Development of Carbon Dioxide Barriers to Deter Invasive Fishes: Insights and Lessons Learned from Bigheaded Carp


Department of Natural Resources and Environmental Sciences, University of Illinois, 1102 S Goodwin Ave, Urbana, IL 61801, USA
Fishes 2020, 5(3), 25; https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030025
Submission received: 2 June 2020
/
Revised: 29 July 2020
/
Accepted: 4 August 2020
/
Published: 13 August 2020
(This article belongs to the Special Issue Biology and Control of Invasive Fishes)
Abstract
:Invasive species are a threat to biodiversity in freshwater. Removing an aquatic invasive species following arrival is almost impossible, and preventing introduction is a more viable management option. Bigheaded carp are an invasive fish spreading throughout the Midwestern United States and are threatening to enter the Great Lakes. This review outlines the development of carbon dioxide gas (CO2) as a non-physical barrier that can be used to deter the movement of fish and prevent further spread. Carbon dioxide gas could be used as a deterrent either to cause avoidance (i.e., fish swim away from zones of high CO2), or by inducing equilibrium loss due to the anesthetic properties of CO2 (i.e., tolerance). The development of CO2 as a fish deterrent started with controlled laboratory experiments demonstrating stress and avoidance, and then progressed to larger field applications demonstrating avoidance at scales that approach real-world scenarios. In addition, factors that influence the effectiveness of CO2 as a fish barrier are discussed, outlining conditions that could make CO2 less effective in the field; these factors that influence efficacy would be of interest to managers using CO2 to target other fish species, or those using other non-physical barriers for fish.
1. Background
The transport of species beyond their native range represents a major global problem. The arrival of an invasive species can lead to the suppression of native populations through competition, the introduction of pathogens, predation, hybridization, and disruptions to habitats and ecosystem function [1,2,3]. Invasive species are therefore believed to be the second most important driver of species extinctions after habitat loss [4], and can lead to billions of dollars in economic costs [1,5]. More importantly, the decrease in biodiversity that invasive species cause can threaten human health and well-being [1,6,7,8]. Freshwater environments are experiencing declines in biodiversity disproportionately large relative to other biomes [9,10], and invasive species are one reason for this decline [1,11]. Studies have suggested that almost 40% of North American freshwater and diadromous fishes are imperiled [12], and the pace at which freshwater fish are becoming imperiled exceeds other vertebrates, and appears to be accelerating [12,13]; invasive species are a key factor contributing to these declines [4,12]. The rate at which humans have been introducing species beyond their native ranges has also accelerated over the past hundred years, driven primarily by the growth in global trade and mobility [3,14]. More importantly, models suggest that the transport of invasive species around the planet is likely to increase in the future [15,16].
While the eradication of an invasive species is theoretically possible, the unfortunate reality is that, once a species is introduced into an area, its removal is often impossible. For an invasive species to be successfully eradicated, a number of conditions must be met. These conditions include: proper planning and establishing lines of authority, a commitment to complete the eradication effort in terms of resources and enthusiasm, the biology of the target species must be amenable with the entire population of the target species put at risk, the target population must be removed faster than it can reproduce, the target species must be detectable at low densities, and efforts must be made to prevent re-invasion (possibly through restoration activities) [17,18]. These conditions are easiest to meet for isolated, small populations with low reproductive rates and poor dispersal capabilities, often for terrestrial vertebrates, with plants and aquatic species proving more challenging [17,18]. Thus, owing to the challenges associated with eradication efforts, the literature is rich with examples of failed attempts to extirpate invasive species, despite efforts that have extended over many years [18,19,20]. In some situations, the goal of completely removing an invasive species can be considered controversial as eradication can be costly, unlikely to succeed, and may result in considerable damage to non-target organisms and the environment [17,18,20]. Owing to the obstacles associated with eradication, a common outcome following the invasion of a species is “maintenance management”, whereby the goal of elimination is abandoned, and the invader is simply controlled to a density that is deemed tolerable and allowed to persist [18,19]. Therefore, to avoid this sustained presence of an invasive species and perpetual “maintenance management”, a more cost-effective, and meaningful approach to invasive species management is to prevent the arrival of an invasive species prior to invasion [1,21,22], or deter the secondary spread of invaders should they arrive [23].
2. Bigheaded Carp
Carps from the family Cyprinidae have been introduced outside of their native range for centuries. Bigheaded carps [24], and particularly bighead carp (Hypophthalmichthys nobilis) and silver carp (H. molitrix), have been introduced widely as phytoplankton control organisms in commercial aquaculture ponds and sewage lagoons owing to their large size and ability to efficiently filter phytoplankton and zooplankton from the water column [25]. Following transport to the United States for use as a biological control agent, floods allowed bigheaded carp to escape into the Mississippi River where they have spread throughout the basin [25], undeterred by locks, dams or other flood control structures [26], and are currently one of the most abundant species in portions of the Illinois River [27]. More importantly, bigheaded carp have had documented negative impacts on aquatic ecosystems [25]. Silver carp, for example, can consume detritus and bacteria [28], and reduce the size and abundance of both phytoplankton and zooplankton [25,27,29,30,31]. As a result, studies have shown that populations of bigheaded carps can result in reduced condition and abundance of native planktivorous fishes [32,33], as well as a reduction in the abundance of adult sport fish that compete with bigheaded carps at the larval and juvenile stages [34]. Owing to their abundance, mobility and impacts on receiving ecosystems, a tremendous amount of resources have been devoted to the suppression, removal and eradication of bigheaded carp from the Illinois River for almost a decade [35]. While efforts to date have been successful at reducing population sizes by removing millions of kilograms of fish through contract harvesting and agency collections [35], populations of bigheaded carp still remain throughout the Illinois River, necessitating suppression efforts to prevent the expansion of the population.
3. Chicago Area Waterway System
Bigheaded carp have direct access into the Great Lakes Basin from the Mississippi Basin due to the presence of the Chicago Area Waterway System (CAWS). The CAWS is a series of human-created canals and channels, completed in the early 1900s, that breached the continental divide between the two basins. The CAWS was constructed for the purpose of removing both sewage effluent and stormwater runoff from Chicago, coupled with allowing the passage of commercial shipping vessels to move from the Great Lakes to the Gulf of Mexico [36,37,38]. At present, the only means of deterring the movement of bigheaded carp through the CAWS from the Mississippi basin into the Great Lakes (beyond extensive suppression/harvest efforts) is a trio of electric barriers near Romeoville, IL, USA, constructed in 2002 [37]. Silver carp and bighead carp currently are over 60 km from Lake Michigan [39], so the effectiveness of these electric barriers at stopping bigheaded carp from passing has not been tested explicitly. However, numerous investigations have documented that these barriers are subject to problems and deficiencies that could allow the passage of bigheaded carp. For example, Dettmers et al. [40] showed that a number of fish confined to cages did not become immobilized when dragged through the barrier alongside steel-hulled barges, Sparks et al. [41] showed that an adult common carp (Cyprinus carpio) outfitted with an acoustic telemetry tag was able to traverse the electric barrier (possibly associated with a passing barge), while Evans and Brouder [42] showed that fish can move through the electric barriers if they are trapped between barges. Parker et al. [43] used stationary sonar deployed within the barriers and showed small fish were able to move through the electric fields independent of the presence of barges. In addition, electricity loses effectiveness when applied to small fish [44], the electric barrier is prone to maintenance shut downs, floods and power loss [38], and no non-physical barrier is effective at stopping 100% of fish [45]. Mitigation measures have been proposed to redesign shipping locks to reduce the possibility of the exchange of invasive species between the Mississippi and the Great Lakes basins. The plan to modify locks will cost billions of dollars, take a decade or more to complete, and has yet to start, leaving the Great Lakes vulnerable to the passage of bigheaded carp through the CAWS for the foreseeable future [46]. The consequences, should bigheaded carps traverse the electric barriers and enter the Great Lakes, are not known and are difficult to predict [47,48,49,50,51]. The consensus is that an invasion of bigheaded carps would not be beneficial, however, making the containment of carp within the Mississippi Basin a critical priority for stakeholders. To supplement existing suppression efforts and increase redundancy and effectiveness at preventing movement or spread through the CAWS, additional barrier technologies would be valuable, ideally a technology that will permit the passage of barges and the downstream transport of wastewater through the CAWS.
Based on the above background, the goals of this review are to (1) outline the development of zones of carbon dioxide gas (CO2) as a non-physical barrier to deter the movement of invasive fishes, with a particular focus on two bigheaded carps: silver carp and bighead carp, and (2) highlight internal and external factors that mediate the performance of CO2 as a non-physical barrier, either increasing or decreasing its effectiveness as a barrier for invasive fish passage. When taken together, this review will not only share the origins of CO2 as a fish barrier, but also help researchers think about ways to improve performance and maximize the ability of different barrier technologies to deter the spread of invasive fishes.
4. Carbon Dioxide in the Atmosphere
The idea that CO2 could be used as a fish barrier is rooted in Earth’s history and the evolution of fishes. Billions of years ago, CO2 levels in the Earth’s atmosphere were high, and O2 was low [52]. As photosynthesizing bacteria on the planet became more abundant, the composition of gasses in the atmosphere changed such that the relative level of O2 increased and the level of CO2 declined [52,53]. This change in atmospheric oxygen was concurrent with metabolic evolution that increased reliance on oxidative phosphorylation that uses oxygen as a final electron acceptor resulting in more efficient metabolism, coupled with the production of CO2 as a waste product [52]. Thus, organisms developed the ability to sense environmental gasses, including CO2, and respond by either avoiding CO2-rich areas that might impair energetic processes, or possibly being drawn to CO2-rich areas if they provide an energetic advantage [53]. Bacterial and fungal pathogens, for example, can sense environmental CO2 associated with hosts and alter growth or life cycles to maximize virulence [53]. Hawkmoths (Manduca sexta, Lepidoptera: Sphingidae) use floral CO2 emissions to quantify food source profitability and the amount of nectar in flowers [54], while honey bees (Apis mallifera) actively fan their hives to remove CO2 wastes, and the number of individuals fanning correlates positively with CO2 levels inside the colony [55]. Carbon dioxide excreted by vertebrates is used by mosquitoes (Aedes spp.) as a signal of a potential host [56,57], while Drosophila will avoid CO2, likely as a signal that rotting fruit is a poor food source [58]. For many terrestrial vertebrates, CO2 is detected by chemoreceptors in the blood stream and brain stem to regulate breathing [59], while, more specifically, mammals detect of CO2 in the air with free nerve endings of the trigeminal system [60]. Together, concentrations of environmental CO2 can be a source of ecologically relevant information, and the ability to detect and respond to CO2 as a stimulus has persisted across kingdoms.
