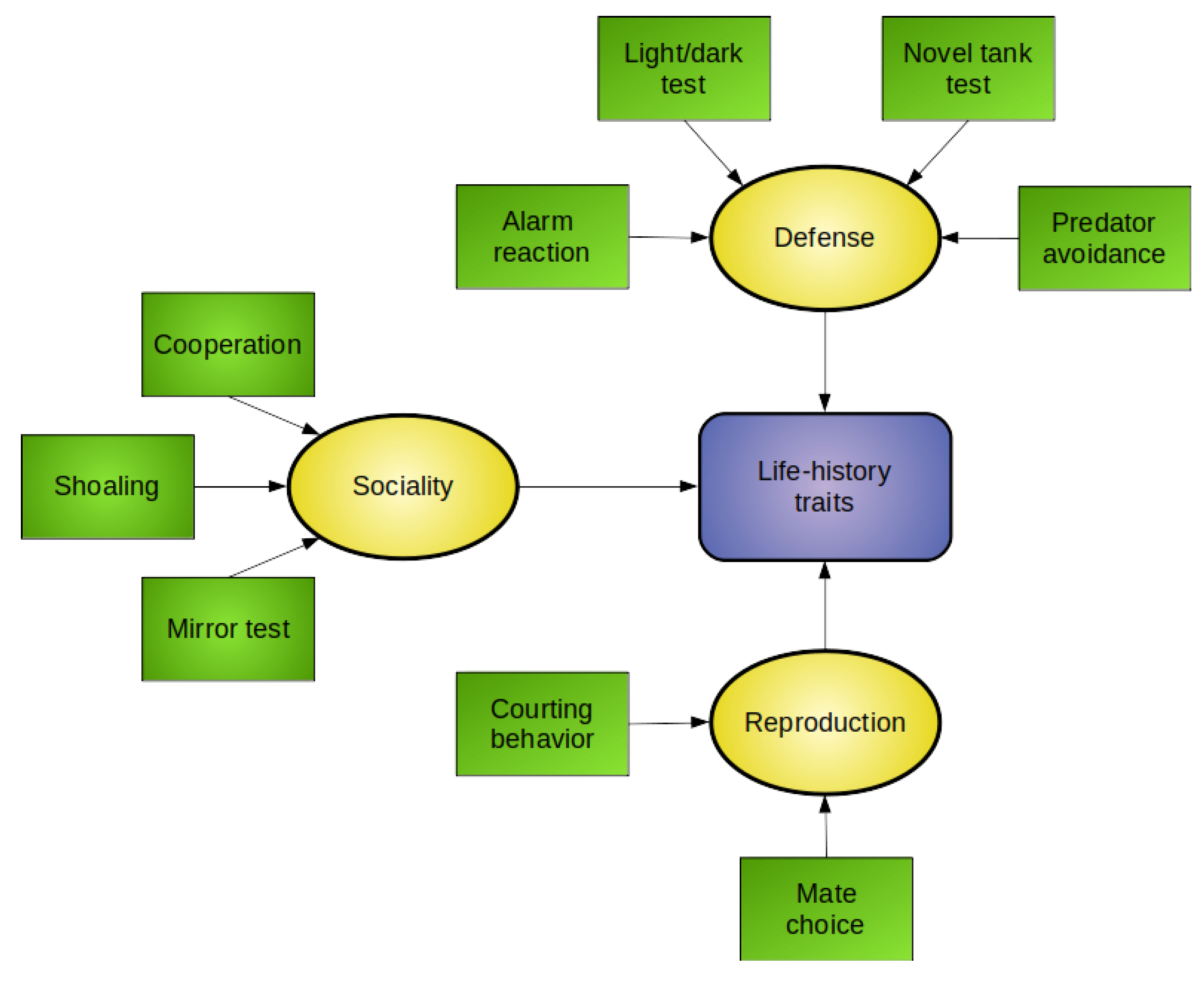
:
1. Introduction
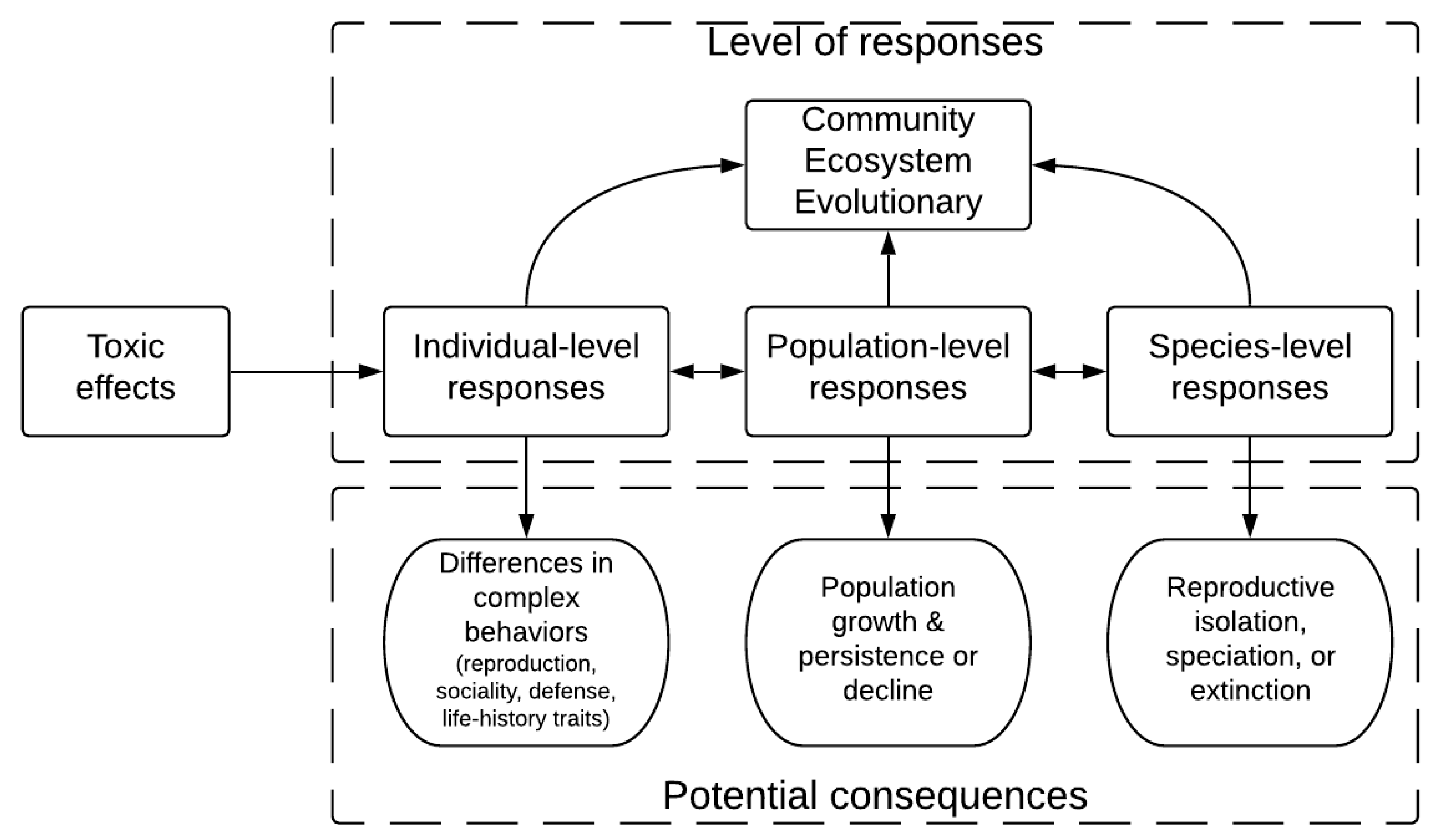
2. Using Fish as Models and Targets in Ecotoxicology
3. Behavioral Bioassays in Neurotoxicology
- Behavior is considered an early warning tool [2].