5. CO2 and Fish Physiology
Carbon dioxide has a pronounced effect on fishes, resulting in a host of physiological and behavioral responses when concentrations above species-specific set-points are experienced. Fish predominantly sense ambient CO2 using peripheral chemoreceptors, largely in the gills, that respond to CO2 tension in the water, not changes in pH; some evidence does exist for the presence of internal CO2 sensors, but the location of these sensors has not been well-defined [61]. When fish are placed in a high carbon dioxide environment, CO2 passively diffuses into the fish down its concentration gradient, and arterial CO2 equilibrates with environmental CO2 within minutes, resulting in an internal acidosis [62,63]. Over time, this pH imbalance is corrected as fish uptake HCO3− from the environment (in exchange for Cl−) and excrete H+ (in exchange for Na+) [63]. Owing to this influx of CO2, hypercarbic environments cause an elevation of the general stress response [64,65,66], a drop in blood pH [67], a loss of ions [68], and, ultimately, equilibrium loss and anesthesia (Stage 2 or Stage 3) [64,67,69,70]. At present, the exact mechanism(s) responsible for the loss of equilibrium and the anesthetic impacts of carbon dioxide have not been well defined, but are believed to result from the movement of CO2 across the blood-brain barrier, which alters brain pH and an impairs brain electrical activity [71,72]; additions of H+ or HCO3− alone will not result in anesthesia for fish [71]. In addition to these physiological changes, studies have documented behavioral changes exhibited by fish in high CO2 environments including hyperventilation, coupled with a reduction in heart rate, likely to facilitate CO2 excretion [61]. Together, exposure to water with elevated concentrations of CO2 has been shown to result in both physiological and behavioral changes to fish.
6. CO2 and Fish Behavior
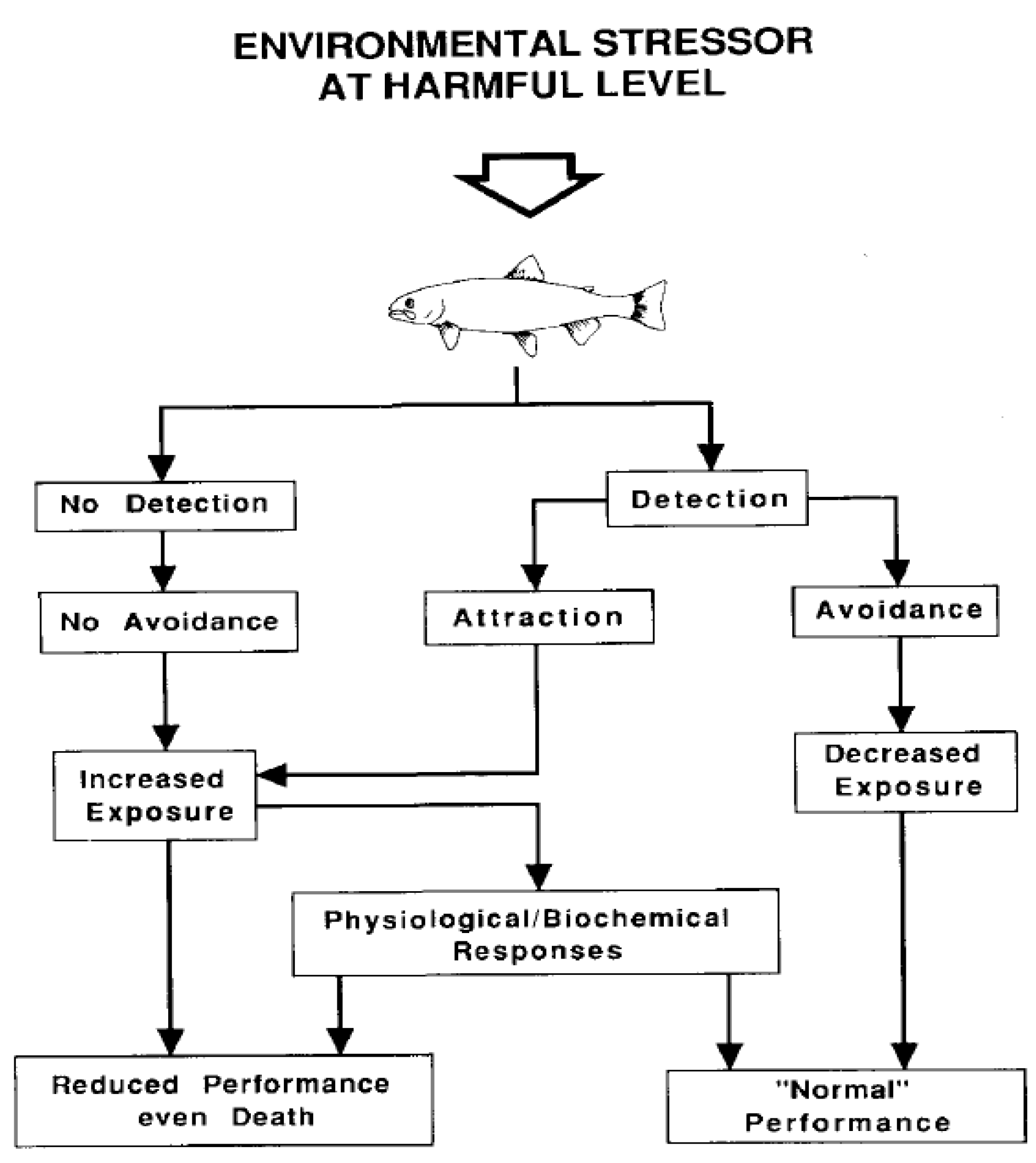
A number of past studies have highlighted the propensity for fish to voluntarily swim away from areas of high carbon dioxide, laying the foundation for the use of CO2 as a fish deterrent. Avoidance reactions are considered the first line of defense for fish that encounter adverse stimuli, and poor water quality can quickly induce a behavioral response that causes fish to depart an area and seek out improved water, presumably to minimize energetic costs [73,74] (Figure 1).
Over a century ago, Shelford and Allee [75] designed a simple experiment to observe the behavior of nine fish species when placed individually in a raceway containing a gradient of CO2, ranging from approximately 2–88 cubic centimeters of CO2 per liter of water. Shelford and Allee [75] showed that, upon entering zones of elevated CO2, some fish started surface ventilations, while others displayed a coughing or yawning reaction coupled with increased ventilation rates. In addition, Shelford and Allee [75] reported that some fish would enter the area of high CO2, stop suddenly, and then move backwards as if they had “encountered a sheet-rubber wall”, and all fish spent less time in areas of high CO2 relative to areas with lower CO2. Powers and Clark [76] used a laboratory gradient tank design similar to Shelford and Allee [75] and showed that both brook trout (Salvelinus f. frontinalis) and rainbow trout (Salmo gairdnerii iridus) also avoided water that had received “very slight” additions of CO2 (a drop of approximately 0.4 pH units). This pattern was also confirmed by Collins [77] who showed that individual alewife (Polumbus pseudoharengus) and glut herring (P. aestivalis) (likely river herring) migrating upriver both avoided water with elevated CO2 that exceeded 0.3 ppm, independent of pH changes. Bishai [78] showed that juvenile brown trout (Salmo trutta) and Atlantic salmon (Salmo salar) demonstrated a stronger avoidance response to a pH change caused by CO2 relative to a pH change caused by hydrochloric acid. Jones et al. [79] noted that individual arctic char (Salvelinus aplinus) will avoid concentrations of CO2 that exceed 50 µmol/L, Ross et al. [80] showed that brook trout and blacknose dace (Rhinichthys atratulus) would avoid water with ≥2% CO2, while Clingerman et al. [81] reported that intentional elevations of CO2 to 60 mg/L in an aquaculture would induce avoidance behavior in groups of rainbow trout (Oncorhynchus mykiss), thereby facilitating harvest and collection in recirculating tanks. Finally, both Bernier and Randall [64], as well as Yoshikawa [82], revealed that rainbow trout exhibited a “violent” struggle upon being exposed to water maintained at 36–350 mmHg CO2, while Clingerman et al. [81] indicated that rainbow trout in aquaculture tanks demonstrated “chaotic” swimming when CO2 levels were increased to 35–60 mg/L. Thus, for over 100 years, studies have documented that many fish species will avoid areas of elevated CO2 once a threshold is reached, providing the proof of concept that CO2 could be a potential non-physical barrier for invasive fishes.
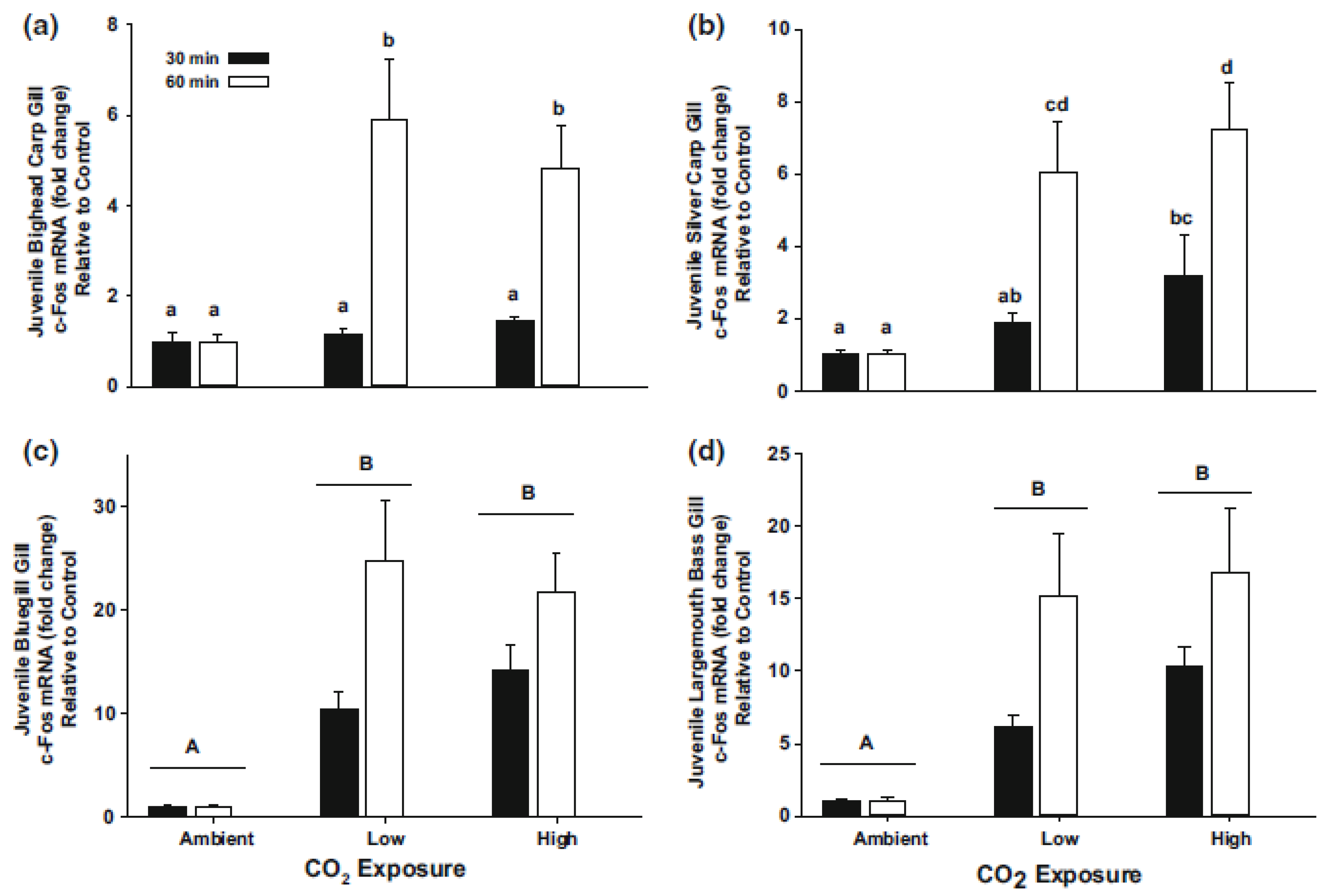
Despite suggestions from past work that CO2 could induce avoidance behaviors and act a barrier to the movement of bigheaded carp, a key unknown was the threshold CO2 level that should be targeted to induce avoidance. While the response of fishes to high concentrations of CO2 when applied as an anesthetic appeared to be consistent [67,83], and the physiological responses of fishes to general hypercarbia had been well-defined [63], relatively less was known about the thresholds or “inflection points” that cause the onset of disturbances (i.e., a dose-response curve), and if those threshold concentrations were consistent across species and life stages. For example, Ross et al. [80] exposed book trout, slimy sculpin (Cottus cognatus) and blacknose dace to four levels of CO2 (0%, 1.4%, 2.8% and 5.1%) for either one or 24 h and noted differences in physiological responses both across species and across exposure durations, suggesting species-specific responses to CO2 exposure. To address this need and define concentrations that induced onset of disturbances, Kates et al. [66] exposed bluegill (Lepomis macrochirus), largemouth bass (Micropterus salmoides), silver carp (>450 mm) and bighead carp (>700 mm) to two different concentrations of CO2 (30 mg/L and 70 mg/L) for three hours and showed that, 30 mg/L CO2 (approximately 2000 µatm CO2) had minimal physiological or behavioral impacts, but a three hour exposure to 70 mg/L CO2 (approximately 50,000 µatm CO2) resulted in a drop in ventilation rates, and an increase in irregular behaviors such as erratic swimming, twitching and escape attempts for silver carp and bighead carp [66]. One of the challenges with the study by Kates et al. [66], however, was that adult bigheaded carp were used, which provided little evidence in support of how small fish, those presumably less vulnerable to the existing electric barriers in Romeoville, IL, would respond to CO2. In an effort to better define the allometric response of fish to CO2 exposure, Dennis et al. [84] exposed larval and juvenile (73 mm) silver carp and bighead carp to 120 mg/L CO2 (approximately 42,000 µatm CO2) for 30 and 60 min. Results from this study were similar to previous work with adult fish, in that exposing larval silver carp and bighead carp to 42,000 µatm CO2 for 30 min resulted in an increase in the mRNA coding for genes associated with the stress response (Hsp70 and c-fos) [84] (Figure 2).
Thus, when results from these two studies are taken together, data suggest that thresholds of approximately 42,000 µatm CO2 (70–120 mg/L) induce a suite of physiological and behavioral responses for a range of sizes of silver and bighead carp consistent with stress or discomfort, providing a target in the development of a non-physical barrier for fish.
7. CO2 and Physiological Responses
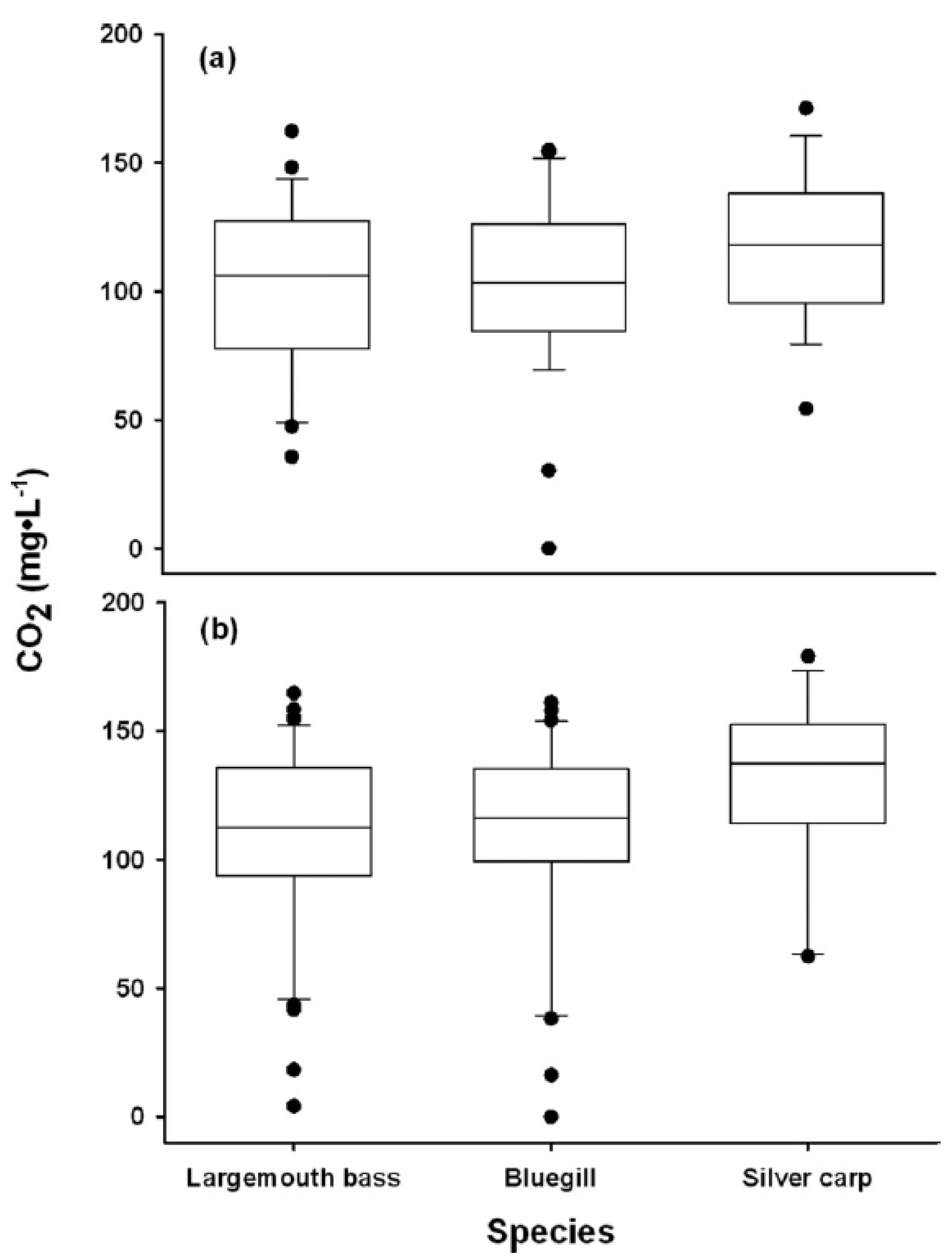
Following the identification of putative thresholds that would induce behavioral and physiological disturbances, studies on CO2 barriers shifted to quantify aspects of avoidance (Figure 1). Despite the research mentioned above that indicated a pattern of fish avoiding zones of elevated CO2, there were suggestions in the literature that avoidance responses may be variable across species. Ross et al. [80], for example, showed that individual slimy sculpin did not avoid zones of elevated CO2 and preferred to rest in place when confronted with hypercarbia, while Summerfelt and Lewis [85] noted that CO2 concentrations from 3.0–9.7 mg/L did not repel green sunfish (Lepomis cyanellus) in a graded laboratory tank. Early work with CO2 avoidance and bigheaded carp was conducted by Kates et al. [66] who used a “shuttle-box” apparatus in a laboratory to show that individual adult silver carp (460 mm) would voluntarily swim away from CO2 once concentrations were elevated to approximately 120 mg/L CO2, although there was considerable variation around this mean value (Figure 3) (Table 1).