3.1. Locomotor Activity Assays
3.1.1. Larvae
3.1.2. Adults
3.2. Anxiety-Like Behavior
3.3. Aggression
4. Moving from Behavioral Toxicology to Ecotoxicology: Ecologically-Relevant Endpoints
5. Sensitivity and Specificity of Behavioral Tests
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bazerman, C.; de los Santos, R. Measuring Incommensurability: Are toxicology and ecotoxicology blind to what the other sees? In Rhetoric and Incommensurability; Harris, R., Ed.; Parlor: West Lafayette, IN, USA, 2005; pp. 424–463. [Google Scholar]
- Hellou, J. Behavioural ecotoxicology, an “early warning” signal to assess environmental quality. Environ. Sci. Pollut. Res. Int. 2011, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Legradi, J.B.; Di Paolo, C.; Kraak, M.H.S.; van der Geest, H.G.; Schymanski, E.L.; Williams, A.J.; Dingemans, M.M.L.; Massei, R.; Brack, W.; Cousin, X.; et al. An ecotoxicological view on neurotoxicity assessment. Environ. Sci. Eur. 2018, 30. [Google Scholar] [CrossRef]
- Klaminder, J.; Jonsson, M.; Leander, J.; Fahlman, J.; Brodin, T.; Fick, J.; Hellström, G. Less anxious salmon smolt become easy prey during downstream migration. Sci. Total Environ. 2019, 687, 488–493. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Lorenz, R.; Spieser, O.H. Effects of atrazine on swimming behavior of zebrafish, Brachydanio rerio. Water Res. 1995, 29, 981–985. [Google Scholar] [CrossRef]
- Maximino, C.; Araujo, J.; Leão, L.K.R.; Grisolia, A.B.A.; Oliveira, K.R.M.; Lima, M.G.; Batista, E.d.J.O.; Crespo-López, M.E.; Gouveia, A.; Herculano, A.M. Possible role of serotoninergic system in the neurobehavioral impairment induced by acute methylmercury exposure in zebrafish (Danio rerio). Neurotoxicol. Teratol. 2011, 33, 727–734. [Google Scholar] [CrossRef]
- Puty, B.; Maximino, C.; Brasil, A.; da Silva, W.L.L.; Gouveia, A.; Oliveira, K.R.M.; Batista, E.d.J.O.; Crespo-Lopez, M.E.; Rocha, F.A.F.; Herculano, A.M. Ascorbic Acid Protects Against Anxiogenic-Like Effect Induced by Methylmercury in Zebrafish: Action on the Serotonergic System. Zebrafish 2014, 11, 365–370. [Google Scholar] [CrossRef]
- Glazer, L.; Hahn, M.E.; Aluru, N. Delayed effects of developmental exposure to low levels of the aryl hydrocarbon receptor agonist 3,3′,4,4′,5-pentachlorobiphenyl (PCB126) on adult zebrafish behavior. NeuroToxicology 2016, 52, 134–143. [Google Scholar] [CrossRef] [Green Version]
- Sackerman, J.; Donegan, J.J.; Cunningham, C.S.; Nguyen, N.N.; Lawless, K.; Long, A.; Benno, R.H.; Gould, G.G. Zebrafish Behavior in Novel Environments: Effects of Acute Exposure to Anxiolytic Compounds and Choice of Danio rerio Line. Int. J. Comp. Psychol. 2010, 23, 43–61. [Google Scholar]
- Haverroth, G.M.B.; Welang, C.; Mocelin, R.N.; Postay, D.; Bertoncello, K.T.; Franscescon, F.; Rosemberg, D.B.; Dal Magro, J.; Dalla Corte, C.L. Copper acutely impairs behavioral function and muscle acetylcholinesterase activity in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2015, 122, 440–447. [Google Scholar] [CrossRef]
- Weber, P.; Vogel, C.; Lang, C.; Baldisserotto, B. Antipredator and alarm reaction responses of silver catfish (Rhamdia quelen) juveniles exposed to waterborne ammonia. Neotropical Ichthyol. 2012, 10, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Tierney, K.B.; Taylor, A.L.; Ross, P.S.; Kennedy, C.J. The alarm reaction of coho salmon parr is impaired by the carbamate fungicide IPBC. Aquat. Toxicol. 2006, 79, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Norrgren, L. Fish models for ecotoxicology. Acta Vet. Scand. 2012, 54, S14. [Google Scholar] [CrossRef] [Green Version]
- Rand, G.M. Behavior. In Fundamentals of Aquatic Toxicology: Methods and Applications; Rand, G.M., Petrocelli, S.R., Eds.; FMC Corp.: Princeton, NJ, USA, 1985; pp. 221–263. [Google Scholar]