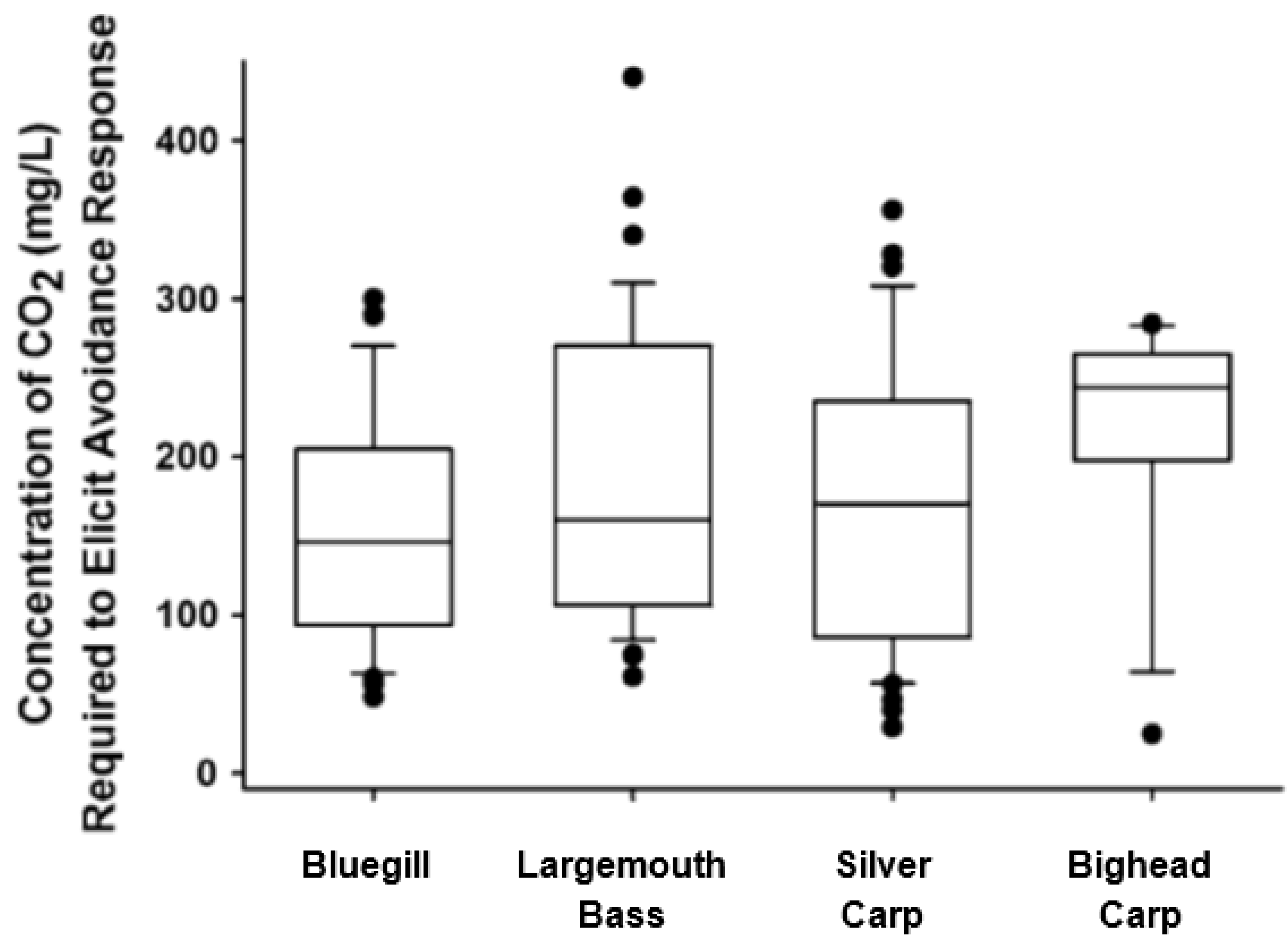
Dennis et al. [84] later used juvenile silver carp and bighead carp (67 mm and 71 mm, respectively) and the shuttle-box apparatus, and, again, showed that individual fish would voluntarily swim away from zones of elevated CO2. The concentration of CO2 required to induce avoidance in this series of tests averaged approximately 180–220 mg/L CO2, and, again, the variance around the mean was considerable (Figure 4). The success of these laboratory trials led to work at larger settings, including Donaldson et al. [86] who showed that a number of fish species, including silver carp and bigheaded carp, released into a 4000 m3 outdoor pond in groups of 5–10 avoided zones of elevated CO2 elevated to approximately 30,000 µatm (60 mg/L), and Cupp et al. [87] who used a two-channel, outdoor raceway (approximately 60 m3) with flowing water and showed that CO2 levels of approximately 30,000–40,000 µatm (~75 mg/L) would deter the movement of both silver and bigheaded carp (278 mm and 212 mm, respectively) when tested in groups of 10 (Table 1). Cupp et al. [88] showed that CO2 deployed at the mouth of an outflow structure draining a backwater lake could reduce the abundance of shoals of mixed fish species by 70–95% at low water flows once a target threshold of 100 mg/L was reached. Finally, Hasler et al. [89] worked in a 12 m long indoor swim flume and showed that bighead carp (145 mm) in shoals of 3 would avoid CO2 in water flowing at approximately 15 cm/seconds (equivalent to 1 body length per second), and CO2 levels in this study were approximately 190,000 µatm. When considered together, these studies used a number of environments (indoor, outdoor, static water, flowing water) to demonstrate that a range of sizes of invasive silver and bighead carp, including small fish presumed to be less vulnerable to electricity, would voluntarily swim away from zones of elevated CO2 once a threshold of approximately 70,000 µatm (100 mg/L) was reached, providing support for the use of CO2 as a non-physical barrier.
8. CO2 as a Potential Fish Barrier
When the general anesthetic properties of CO2 exposure [67] were combined with results from field and lab avoidance trials, there was a considerable amount of evidence to suggest that zones of elevated CO2 could deter the spread of invasive bigheaded carps. More specifically, a CO2 barrier could be deployed in one of two different ways. First, CO2 could be deployed as a “fence” or wall with the goal of inducing avoidance behaviors in fishes, exploiting the fact that fish voluntarily swim away from areas of high CO2 once a threshold had been reached. For example, CO2 could be used to confine carp in backwater areas [90] preventing access to turbulent, high-velocity water flowing river environments used for spawning [91], or at a choke-point in a river (e.g., shipping lock) to stop movement. A CO2 barrier deployed in this way could be temporary (e.g., deployed only during summer or during harvest), or for longer periods of time. Secondly, zones of CO2 could be deployed to intentionally induce equilibrium loss for fish, taking advantage of the ability of fish to tolerate CO2 as an anesthetic. Again, an application of this kind could be temporary (i.e., deployed at specific times of the year) or longer-term (e.g., added to a shipping lock) [46].
9. Questions from Avoidance Data
While the concept of using CO2 as a barrier based on either avoidance or a tolerance has support from a number of studies, there were several puzzling trends in the data, which generated questions and presented challenges related to possible deployment. For example, shuttle-box work by Kates et al. [66] showed that avoidance thresholds for individual adult silver carp and bighead carp spanned from approximately 50 mg/L to 160 mg/L (Figure 3) (Table 1). Subsequent shuttle-box work by Dennis et al. [84] with juvenile fishes, showed that avoidance thresholds ranged 6-fold, from approximately 50–300 mg/L (Figure 4). This variation in avoidance is further complicated by work from outdoor ponds showing avoidance occurred for groups of bighead carp, but CO2 never exceed 60–75 mg/L [86,87]. Questions therefore arose related to the source of this variation (i.e., Is there inter-individual variation? What is the nature of the differing test environments? Is this variation inherent in how animals respond to zones of CO2?), the potential for inter-individual differences in tolerance, and the effectiveness/consistency of CO2 across time periods or environments. Thus, it was difficult to make recommendations to managers on target thresholds necessary to achieve an effective CO2 barrier, or to predict possible changes in barrier effectiveness, without a more thorough understanding of the response of fish to CO2 barriers. A series of studies were therefore conducted to quantify endogenous and exogenous factors that influenced the avoidance and tolerance of fishes to elevated CO2 in hopes of refining this technology, providing stronger, more definitive recommendations to managers on target thresholds for CO2 barriers, and improving the likelihood of long-term performance of CO2 as a non-physical barrier.
10. Factors Influencing the Avoidance of CO2
Several different endogenous and exogenous factors have the potential to influence the avoidance response of CO2 in the context of a non-physical barrier (Table 2). For instance, in recent years, it has become apparent that fish consistently differ from each other in behavior, often termed “personality”; some individuals are more bold than others, some are more active, and some are more likely to explore novel areas [92]. Invasive round goby at the leading edge of their range, for example, were shown to be more bold and willing to explore novel areas than individuals from established, core populations [93], and it is therefore plausible that individual differences in personality could be manifesting in inter-individual differences to CO2 avoidance [94] (Figure 3 and Figure 4). More importantly, personality differences covary with characteristics such as the response to stressors, life span and growth rate through the pace-of-life continuum [92]. Therefore, if the response to an environmental stress and avoidance thresholds are mediated through behavior (e.g., proactive vs. reactive coping styles [92,95]), due to links between personality, life history and fitness [96], if target levels for an avoidance barrier are too low and fish of a particular behavioral type are able to pass, this could translate to population-level shifts in phenotypes, possibly changing avoidance thresholds for a population. In exploration of this concept, Tucker et al. [97] showed that aspects of personality (e.g., activity and boldness) did not influence CO2 avoidance in individual bluegill, with fish of all personality types avoiding CO2 at a threshold of approximately 67,000 µatm. In addition, Tucker and Suski [98] showed that social personality in bluegill (e.g., sociability, clustering with conspecifics and conspecific aggression) also did not influence CO2 avoidance thresholds or the order that fish avoided CO2. Related to this, past work has shown that food deprivation can alter the behavior of fish through plastic or flexible changes, with animals deprived of food taking more risks and becoming more active, likely as they search for food [99,100]. Interestingly, Suski et al. [101] showed that nine days of food deprivation did not influence CO2 avoidance thresholds for individual largemouth bass; fish that had been fed and fish that had been deprived of food, both avoided high CO2 at thresholds of approximately 70,000 µatm. However, Tucker et al. [97] showed that CO2 avoidance in individual largemouth bass was influenced by artificial activation of the stress axis as fish that received an intraperitoneal injection of cortisol (hydrocortisone 21-hemisuccinate) required 45% more CO2 to induce avoidance behavior relative to fish that did not receive an artificial elevation of the stress axis. Many initial studies of CO2 avoidance [66,84] were conducted on individual fish, but shoals have a number of benefits for fish including predator vigilance and food detection, resulting in a calming effect and a reduced response to environmental stressors [102]. Tucker and Suski [98] showed pronounced differences in CO2 avoidance thresholds for individual fish relative to shoals, with groups of bluegill choosing to swim away from CO2 at significantly lower thresholds than individual bluegill; interestingly Tucker et al. [97] also showed that shuttling thresholds were not repeatable within individuals. Allometry is known to influence a number of characteristics of fish including metabolism and survival, but intraspecific differences in CO2 avoidance thresholds across size categories is not clear. When avoidance thresholds for small and large bighead carp are compared across Kates et al. [66] and Dennis et al. [103], small fish appear to require higher CO2 thresholds to induce avoidance. These results, however, were obtained in separate studies, not in a single investigation, so inter-study differences may have been a complicating factor. The quantity of CO2 necessary to induce avoidance in round goby (Neogobius melanostomus) [104], silver carp and bighead carp [105] correlated positively with water temperature (range from 5–25 °C), such that more CO2 was required to induce avoidance at high temperatures for all three species tested. Note that both Cupp et al. [104] and Tix et al. [105] did not acclimate fish at test temperatures for a period of two to three weeks as is common [106,107], with holding times listed at two to six days, which could have influenced these results. Together, a number of factors have been shown to influence the threshold of CO2 required to induce avoidance behaviors, which have implications for the application of CO2 as an avoidance barrier to deter the movement of invasive fishes (Table 2).
11. Factors Influencing CO2 Tolerance
Similar to work with avoidance, a number of studies have been carried out to quantify inter-individual differences in CO2 tolerance, as well as potential mechanisms for any differences (Table 3). Importantly, Hasler et al. [108] showed that, for largemouth bass, CO2 tolerance not only varied across individuals, with some fish losing equilibrium in high CO2 sooner than others, but also that the individual response to high CO2 was repeatable within individuals, suggesting potential for this to be a heritable trait that can be acted upon by natural selection [109]. In general, tolerance to CO2 is a function of the interaction of exposure time × concentration, further mediated by temperature [64,69,70,110,111,112]. More specifically, a brief exposure to a high concentration of ambient CO2, or an extended exposure to lower concentrations of CO2, will both result in equilibrium loss, provided that the concentration of ambient CO2 is sufficient to passively diffuse into the bloodstream of the fish [64,70,110,112]. Owing to reduced respiratory and metabolic rates at low temperatures, fish typically require additional time at lower temperatures before anesthetic effects are realized relative to high temperatures [112]. Indeed, this has been demonstrated for CO2 as both Fish [69] and Gelwicks et al. [110] used study designs where individual fish were transferred to containers of CO2 at a target concentration and showed decreased time to equilibrium loss at high temperatures, suggesting that fish are more sensitive to CO2 at high temperatures. Interestingly, both Cupp et al. [104], and Tix et al. [105], showed that, when CO2 was continually added to a test tank, round gobies [104], silver carp and bighead carp [105] all required higher concentrations of CO2 before equilibrium loss occurred when animals were at high temperatures relative to low temperatures, suggesting that fish were more tolerant to CO2 at higher temperatures. There are three potential explanations for the discrepancies across these studies. Firstly, differences across studies could be due to experimental animals, as Fish [69] and Gelwicks et al. [110] worked with salmonids, while Tix et al. [104] and Cupp et al. [105] used round goby and bigheaded carp. Secondly, Tix et al. [105] and Cupp et al. [104] applied CO2 to fish continuously until equilibrium loss occurred, while Fish [69] and Gelwicks et al. [110] pre-treated tanks of water with CO2 to a set point and added fish. Finally, both Cupp et al. [104] and Tix et al. [105] did not acclimate animals to each test temperature for extended periods of time, and, rather, animals were first held at 12 °C and then transferred to the test temperatures for 24–144 h prior to testing, which may have influenced their response to CO2 exposure [106,107]. Clingerman [81] showed that, when CO2 level was held constant, large rainbow trout were more likely to lose equilibrium than small rainbow trout in aquaculture tanks, suggesting an increased tolerance for smaller fish. Tucker et al. [97] showed that aspects of personality (e.g., activity and boldness) did not influence CO2 tolerance in bluegill, with fish of all personality types requiring similar durations of time to induce equilibrium loss when exposed to 123,000 µatm. Hasler et al. [108] showed that tolerance to CO2 was influenced by the metabolic phenotype of largemouth bass, and fish with higher anaerobic performance, quantified as time to become exhausted when burst swimming, required less time to lose equilibrium when exposed to high CO2, and also that aerobic aspects of metabolic phenotype (i.e., standard metabolic rate, aerobic scope) did not influence tolerance to carbon dioxide. Suski et al. [101] showed that largemouth bass that had been deprived of food for 14 days required 25% longer exposure to high CO2, relative to fish that had been fed over this 14 day period, thereby demonstrating an increased tolerance to CO2 from food deprivation. Together, tolerance to high CO2 can vary due to a number of endogenous and exogenous factors and should be considered should CO2 be deployed to deter the movement of invasive fishes (Table 3).
12. Management Implications
There are a number of potential non-physical barriers that can be deployed to prevent the spread of invasive fishes, including bubble screens, sound or electricity, each with particular strengths and weaknesses [45]. A non-physical barrier that uses zones of elevated carbon dioxide to deter fish movements has a number an advantageous as a chemical control tool relative to other technologies as it has few human health concerns, can be applied in a carbon neutral fashion using repurposed CO2 (i.e., harvesting waste CO2 destined to be released into the atmosphere), is relatively inexpensive and readily available, can be deployed with relatively little infrastructure, and residual CO2 does not persist in the environment [113]. Carbon dioxide was recently registered with the United States Environmental Protection Agency (USEPA) as a pesticide for use as a deterrent of bigheaded carp under the name Carbon Dioxide—Carp (EPA Registration Number 6704-95). Dennis et al. [103] held largemouth bass at 21,000 µatm (13 mg/L) CO2 for almost two months and showed no decline in avoidance thresholds, suggesting that acclimation to the presence of high CO2 is not likely. Most important, the avoidance response of fishes to environmental CO2 appears to be canalized, demonstrated by virtually all fish species tested, while CO2 tolerance is repeatable and consistent [108], giving CO2 a number of advantages as a non-physical fish barrier as a tool to deter invasive fishes.
Results from the studies listed above have a number of implications for the deployment of CO2 as a non-physical barrier and can be used to minimize the likelihood of unintentional fish passage, while also helping minimize waste CO2 and reduce deployment costs. For example, for a CO2 barrier deployed with the intention of causing avoidance, it is important for managers to consider the context in which the barrier is deployed. More specifically, although not repeatable within individuals, avoidance of CO2 has been shown to be consistent across virtually all species tested when CO2 pressures reach approximately 30,000–60,000 µatm (60–100 mg/L). However, avoidance thresholds will likely be lower for fish in shoals (rather than individual fish) but will increase if fish are experiencing stress (independent of food availability), such as chronic hypoxia or environmental pollution. Finally, studies suggest that higher concentrations of CO2 may be required to induce avoidance at warmer water temperatures (summer) relative to cooler conditions (Table 2). It should be noted that Schneider et al. [114] showed that CO2 did not impair either the burst or sprint swimming performance of largemouth bass until thresholds of 100,000 µatm were reached (approximately 150 mg/L), well in excess of thresholds required to induce avoidance, suggesting that, if fish choose to challenge a CO2 barrier and burst through it, the barrier will likely not impair swimming performance. When considered together, a number of factors should be considered to ensure maximum effectiveness should CO2 be used in the field to deter the movement of invasive fishes (Table 2).
If a CO2 barrier is deployed with the intent of stopping fish via equilibrium loss (tolerance), aspects of individual fish need to be considered as these factors can influence effectiveness. At present, the relationship between exposure time × CO2 concentrations that results in equilibrium loss for most species has not been defined, so these data would need to be collected to help guide management targets, and owing to individual variation in the loss of equilibrium time for fish [108], a large number of fish would need to be assessed to quantify a range of equilibrium loss times. In general, however, small fish, and individuals that had been deprived of food, would be expected to have improved tolerance in high CO2 relative to larger, well-fed individuals. The role of environmental temperature has not been clearly defined, but studies suggest that a longer exposure time may be required at lower water temperatures and in periods of low food availability (e.g., winter) (Table 3).
13. Future Work
At present, there are five areas that should be the focus of future studies to improve the performance and efficacy of CO2 as a non-physical barrier. Firstly, additional work should focus on defining differences in both avoidance and tolerance thresholds across fish of different sizes; this is particularly important given the possibility that electricity as a barrier may lose effectiveness against small fish [43]. Currently, work that quantifies avoidance and tolerance thresholds across a range of sizes of fishes, within a single study with consistent methods, has not occurred. Owing to the likelihood that a CO2 barrier would be encountered by fish of a range of sizes, the ability to confidently predict the response of different sized individuals to either a tolerance or avoidance application of CO2 is critical. Secondly, the exact parameters of the time × concentration interaction to induce equilibrium loss associated with a tolerance-focused barrier have not been defined extensively, and would need to be parameterized across target species before tolerance barriers could be developed and/or implemented. Ideally, this work would be conducted across a range of temperatures. Thirdly, work should be conducted that pairs CO2 barriers with additional stimuli (e.g., deploy a sound barrier and CO2 barrier concurrently as in Ruebush et al. [115], or use CO2 as part of a bubble curtain rather than compressed air). No non-physical barrier is 100% effective at stopping all fish [45], but a CO2 barrier paired with a second stimulus (e.g., light or sound barriers) could synergistically improve the overall effectiveness of each barrier, increasing the potential to deter invasive fishes across a range of conditions. Penultimately, efforts need to occur to quantify the logistics of CO2 deployment, including cost estimates, deployment feasibility and infrastructure requirements to assist with future planning efforts. The design for deploying a CO2 barrier will vary across sites and situations, but efforts to share costs and strategies across successful applications will help improve deployment efficiency and ensure success across locations. Finally, owing to the unavoidable reductions in pH that occur with zones of elevated CO2, work should continue to quantify the environmental impacts [116,117], consequences for non-target organisms (e.g., mussels [118,119]; native fishes [120,121,122]; crayfish [123]) and strategies for CO2 off-gassing. Work to both mitigate CO2 applications, coupled with efforts to predict possible impacts to non-target organisms or the receiving environment, will help improve the likelihood of a successful application. Together, work to address these 5 concerns will not only help improve the effectiveness of CO2 as a non-physical barrier, but also will help minimize unintended environmental consequences and improve the efficiency of CO2 as a tool to deter invasive fishes.