- Faimali, M.; Gambardella, C.; Costa, E.; Piazza, V.; Morgana, S.; Estévez-Calvar, N.; Garaventa, F. Old model organisms and new behavioral end-points: Swimming alteration as an ecotoxicological response. Mar. Environ. Res. 2017, 128, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ankeny, R.A.; Leonelli, S. What’s so special about model organisms? Stud. Hist. Philos. Sci. Part A 2011, 42, 313–323. [Google Scholar] [CrossRef]
- Leonelli, S.; Ankeny, R.A. What makes a model organism? Endeavour 2013, 37, 209–212. [Google Scholar] [CrossRef] [Green Version]
- Berzas Nevado, J.J.; Rodríguez Martín-Doimeadios, R.C.; Guzmán Bernardo, F.J.; Jiménez Moreno, M.; Herculano, A.M.; do Nascimento, J.L.M.; Crespo-López, M.E. Mercury in the Tapajós River basin, Brazilian Amazon: A review. Environ. Int. 2010, 36, 593–608. [Google Scholar] [CrossRef]
- Planchart, A.; Mattingly, C.J.; Allen, D.; Ceger, P.; Casey, W.; Hinton, D.; Kanungo, J.; Kullman, S.W.; Tal, T.; Bondesson, M.; et al. Advancing toxicology research using in vivo high throughput toxicology with small fish models. ALTEX 2016, 33, 435–452. [Google Scholar] [CrossRef] [Green Version]
- Carvan, M.J.; Incardona, J.P.; Rise, M.L. Meeting the Challenges of Aquatic Vertebrate Ecotoxicology. BioScience 2008, 58, 1015–1025. [Google Scholar] [CrossRef] [Green Version]
- Embry, M.R.; Belanger, S.E.; Braunbeck, T.A.; Galay-Burgos, M.; Halder, M.; Hinton, D.E.; Léonard, M.A.; Lillicrap, A.; Norberg-King, T.; Whale, G. The fish embryo toxicity test as an animal alternative method in hazard and risk assessment and scientific research. Aquat. Toxicol. 2010, 97, 79–87. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.T.; Van Rompay, A.R.; De Coen, W.; Witters, H.E. Development of a screening assay to identify teratogenic and embryotoxic chemicals using the zebrafish embryo. Reprod. Toxicol. 2009, 28, 308–320. [Google Scholar] [CrossRef]
- Dai, Y.-J.; Jia, Y.-F.; Chen, N.; Bian, W.-P.; Li, Q.-K.; Ma, Y.-B.; Chen, Y.-L.; Pei, D.-S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Fish in behavior research: Unique tools with a great promise! J. Neurosci. Methods 2014, 234, 54–58. [Google Scholar] [CrossRef] [PubMed]
- OECD. OECD, Guidelines for the Testing of Chemicals, No. 203: Fish Acute Toxicity Test; OECD: Paris, France, 2019. [Google Scholar] [CrossRef]
- OECD. OECD, Guidelines for the Testing of Chemicals, Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD: Paris, France. [CrossRef]
- McKim, J.M.; Schmieder, P.K.; Carlson, R.W.; Hunt, E.P.; Niemi, G.J. Use of respiratory-cardiovascular responses of rainbow trout (Salmo gairdneri) in identifying acute toxicity syndromes in fish: Part 1. pentachlorophenol, 2,4-dinitrophenol, tricaine methanesulfonate and 1-octanol. Environ. Toxicol. Chem. 1987, 6, 295–312. [Google Scholar] [CrossRef]
- McKim, J.M.; Schmieder, P.K.; Niemi, G.J.; Carlson, R.W.; Henry, T.R. Use of respiratory-cardiovascular responses of rainbow trout (Salmo gairdneri) in identifying acute toxicity syndromes in fish: Part 2. malathion, carbaryl, acrolein and benzaldehyde. Environ. Toxicol. Chem. 1987, 6, 313–328. [Google Scholar] [CrossRef]
- Bradbury, S.P.; Carlson, R.W.; Niemi, G.J.; Henry, T.R. Use of respiratory-cardiovascular responses of rainbow trout (Oncorhynchus mykiss) in identifying acute toxicity syndromes in fish: Part 4. Central nervous system seizure agents. Environ. Toxicol. Chem. 1991, 10, 115–131. [Google Scholar] [CrossRef]
- Peterson, E.K.; Buchwalter, D.B.; Kerby, J.L.; LeFauve, M.K.; Varian-Ramos, C.W.; Swaddle, J.P. Integrative behavioral ecotoxicology: Bringing together fields to establish new insight to behavioral ecology, toxicology, and conservation. Curr. Zool. 2017, 63, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handy, R.D.; Depledge, M.H. Physiological Responses: Their Measurement and Use as Environmental Biomarkers in Ecotoxicology. Ecotoxicology 1999, 8, 329–349. [Google Scholar] [CrossRef]
- Wester, P.W.; van der Ven, L.T.M.; Vethaak, A.D.; Grinwis, G.C.M.; Vos, J.G. Aquatic toxicology: Opportunities for enhancement through histopathology. Environ. Toxicol. Pharmacol. 2002, 11, 289–295. [Google Scholar] [CrossRef]