14. Conclusions
Invasive species represent a significant threat to global biodiversity, and models suggest that the rate of introduction of invasive species will likely accelerate in the future [16]. For fishes in North America, bigheaded carp represent a current threat to the Mississippi ecosystem, and there is potential for them to gain access to waterbodies in the eastern portion of the continent should they pass through the Chicago Area Waterway System (CAWS) into the Great Lakes basin. Carbon dioxide (CO2) is a naturally occurring compound that provides ecologically-relevant information to a host of taxa. A number of different studies, conducted across a range of conditions, have demonstrated that zones of elevated carbon dioxide gas can be an effective non-physical barrier to deter the spread of invasive fishes. More specifically, fish will voluntarily swim away from zones of high CO2 once a target threshold has been reached, or else equilibrium loss will occur due to the anesthetic properties of CO2, providing two different mechanisms by which carbon dioxide can deter fish movement. This response has been documented for a number of taxonomically diverse species of fish, and also across a range of sizes spanning from larvae to adults. In addition, unlike physical barriers, a CO2 barrier can be deployed without requiring the construction of permanent structures that can modify water flow or boat traffic. Several internal and external factors can influence the response of fishes to CO2, making them more effective, or less effective (e.g., fish experiencing stress will require additional CO2 to induce avoidance relative to non-stressed individuals; shoals of fish require less CO2 to induce avoidance relative to solitary individuals) and need to be considered when defining target thresholds should CO2 be deployed in the field. Additional studies to define effective deployment strategies at large scales, cost, and impacts to the receiving environment should continue as CO2 barriers grow in popularity and field applications. Work is currently ongoing to develop other non-physical barriers to deter invasive fishes (e.g., sound, electricity, strobe lights), and the lessons learned and experiences described here from CO2 can serve as potential considerations to refine the application of other barrier technologies to increase their effectiveness. Together, with continued exploration and testing, it is hoped that barrier technologies can be further developed to prevent the spread of invasive fishes and protect freshwater biodiversity.
Funding
This research was funded by the Illinois Department of Natural Resources, and the United States Geological Survey, through funds provided by the United States Environmental Protection Agency’s Great Lakes Restoration Initiative.
Acknowledgments
A number of individuals have contributed to field and laboratory work that have generated the data contained in this review, including Caleb Hasler, Jennifer Jeffrey, Kelly Hannan, John Tix, Eric Schneider, Emi Tucker, Madison Philipp, Ian Bouyoucos, Christa Woodley, Cody Sullivan, Jason Romine, Dave Smith, Steve Midway, Aaron Cupp, Jon Amberg, Mark Gaikowski, Clark Dennis, Matt Noatch, Dan Kates, Shivani Adhikari, Michael Donaldson, and Adam Wright. Jake Wolf fish hatchery provided fish for experiments, and staff at the Illinois River Biological Station facilitated the collection of carp. Caleb Hasler provided comments on an early version of this review.
Conflicts of Interest
The author declares no conflict of interest.
References
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological Invasions: Recommendations for US Policy and Management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef] [Green Version]
- Britton, J.R.; Davies, G.D.; Harrod, C. Trophic Interactions and Consequent Impacts of the Invasive Fish Pseudorasbora Parva in a Native Aquatic Foodweb: A Field Investigation in the UK. Boil. Invasions 2009, 12, 1533–1542. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current Knowledge on Non-Native Freshwater Fish Introductions. J. Fish Boil. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. Quantifying Threats to Imperiled Species in the United States. BioScience 1998, 48, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the Environmental and Economic Costs Associated with Alien-Invasive Species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Pejchar, L.; Mooney, H.A. Invasive Species, Ecosystem Services and Human Well-Being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef]
- Strayer, D.L.; Dudgeon, D. Freshwater Biodiversity Conservation: Recent Progress and Future Challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. Landscape Sustainability Science: Ecosystem Services and Human Well-Being in Changing Landscapes. Landsc. Ecol. 2013, 28, 999–1023. [Google Scholar] [CrossRef]
- Jenkins, M. Prospects for Biodiversity. Science 2003, 302, 1175–1177. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging Threats and Persistent Conservation Challenges for Freshwater Biodiversity. Boil. Rev. 2018, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater Biodiversity: Importance, Threats, Status and Conservation Challenges. Boil. Rev. 2005, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Jelks, H.L.; Walsh, S.J.; Burkhead, N.M.; Contreras-Balderas, S.; Díaz-Pardo, E.; Hendrickson, D.A.; Lyons, J.; Mandrak, N.E.; McCormick, F.; Nelson, J.S.; et al. Conservation Status of Imperiled North American Freshwater and Diadromous Fishes. Fisheries 2008, 33, 372–407. [Google Scholar] [CrossRef]
- Burkhead, N.M. Extinction Rates in North American Freshwater Fishes, 1900–2010. BioScience 2012, 62, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.; et al. Consequences of Changing Biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Sardain, A.; Sardain, E.; Leung, B. Global Forecasts of Shipping Traffic and Biological Invasions to 2050. Nat. Sustain. 2019, 2, 274–282. [Google Scholar] [CrossRef]
- Clout, M.N.; Veitch, C.R. Turning the tide of biological invasion: the potential for eradicating invasive species. In Turning the Tide: The Eradication of Invasive Species; IUCN SSC Invasive Species Specialist Group: Gland, Switzerland; Cambridge, UK, 2002; pp. 1–3. Available online: http://www.issg.org/pdf/publications/turning_the_tide.pdf (accessed on 1 June 2020).
- Simberloff, D. Eradication—Preventing Invasions at the Outset. Weed Sci. 2003, 51, 247–253. [Google Scholar] [CrossRef]
- Simberloff, D. We can eliminate invasions or live with them. Successful management projects. In Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems; Langor, D., Sweeney, J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 149–157. ISBN 978-1-4020-9680-8. [Google Scholar]
- Simberloff, D. Eradication: Pipe dream or real option? In Plant Invasions in Protected Areas; Foxcroft, L.C., Pyšek, P., Richardson, D.M., Genovesi, P., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 549–559. ISBN 978-94-007-7750-7. [Google Scholar]
- Leung, B.; Lodge, D.M.; Finnoff, D.; Shogren, J.F.; Lewis, M.A.; Lamberti, G. An Ounce of Prevention or a Pound of Cure: Bioeconomic Risk Analysis of Invasive Species. Proc. R. Soc. B Boil. Sci. 2002, 269, 2407–2413. [Google Scholar] [CrossRef] [Green Version]
- Finnoff, D.; Shogren, J.F.; Leung, B.; Lodge, D. Take a Risk: Preferring Prevention over Control of Biological Invaders. Ecol. Econ. 2007, 62, 216–222. [Google Scholar] [CrossRef]
- Zanden, M.J.V.; Olden, J.D. A Management Framework for Preventing the Secondary Spread of Aquatic Invasive Species. Can. J. Fish. Aquat. Sci. 2008, 65, 1512–1522. [Google Scholar] [CrossRef]
- Kocovsky, P.M.; Chapman, D.C.; Qian, S. “Asian Carp” Is Societally and Scientifically Problematic. Let’s Replace It. Fisheries 2018, 43, 311–316. [Google Scholar] [CrossRef]
- Kolar, C.S.; Chapman, D.C.; Courtenay, W.R.J.; Housel, C.M.; Williams, J.D.; Jennings, D.P. Asian Carps of the Genus Hypophthalmichthys (Pisces, Cyprinidae)—A Biological Synopsis and Environmental Risk Assessment; American Fisheries Society Special Publication: Bethesda, MD, USA, 2007; Volume 33, ISBN 978-1-888569-79-7. [Google Scholar]
- Whitledge, G.W.; Knights, B.; Vallazza, J.; Larson, J.; Weber, M.J.; Lamer, J.T.; Phelps, Q.E.; Norman, J.D. Identification of Bighead Carp and Silver Carp Early-Life Environments and Inferring Lock and Dam 19 Passage in the Upper Mississippi River: Insights from Otolith Chemistry. Boil. Invasions 2018, 21, 1007–1020. [Google Scholar] [CrossRef] [Green Version]
- Sass, G.G.; Hinz, C.; Erickson, A.C.; McClelland, N.N.; McClelland, M.A.; Epifanio, J.M. Invasive Bighead and Silver Carp Effects on Zooplankton Communities in the Illinois River, Illinois, USA. J. Great Lakes Res. 2014, 40, 911–921. [Google Scholar] [CrossRef]
- Kuznetsov, Y.A. Consumption of Bacteria by the Silver Carp (Hypophthalmichthys molitrix). J. Ichthyol. 1977, 17, 398–403. [Google Scholar]
- Fukushima, M.; Takamura, N.; Sun, L.; Nakagawa, M.; Matsushige, K.; Xie, P. Changes in the Plankton Community Following Introduction of Filter-Feeding Planktivorous Fish. Freshw. Boil. 1999, 42, 719–735. [Google Scholar] [CrossRef]
- Laws, E.A.; Weisburd, R. Use of Silver Carp to Control Algal Biomass in Aquaculture Ponds. Progress. Fish-Culturist 1990, 52, 1–8. [Google Scholar] [CrossRef]
- Lieberman, D.M. Use of Silver Carp (Hypophthalmichthys molotrix) and Bighead Carp (Aristichthys nobilis) for Algae Control in a Small Pond: Changes in Water Quality. J. Freshw. Ecol. 1996, 11, 391–397. [Google Scholar] [CrossRef]
- Irons, K.S.; Sass, G.G.; McClelland, M.A.; Stafford, J.D. Reduced Condition Factor of Two Native Fish Species Coincident with Invasion of Non-Native Asian Carps in the Illinois River, USA Is This Evidence for Competition and Reduced Fitness? J. Fish Boil. 2007, 71, 258–273. [Google Scholar] [CrossRef]
- Pendleton, R.M.; Schwinghamer, C.; Solomon, L.E.; Casper, A.F. Competition among River Planktivores: Are Native Planktivores Still Fewer and Skinnier in Response to the Silver Carp Invasion? Environ. Boil. Fishes 2017, 100, 1213–1222. [Google Scholar] [CrossRef]
- Chick, J.H.; Gibson-Reinemer, D.K.; Soeken-Gittinger, L.; Casper, A.F. Invasive Silver Carp is Empirically Linked to Declines of Native Sport Fish in the Upper Mississippi River System. Boil. Invasions 2019, 22, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Asian Carp Regional Coordinating Committee. Asian Carp Monitoring and Response Plan. 2018. Available online: https://www.asiancarp.us/Documents/MRP2018.pdf (accessed on 1 June 2020).
- Hill, L. The Chicago River: A Natural and Unnatural History; Lake Claremont Press: Chicago, IL, USA, 2000. [Google Scholar]
- Moy, P.B.; Polls, I.; Dettmers, J.M. The Chicago Sanitary and Ship Canal aquatic nuisance species dispersal barrier. In Invasive Asian Carps in North America, American Fisheries Society Symposium; Chapman, D.C., Hoff, M.H., Eds.; American Fisheries Society: Bethesda, MD, USA, 2010; Volume 74, pp. 121–137. [Google Scholar]
- Rasmussen, J.L.; Regier, H.A.; Sparks, R.E.; Taylor, W.W. Dividing the Waters: The Case for Hydrologic Separation of the North American Great Lakes and Mississippi River Basins. J. Great Lakes Res. 2011, 37, 588–592. [Google Scholar] [CrossRef]
- Asian Carp Regional Coordinating Committee. Asian Carp Action Plan. 2020. Available online: https://www.asiancarp.us/Documents/2020-Action-Plan.pdf (accessed on 1 June 2020).
- Dettmers, J.M.; Boisvert, B.A.; Barkley, T.; Sparks, R.E. Potential Impact of Steel-Hulled Barges on Movement of Fish across an Electric Barrier to Prevent the Entry of Invasive Carp into Lake Michigan; Aquatic Ecology Technical Report 2005/19; Illinois Natural History Survey Center for Aquatic Ecology: Zion, IL, USA, 2005; Available online: https://www.ideals.illinois.edu/bitstream/handle/2142/10091/inhscaev02005i00019_opt.pdf?sequence=2&isAllowed=y (accessed on 1 June 2020).
- Sparks, R.E.; Barkley, T.L.; Creque, S.M.; Dettmers, J.M.; Stainbrook, K.M. Evaluation of an electric fish dispersal barrier in the Chicago Sanitary and Ship Canal. In Invasive Asian Carps in North America, American Fisheries Society Symposium; Chapman, D.C., Hoff, M.H., Eds.; American Fisheries Society: Bethesda, MD, USA, 2010; Volume 74, pp. 139–161. [Google Scholar]
- Evans, N.T.; Brouder, M.J. Asian Carp Entrainment, Retainment and Upstream Transport by Commercial Barge tows on the Illinois Waterway—2018 Trials; US Fish & Wildlife Service Report; US Fish and Wildlife Service Carterville Fish and Wildlife Conservation Office: Willmington, IL, USA, 2020. Available online: https://www.fws.gov/midwest/fisheries/carterville/documents/2018-Barge-Entrainment-Study-Report.pdf (accessed on 1 June 2020).
- Parker, A.D.; Rogers, P.B.; Finney, S.T.; Simmonds, R.L.J. Preliminary Results of Fixed DIDSON Evaluations at the Electric Dispersal Barrier in the Chicago Sanitary and Ship Canal; US Fish & Wildlife Service Report; US Fish and Wildlife Service, Carterville Fish and Wildlife Conservation Office: Willmington, IL, USA, 2013. Available online: https://www.fws.gov/midwest/fisheries/carterville/documents/DIDSON.pdf (accessed on 1 June 2020).
- Reynolds, J.B. Electrofishing. In Fisheries Techniques, 2nd ed.; Murphy, B.R., Willis, D.W., Eds.; American Fisheries Society: Bethesda, MD, USA, 1996; pp. 221–253. ISBN 9781888569001. [Google Scholar]
- Noatch, M.R.; Suski, C.D. Non-Physical Barriers to Deter Fish Movements. Environ. Rev. 2012, 20, 71–82. [Google Scholar] [CrossRef]
- USACE. The Great Lakes and Mississippi River Interbasin Study—Brandon Road Final Integrated Feasibility Study and Environmental Impact Statement—Will County, Illinois; US Army Corps of Engineers, Rock Island and Chicago Districts: Rock Island, TN, USA; Chicago, IL, USA, 2018; Available online: https://usace.contentdm.oclc.org/utils/getfile/collection/p16021coll7/id/11394 (accessed on 1 June 2020).
- Cooke, S.; Hill, W.R. Can Filter-Feeding Asian Carp Invade the Laurentian Great Lakes? A Bioenergetic Modelling Exercise. Freshw. Boil. 2010, 55, 2138–2152. [Google Scholar] [CrossRef]
- Cuddington, K.; Currie, W.J.S.; Koops, M.A. Could an Asian Carp Population Establish in the Great Lakes from a Small Introduction? Boil. Invasions 2013, 16, 903–917. [Google Scholar] [CrossRef]
- Wittmann, M.E.; Cooke, R.M.; Rothlisberger, J.D.; Rutherford, E.S.; Zhang, H.; Mason, D.M.; Lodge, D.M. Use of Structured Expert Judgment to Forecast Invasions by Bighead and Silver Carp in Lake Erie. Conserv. Boil. 2014, 29, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauber, T.B.; Stedman, R.C.; Connelly, N.A.; Rudstam, L.G.; Ready, R.C.; Poe, G.L.; Bunnell, D.B.; Höök, T.O.; Koops, M.A.; Ludsin, S.A.; et al. Using Scenarios to Assess Possible Future Impacts of Invasive Species in the Laurentian Great Lakes. N. Am. J. Fish. Manag. 2016, 36, 1292–1307. [Google Scholar] [CrossRef]
- Zhang, H.; Rutherford, E.S.; Mason, D.M.; Breck, J.T.; Wittmann, M.E.; Cooke, R.M.; Lodge, D.M.; Rothlisberger, J.D.; Zhu, X.; Johnson, T.B. Forecasting the Impacts of Silver and Bighead Carp on the Lake Erie Food Web. Trans. Am. Fish. Soc. 2015, 145, 136–162. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.P.; Strowitzki, M.J.; Taylor, C.T. Mechanisms and Consequences of Oxygen and Carbon Dioxide Sensing in Mammals. Physiol. Rev. 2020, 100, 463–488. [Google Scholar] [CrossRef]
- Cummins, E.P.; Selfridge, A.C.; Sporn, P.H.S.; Sznajder, J.I.; Taylor, C.T. Carbon Dioxide-Sensing in Organisms and Its Implications for Human Disease. Cell. Mol. Life Sci. 2013, 71, 831–845. [Google Scholar] [CrossRef] [Green Version]
- Thom, C.; Guerenstein, P.G.; Mechaber, W.L.; Hildebrand, J.G. Floral CO2 Reveals Flower Profitability to Moths. J. Chem. Ecol. 2004, 30, 1285–1288. [Google Scholar] [CrossRef]
- Seeley, T.D. Atmospheric Carbon Dioxide Regulation in Honey-Bee (Apis mellifera) Colonies. J. Insect Physiol. 1974, 20, 2301–2305. [Google Scholar] [CrossRef]
- Gillies, M.T. The Role of Carbon Dioxide in Host-Finding by Mosquitoes (Diptera: Culicidae): A Review. Bull. Entomol. Res. 1980, 70, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Takken, W.; Knols, B.G.J. Odor-Mediated Behavior of Afrotropical Malaria Mosquitoes. Annu. Rev. Entomol. 1999, 44, 131–157. [Google Scholar] [CrossRef] [PubMed]
- Faucher, C. Behavioral Responses of Drosophila to Biogenic Levels of Carbon Dioxide Depend on Life-Stage, Sex and Olfactory Context. J. Exp. Boil. 2006, 209, 2739–2748. [Google Scholar] [CrossRef] [Green Version]
- Lahiri, S.; Forster II, R.E. CO2/H+ Sensing: Peripheral and Central Chemoreception. Int. J. Biochem. Cell Biol. 2003, 35, 1413–1435. [Google Scholar] [CrossRef]
- Shusterman, D. Individual Factors in Nasal Chemesthesis. Chem. Senses 2002, 27, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Tresguerres, M.; Milsom, W.K.; Perry, S.F. CO2 and acid-base sensing. In Carbon Dioxide; Farrell, A.P., Brauner, C.J., Eds.; Elsevier: San Diego, CA, USA, 2019; Volume 37, pp. 33–68. ISBN 9780128176108. [Google Scholar]
- Eddy, F.B.; Lomholt, J.P.; Weber, R.E.; Johansen, K. Blood Respiratory Properties of Rainbow Trout (Salmo gairdneri) Kept in Water of High CO2 Tension. J. Exp. Boil. 1977, 67, 37–47. [Google Scholar]
- Brauner, C.J.; Baker, D.W. Patterns of acid–base regulation during exposure to hypercarbia in fishes. In Cardio-Respiratory Control in Vertebrates; Glass, M.L., Wood, S.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 43–63. ISBN 978-3-540-93984-9. [Google Scholar]
- Bernier, N.J.; Randall, D.J. Carbon Dioxide Anaesthesia in Rainbow Trout: Effects of Hypercapnic Level and Stress on Induction and Recovery from Anaesthetic Treatment. J. Fish Biol. 1998, 52, 621–637. [Google Scholar]
- Dennis, C.E.; Kates, D.F.; Noatch, M.R.; Suski, C.D. Molecular Responses of Fishes to Elevated Carbon Dioxide. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 187, 224–231. [Google Scholar] [CrossRef]
- Kates, D.; Dennis, C.; Noatch, M.R.; Suski, C.D. Responses of Native and Invasive Fishes to Carbon Dioxide: Potential for a Nonphysical Barrier to Fish Dispersal. Can. J. Fish. Aquat. Sci. 2012, 69, 1748–1759. [Google Scholar] [CrossRef]
- Iwama, G.K.; McGeer, J.C.; Pawluk, M.P. The Effects of Five Fish Anaesthetics on Acid-Base Balance, Hematocrit, Blood Gases, Cortisol, and Adrenaline in Rainbow Trout. Can. J. Zool. 1989, 67, 2065–2073. [Google Scholar] [CrossRef]
- Brauner, C.J.; Seidelin, M.; Madsen, S.S.; Jensen, F.B. Effects of Freshwater Hyperoxia and Hypercapnia and Their Influences on Subsequent Seawater Transfer in Atlantic Salmon (Salmo salar) Smolts. Can. J. Fish. Aquat. Sci. 2000, 57, 2054–2064. [Google Scholar] [CrossRef]
- Fish, F.F. The Anaesthesia of Fish by High Carbon-Dioxide Concentrations. Trans. Am. Fish. Soc. 1943, 72, 25–29. [Google Scholar] [CrossRef]
- Post, G. Carbonic Acid Anesthesia for Aquatic Organisms. Progress. Fish-Culturist 1979, 41, 142–144. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Yokoyama, Y.; Ueno, S.; Mitsuda, H. Changes of Blood Gas in Carp, Cyprinus Carpio, Anesthetized with Carbon Dioxide. Comp. Biochem. Physiol. Part A Physiol. 1991, 98, 431–436. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Kawai, F.; Kanamori, M. The Relationship between the EEG and Brain pH in Carp, Cyprinus carpio, Subjected to Environmental Hypercapnia at an Anesthetic Level. Comp. Biochem. Physiol. A Physiol. 1994, 107, 307–312. [Google Scholar]
- Beitinger, T.L. Behavioral Reactions for the Assessment of Stress in Fishes. J. Great Lakes Res. 1990, 16, 495–528. [Google Scholar] [CrossRef]
- Tierney, K.B. Chemical Avoidance Responses of Fishes. Aquat. Toxicol. 2016, 174, 228–241. [Google Scholar] [CrossRef]