- Brandão, F.P.; Rodrigues, S.; Castro, B.B.; Gonçalves, F.; Antunes, S.C.; Nunes, B. Short-term effects of neuroactive pharmaceutical drugs on a fish species: Biochemical and behavioural effects. Aquat. Toxicol. 2013, 144, 218–229. [Google Scholar] [CrossRef]
- Scott, G.R.; Sloman, K.A. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef]
- Gerhardt, A. Aquatic Behavioral Ecotoxicology—Prospects and Limitations. Hum. Ecol. Risk Assess. Int. J. 2007, 13, 481–491. [Google Scholar] [CrossRef]
- Brown, R.E.; Bolivar, S. The importance of behavioural bioassays in neuroscience. J. Neurosci. Methods 2018, 300, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Maximino, C.; van der Staay, F.J. Behavioral models in psychopathology: Epistemic and semantic considerations. Behav. Brain Funct. 2019, 15, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, P. The validity of animal models of depression. Psychopharmacology 1984, 83, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Kysil, E.V.; Meshalkina, D.A.; Frick, E.E.; Echevarria, D.J.; Rosemberg, D.B.; Maximino, C.; Lima, M.G.; Abreu, M.S.; Giacomini, A.C.; Barcellos, L.J.G.; et al. Comparative Analyses of Zebrafish Anxiety-Like Behavior Using Conflict-Based Novelty Tests. Zebrafish 2017, 14, 197–208. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Blanchard, R.J. Ethoexperimental approaches to the biology of emotion. Annu. Rev. Psychol. 1988, 39, 43–68. [Google Scholar] [CrossRef]
- Soares, M.C.; Gerlai, R.; Maximino, C. The integration of sociality, monoamines and stress neuroendocrinology in fish models: Applications in the neurosciences. J. Fish Biol. 2018, 93, 170–191. [Google Scholar] [CrossRef]
- Maximino, C.; de Brito, T.M.; da Silva Batista, A.W.; Herculano, A.M.; Morato, S.; Gouveia, A. Measuring anxiety in zebrafish: A critical review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef]
- Way, G.P.; Ruhl, N.; Snekser, J.L.; Kiesel, A.L.; McRobert, S.P. A comparison of methodologies to test aggression in zebrafish. Zebrafish 2015, 12, 144–151. [Google Scholar] [CrossRef]
- Gerlai, R. Antipredatory behavior of zebrafish: Adaptive function and a tool for translational research. Evol. Psychol. 2013, 11, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Blaser, R.E.; Vira, D.G. Experiments on learning in zebrafish (Danio rerio): A promising model of neurocognitive function. Neurosci. Biobehav. Rev. 2014, 42, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Irons, T.D.; MacPhail, R.C.; Hunter, D.L.; Padilla, S. Acute neuroactive drug exposures alter locomotor activity in larval zebrafish. Neurotoxicol. Teratol. 2010, 32, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Selderslaghs, I.W.T.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Drapeau, P.; Saint-Amant, L.; Buss, R.R.; Chong, M.; McDearmid, J.R.; Brustein, E. Development of the locomotor network in zebrafish. Prog. Neurobiol. 2002, 68, 85–111. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Echevarria, D.J.; Homechaudhuri, S.; Stewart, A.M.; Collier, A.D.; Kaluyeva, A.A.; Li, S.; Liu, Y.; Chen, P.; Wang, J.; et al. Zebrafish neurobehavioral phenomics for aquatic neuropharmacology and toxicology research. Aquat. Toxicol. 2016, 170, 297–309. [Google Scholar] [CrossRef]
- Burgess, H.A.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Noldus, L.P.J.J.; Tegelenbosch, R.A.J.; Richardson, M.K. Zebrafish embryos and larvae in behavioural assays. Behaviour 2012, 149, 1241–1281. [Google Scholar]
- Ellis, L.D.; Seibert, J.; Soanes, K.H. Distinct models of induced hyperactivity in zebrafish larvae. Brain Res. 2012, 1449, 46–59. [Google Scholar] [CrossRef]
- Boehmler, W.; Carr, T.; Thisse, C.; Thisse, B.; Canfield, V.A.; Levenson, R. D4 Dopamine receptor genes of zebrafish and effects of the antipsychotic clozapine on larval swimming behaviour. Genes Brain Behav. 2007, 6, 155–166. [Google Scholar] [CrossRef]
- Airhart, M.J.; Lee, D.H.; Wilson, T.D.; Miller, B.E.; Miller, M.N.; Skalko, R.G. Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC). Neurotoxicol. Teratol. 2007, 29, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, C.; Hu, C.; Yu, K.; Yang, L.; Zhoug, B. Acute exposure to DE-71: Effects on locomotor behavior and developmental neurotoxicity in zebrafish larvae. Environ. Toxicol. Chem. 2012, 31, 2338–2344. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.G.; Zhou, R.; Jiang, D.; Song, J.E.; Xu, Q.; Si, J.; Chen, Y.P.; Zhou, X.; Gan, L.; Li, J.Z.; et al. Toxicity of Graphene Quantum Dots in Zebrafish Embryo. Biomed. Environ. Sci. 2015, 28, 341–351. [Google Scholar] [PubMed]
- Duan, J.; Yu, Y.; Shi, H.; Tian, L.; Guo, C.; Huang, P.; Zhou, X.; Peng, S.; Sun, Z. Toxic Effects of Silica Nanoparticles on Zebrafish Embryos and Larvae. PLoS ONE 2013, 8, e74606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Huang, C.; Wang, L.; Ye, X.; Bai, C.; Simonich, M.T.; Tanguay, R.L.; Dong, Q. Toxicity, uptake kinetics and behavior assessment in zebrafish embryos following exposure to perfluorooctanesulphonicacid (PFOS). Aquat. Toxicol. 2010, 98, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Saili, K.S.; Corvi, M.M.; Weber, D.N.; Patel, A.U.; Das, S.R.; Przybyla, J.; Anderson, K.A.; Tanguay, R.L. Neurodevelopmental low-dose bisphenol A exposure leads to early life-stage hyperactivity and learning deficits in adult zebrafish. Toxicology 2012, 291, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Ton, C.; Lin, Y.; Willett, C. Zebrafish as a model for developmental neurotoxicity testing. Birt. Defects Res. Part A Clin. Mol. Teratol. 2006, 76, 553–567. [Google Scholar] [CrossRef]
- Deeti, S.; O’Farrell, S.; Kennedy, B.N. Early safety assessment of human oculotoxic drugs using the zebrafish visualmotor response. J. Pharmacol. Toxicol. Methods 2014, 69, 1–8. [Google Scholar] [CrossRef]
- Richards, F.M.; Alderton, W.K.; Kimber, G.M.; Liu, Z.; Strang, I.; Redfern, W.S.; Valentin, J.-P.; Winter, M.J.; Hutchinson, T.H. Validation of the use of zebrafish larvae in visual safety assessment. J. Pharmacol. Toxicol. Methods 2008, 58, 50–58. [Google Scholar] [CrossRef]
- Rice, C.; Ghorai, J.K.; Zalewski, K.; Weber, D.N. Developmental lead exposure causes startle response deficits in zebrafish. Aquat. Toxicol. 2011, 105, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.N. Dose-dependent effects of developmental mercury exposure on C-start escape responses of larval zebrafish Danio rerio. J. Fish Biol. 2006, 69, 75–94. [Google Scholar] [CrossRef]
- Xu, T.; Liu, Y.; Pan, R.; Zhang, B.; Yin, D.; Zhao, J.; Zhao, Q. Vision, Color Vision, and Visually Guided Behavior: The Novel Toxicological Targets of 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47). Environ. Sci. Technol. Lett. 2017, 4, 132–136. [Google Scholar] [CrossRef]
- Dehnert, G.K.; Karasov, W.H.; Wolman, M.A. 2,4-Dichlorophenoxyacetic acid containing herbicide impairs essential visually guided behaviors of larval fish. Aquat. Toxicol. 2019, 209, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Little, E.E.; Finger, S.E. Swimming behavior as an indicator of sublethal toxicity in fish. Environ. Toxicol. Chem. 1990, 9, 13–19. [Google Scholar] [CrossRef]
- Kane, A.S.; Salierno, J.D.; Brewer, S.K.; Salierno, J.D.; Brewer, S.K. Fish Models in Behavioral Toxicology: Automated Techniques, Updates and Perspectives. In Methods in Aquatic Toxicology; Ostrander, G.K., Ed.; Lewis Publishers: Boca Raton, FL, USA, 2005; Chapter 23; pp. 559–590. [Google Scholar]
- Stewart, A.; Gaikwad, S.; Kyzar, E.; Green, J.; Roth, A.; Kalueff, A.V. Modeling anxiety using adult zebrafish: A conceptual review. Neuropharmacology 2012, 62, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.M.; Kaluyeva, A.A.; Poudel, M.K.; Nguyen, M.; Song, C.; Kalueff, A.V. Building Zebrafish Neurobehavioral Phenomics: Effects of Common Environmental Factors on Anxiety and Locomotor Activity. Zebrafish 2015, 12, 339–348. [Google Scholar] [CrossRef]
- King, J.A. The Ecology of Aggressive Behavior. Annu. Rev. Ecol. Syst. 1973, 4, 117–138. [Google Scholar] [CrossRef]
- Lorenz, K. On Aggression; Routledge: England, UK, 2002; ISBN 978-0-203-99426-9. [Google Scholar]
- Colman, J.R.; Baldwin, D.; Johnson, L.L.; Scholz, N.L. Effects of the synthetic estrogen, 17α-ethinylestradiol, on aggression and courtship behavior in male zebrafish (Danio rerio). Aquat. Toxicol. 2009, 91, 346–354. [Google Scholar] [CrossRef]
- Söffker, M.; Stevens, J.R.; Tyler, C.R. Comparative Breeding and Behavioral Responses to Ethinylestradiol Exposure in Wild and Laboratory Maintained Zebrafish (Danio rerio) Populations. Environ. Sci. Technol. 2012, 46, 11377–11383. [Google Scholar] [CrossRef]
- Chen, J.; Tanguay, R.L.; Simonich, M.; Nie, S.; Zhao, Y.; Li, L.; Bai, C.; Dong, Q.; Huang, C.; Lin, K. TBBPA chronic exposure produces sex-specific neurobehavioral and social interaction changes in adult zebrafish. Neurotoxicol. Teratol. 2016, 56, 9–15. [Google Scholar] [CrossRef]
- Strungaru, S.-A.; Robea, M.A.; Plavan, G.; Todirascu-Ciornea, E.; Ciobica, A.; Nicoara, M. Acute exposure to methylmercury chloride induces fast changes in swimming performance, cognitive processes and oxidative stress of zebrafish (Danio rerio) as reference model for fish community. J. Trace Elem. Med. Biol. 2018, 47, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.E.; Nunes, M.E.; Menezes, C.C.; Marins, A.T.; Leitemperger, J.; Gressler, A.C.L.; Carvalho, F.B.; de Freitas, C.M.; Quadros, V.A.; Fachinetto, R.; et al. Sodium Selenite Prevents Paraquat-Induced Neurotoxicity in Zebrafish. Mol. Neurobiol. 2018, 55, 1928–1941. [Google Scholar] [CrossRef]
- Little, E.E.; Dwyer, F.J.; Fairchild, J.F.; Delonay, A.J.; Zajicejk, J.L. Survival of bluegill and their behavioral responses during continuous and pulsed exposures to esfenvalerate, a pyrethroid insecticide. Environ. Toxicol. Chem. 1993, 12, 871–878. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Ren-Patterson, R.F.; LaPorte, J.L.; Murphy, D.L. Domain interplay concept in animal models of neuropsychiatric disorders: A new strategy for high-throughput neurophenotyping research. Behav. Brain Res. 2008, 188, 243–249. [Google Scholar] [CrossRef]
- Cairns, J. The threshold problem in ecotoxicology. Ecotoxicology 1992, 1, 3–16. [Google Scholar] [CrossRef]
- Dell’Omo, G. Behavioural Ecotoxicology; John Wiley & Sons: Hoboken, NJ, USA, 2002; ISBN 978-0-471-96852-8. [Google Scholar]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Altman, D.G.; Bland, J.M. Diagnostic tests. 1: Sensitivity and specificity. BMJ 1994, 308, 1552. [Google Scholar] [CrossRef] [Green Version]
- Begum, G.; Rao, J.V.; Srikanth, K. Oxidative stress and changes in locomotor behavior and gill morphology of Gambusia affinis exposed to chromium. Toxicol. Environ. Chem. 2006, 88, 355–365. [Google Scholar] [CrossRef]
- Mishra, A.K.; Mohanty, B. Acute toxicity impacts of hexavalent chromium on behavior and histopathology of gill, kidney and liver of the freshwater fish, Channa punctatus (Bloch). Environ. Toxicol. Pharmacol. 2008, 26, 136–141. [Google Scholar] [CrossRef]
- Rao, J.V.; Begum, G.; Jakka, N.M.; Srikanth, K.; Rao, R.N. Sublethal Effects of Profenofos on Locomotor Behavior and Gill Architecture of the Mosquito Fish, Gambusia affinis. Drug Chem. Toxicol. 2006, 29, 255–267. [Google Scholar] [CrossRef]
- Tellis, M.S.; Alsop, D.; Wood, C.M. Effects of copper on the acute cortisol response and associated physiology in rainbow trout. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 281–289. [Google Scholar] [CrossRef]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef]
- Cairns, J. Ethics in science: Ecotoxicology. Ethics Sci. Environ. Politics 2003, 3, 33–39. [Google Scholar] [CrossRef]
- Eckersley, R. Diving Evolution: The Ecological Ethics of Murray Bookchin. Env. Ethics. 1989, 11, 99–116. [Google Scholar] [CrossRef]
- Padovan, D. Social Morals and Ethics of Nature: From Peter Kropotkin to Murray Bookchin. Democr. Nat. 1999, 5, 485–500. [Google Scholar]
- Cairns, J. Putting the eco in ecotoxicology. Regul. Toxicol. Pharmacol. 1988, 8, 226–238. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Class | Compound | Concentration Range | Age | Ref. |
|---|---|---|---|---|
| Sedative-like | 4-aminopyridine | 0.6 mM | 5 dpf | [53] |
| Clozapine | 12.5–50 mM | 7 dpf | [54] | |
| Cocaine | 0.2–50 μM | 6 dpf | [47] | |
| Amphetamine | 0.1–20 μM | 6 dpf | [47] | |
| Diazepam | 10–100 nM | 7–14 dpf | [47] | |
| Fluoxetine | 4.6 mM | 3–6 dpf | [55] | |
| Polybrominated diphenyl ether (PBDE-71) | 63.8136 nM | 5 dpf | [56] | |
| Graphene quantum dots (light period) | 1.0408–16.6528 mM | 4–96 hpf | [57] | |
| Silica nanoparticles (dark period) | 416.1119 and 832.2237 nM | 4–96 hpf | [58] | |
| Perfluorooctanesulphonic acid (PFOS) | 0–15.9958 µM | 6–120 hpf | [59] | |
| Stimulant-like | Silica nanoparticles (dark period) | 1.664 and 3.328 µM | 4–96 hpf | [58] |
| 4-Aminopyiridine | 0.8–2.5 mM | 5 | [53] | |
| Aconitine | 2.5–25 µM | 5 | [53] | |
| Bisphenol A | 0.01–1 µM | 5 | [60] | |
| Pentylenetetrazole | 10 mM | 5 | [53] |
| Behavior | Age at Observations | Stimulus | Example Compound |
|---|---|---|---|
| Coiling | 17–21 hpf | None | Perfluorooctanesulfonic acid (PFOS ) [59] Chlorpyrifos [48] |
| Touch-induced escape responses (touch response) | 22–27 hpf | Touch | Dichlorodiphenyltrichloroethane (DDT) [61] Dieldrin [61] Fipronil [61] Nonylphenol [61] |
| Optokinetic response (OKR) | 73–80 hpf | Moving objects | Digoxin [62] Gentamicin [62] Ibuprofen [62] Minoxidil [62] Quinine [62] |
| Optomotor response | 5 dpf | Moving objects | Bisoprolol [63] Chlorpromazine [63] Cisapride [63] Cisplatin [63] Gentamicin [63] Nicotinic acid [63] Quinine [63] |
| Startle responses | 5 dpf onwards | Vibrational or acoustic stimuli | Lead [64] Mercury [65] |
| Shadow response | 8 dpf | Looming shadows | 2,2′,4,4′-tetrabromodiphenyl ether (BDE-47) [66] |
| Prey capture | 9 dpf | Prey | 2,4-Dichlorophenoxyacetic acid (2,4-D) [67] |
| Toxicant | Concentration/Dose Range | Duration of Treatment | Test | Ref. |
|---|---|---|---|---|
| Atrazine | 23.1825 nM–14.4891 μM | 4 weeks | Light/dark test (LDT) | [5] |
| Methylmercury | 1–5 mg/kg | Acute | LDT, Novel tank test (NTT) | [6,7] |
| PCB126 | 0.3–1.2 nM | Developmental (4–24 hpf) | NTT | [8] |
| Dimethyl sulfoxide | 7.05 mM | Acute | NTT | [9] |
| Copper sulfate | 60.0769 nM | Acute | LDT, NTT | [10] |
| Toxicant | Concentration/Dose Range | Duration of Treatment | Test | Ref. |
|---|---|---|---|---|
| 17α-ethinylestradiol | 1.6869–168.6893 pM | 48 h | Dyadic interaction | [74] |
| 13.4951–74.2233 pM | 14 days | Group interaction | [75] | |
| Tetrabromobisphenol A (TBBPA) | 5–50 nM | Developmental (1–120 dpf) | Mirror test | [76] |
| Methylmercury | 4.6376–69.5636 nM | 32 h | Mirror test | [77] |
| Paraquat | 20 mg/kg | 6 injections for 16 days | Mirror test | [78] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutra Costa, B.P.; Aquino Moura, L.; Gomes Pinto, S.A.; Lima-Maximino, M.; Maximino, C. Zebrafish Models in Neural and Behavioral Toxicology across the Life Stages. Fishes 2020, 5, 23. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030023
Dutra Costa BP, Aquino Moura L, Gomes Pinto SA, Lima-Maximino M, Maximino C. Zebrafish Models in Neural and Behavioral Toxicology across the Life Stages. Fishes. 2020; 5(3):23. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030023
Chicago/Turabian StyleDutra Costa, Bruna Patricia, Layana Aquino Moura, Sabrina Alana Gomes Pinto, Monica Lima-Maximino, and Caio Maximino. 2020. "Zebrafish Models in Neural and Behavioral Toxicology across the Life Stages" Fishes 5, no. 3: 23. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030023