- Shelford, V.E.; Allee, W.C. The Reactions of Fishes to Gradients of Dissolved Atmospheric Gases. J. Exp. Zool. 1913, 14, 207–266. [Google Scholar] [CrossRef]
- Powers, E.B.; Clark, R.T. Further Evidence on Chemical Factors Affecting the Migratory Movements of Fishes, Especially the Salmon. Ecology 1943, 24, 109–113. [Google Scholar] [CrossRef]
- Collins, B.G. Factors Influencing the Orientation of Migrating Anadromous Fishes. Fish. Bull. 1952, 52, 375–396. [Google Scholar]
- Bishai, H.M. Reactions of Larval and Young Salmonids to Different Hydrogen Ion Concentrations. ICES J. Mar. Sci. 1962, 27, 181–191. [Google Scholar] [CrossRef]
- Jones, K.A.; Hara, T.J.; Scherer, E. Locomotor Response by Arctic Char (Salvelinus alpinus) to Gradients of H+ and CO2. Physiol. Zool. 1985, 58, 413–420. [Google Scholar] [CrossRef]
- Ross, R.M.; Krise, W.F.; Redell, L.A.; Bennett, R.M. Effects of Dissolved Carbon Dioxide on the Physiology and Behavior of Fish in Artificial Streams. Environ. Toxicol. 2001, 16, 84–95. [Google Scholar] [CrossRef]
- Clingerman, J.; Bebak, J.; Mazik, P.M.; Summerfelt, S.T. Use of Avoidance Response by Rainbow Trout to Carbon Dioxide for Fish Self-Transfer between Tanks. Aquac. Eng. 2007, 37, 234–251. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, H.; Ishida, Y.; Ueno, S.; Mitsuda, H. Anesthetic Effect of CO2 on Fish. I. Changes in Depth of Anesthesia of the Carp Anesthetized with a Constant Level of CO2. Nippon. Suisan Gakkaishi 1988, 54, 457–462. [Google Scholar] [CrossRef]
- Pirhonen, J.; Schreck, C.B. Effects of Anaesthesia with MS-222, Clove Oil and CO2 on Feed Intake and Plasma Cortisol in Steelhead Trout (Oncorhynchus mykiss). Aquaculture 2003, 220, 507–514. [Google Scholar] [CrossRef]
- Dennis, C.E.; Adhikari, S.; Suski, C.D. Molecular and Behavioral Responses of Early-Life Stage Fishes to Elevated Carbon Dioxide. Boil. Invasions 2015, 17, 3133–3151. [Google Scholar] [CrossRef]
- Summerfelt, R.C.; Lewis, W.M. Repulsion of Green Sunfish by Certain Chemicals. J. Water Pollut. Control. Fed. 1967, 39, 2030–2038. [Google Scholar]
- Donaldson, M.R.; Amberg, J.; Adhikari, S.; Cupp, A.R.; Jensen, N.; Romine, J.; Wright, A.; Gaikowski, M.P.; Suski, C.D. Carbon Dioxide as a Tool to Deter the Movement of Invasive Bigheaded Carps. Trans. Am. Fish. Soc. 2016, 145, 657–670. [Google Scholar] [CrossRef]
- Cupp, A.R.; Erickson, R.; Fredricks, K.T.; Swyers, N.M.; Hatton, T.W.; Amberg, J.J. Responses of Invasive Silver and Bighead Carp to a Carbon Dioxide Barrier in Outdoor Ponds. Can. J. Fish. Aquat. Sci. 2017, 74, 297–305. [Google Scholar] [CrossRef]
- Cupp, A.R.; Smerud, J.; Tix, J.; Schleis, S.; Fredricks, K.; Erickson, R.A.; Amberg, J.; Morrow, W.; Koebel, C.; Murphy, E.; et al. Field Evaluation of Carbon Dioxide as a Fish Deterrent at a Water Management Structure along the Illinois River. Manag. Boil. Invasions 2018, 9, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Hasler, C.T.; Woodley, C.M.; Schneider, E.V.; Hixson, B.K.; Fowler, C.J.; Midway, S.R.; Suski, C.D.; Smith, D.L. Avoidance of Carbon Dioxide in Flowing Water by Bighead Carp. Can. J. Fish. Aquat. Sci. 2019, 76, 961–969. [Google Scholar] [CrossRef]
- Sampson, S.J.; Chick, J.H.; Pegg, M.A. Diet Overlap among Two Asian Carp and Three Native Fishes in Backwater Lakes on the Illinois and Mississippi Rivers. Boil. Invasions 2008, 11, 483–496. [Google Scholar] [CrossRef]
- Deters, J.E.; Chapman, D.C.; McElroy, B. Location and Timing of Asian Carp spawning in the Lower Missouri River. Environ. Boil. Fishes 2012, 96, 617–629. [Google Scholar] [CrossRef]
- Réale, D.; Garant, D.; Humphries, M.M.; Bergeron, P.; Careau, V.; Montiglio, P.-O. Personality and the Emergence of the Pace-Of-Life Syndrome Concept at the Population Level. Philos. Trans. R. Soc. B Boil. Sci. 2010, 365, 4051–4063. [Google Scholar] [CrossRef] [Green Version]
- Myles-Gonzalez, E.; Burness, G.; Yavno, S.; Rooke, A.C.; Fox, M.G. To Boldly Go Where No Goby Has Gone Before: Boldness, Dispersal Tendency, and Metabolism at the Invasion Front. Behav. Ecol. 2015, 26, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Cockrem, J.F. Stress, Corticosterone Responses and Avian Personalities. J. Ornithol. 2007, 148, 169–178. [Google Scholar] [CrossRef]
- Koolhaas, J.; Korte, S.M.; De Boer, S.; Van Der Vegt, B.; Van Reenen, C.; Hopster, H.; De Jong, I.; Ruis, M.; Blokhuis, H. Coping Styles in Animals: Current Status in Behavior and Stress-Physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating Animal Temperament within Ecology and Evolution. Boil. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Tucker, E.K.; Suski, C.D.; Philipp, M.A.; Jeffrey, J.D.; Hasler, C.T. Glucocorticoid and Behavioral Variation in Relation to Carbon Dioxide Avoidance across Two Experiments in Freshwater Teleost Fishes. Boil. Invasions 2018, 21, 505–517. [Google Scholar] [CrossRef]
- Tucker, E.K.; Suski, C.D. Presence of Conspecifics Reduces Between-Individual Variation and Increases Avoidance of Multiple Stressors in Bluegill. Anim. Behav. 2019, 158, 15–24. [Google Scholar] [CrossRef]
- Killen, S.S.; Marras, S.; Metcalfe, N.B.; McKenzie, D.J.; Domenici, P. Environmental Stressors Alter Relationships between Physiology and Behaviour. Trends Ecol. Evol. 2013, 28, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, N.B.; Van Leeuwen, T.E.; Killen, S.S. Does Individual Variation in Metabolic Phenotype Predict Fish Behaviour and Performance? J. Fish Boil. 2015, 88, 298–321. [Google Scholar] [CrossRef] [PubMed]
- Suski, C.D.; Philipp, M.A.; Hasler, C.T. Influence of Nutritional Status on Carbon Dioxide Tolerance and Avoidance Behavior in a Freshwater Teleost. Trans. Am. Fish. Soc. 2019, 148, 914–925. [Google Scholar] [CrossRef]
- Nadler, L.E.; Killen, S.S.; McClure, E.C.; Munday, P.L.; McCormick, M.I. Shoaling Reduces Metabolic Rate in a Gregarious Coral Reef Fish Species. J. Exp. Boil. 2016, 219, 2802–2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, C.E.; Adhikari, S.; Wright, A.W.; Suski, C.D.; Dennis, C.E. Molecular, Behavioral, and Performance Responses of Juvenile Largemouth Bass Acclimated to an Elevated Carbon Dioxide Environment. J. Comp. Physiol. B 2016, 186, 297–311. [Google Scholar] [CrossRef]
- Cupp, A.R.; Tix, J.; Smerud, J.; Erickson, R.A.; Fredricks, K.; Amberg, J.; Suski, C.D.; Wakeman, R. Using Dissolved Carbon Dioxide to Alter the Behavior of Invasive round Goby. Manag. Boil. Invasions 2017, 8, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Tix, J.A.; Cupp, A.R.; Smerud, J.R.; Erickson, R.; Fredricks, K.T.; Amberg, J.J.; Suski, C.D. Temperature Dependent Effects of Carbon Dioxide on Avoidance Behaviors in Bigheaded Carps. Boil. Invasions 2018, 20, 3095–3105. [Google Scholar] [CrossRef]
- Beitinger, T.L.; Lutterschmidt, W.I. Measures of thermal tolerance. In Encyclopedia of Fish Physiology, 1st ed.; Farrell, A.P., Ed.; Elsevier: Waltham, MA, USA, 2011; pp. 1695–1702. ISBN 9780080923239. [Google Scholar]
- Somero, G. Temporal Patterning of Thermal Acclimation: From Behavior to Membrane Biophysics. J. Exp. Boil. 2015, 218, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Hasler, C.T.; Bouyoucos, I.A.; Suski, C.D. Tolerance to Hypercarbia Is Repeatable and Related to a Component of the Metabolic Phenotype in a Freshwater Fish. Physiol. Biochem. Zool. 2017, 90, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Killen, S.S.; Adriaenssens, B.; Marras, S.; Claireaux, G.; Cooke, S.J. Context Dependency of Trait Repeatability and Its Relevance for Management and Conservation of Fish Populations. Conserv. Physiol. 2016, 4, cow007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelwicks, K.R.; Zafft, D.J.; Bobbitt, J.P. Efficacy of Carbonic Acid as an Anesthetic for Rainbow Trout. N. Am. J. Fish. Manag. 1998, 18, 432–438. [Google Scholar] [CrossRef]
- Fivelstad, S.; Waagbø, R.; Stefansson, S.; Olsen, A.B. Impacts of Elevated Water Carbon Dioxide Partial Pressure at Two Temperatures on Atlantic Salmon (Salmo salar L.) Parr Growth and Haematology. Aquaculture 2007, 269, 241–249. [Google Scholar] [CrossRef]
- Neiffer, D.L.; Stamper, M.A. Fish Sedation, Anesthesia, Analgesia, and Euthanasia: Considerations, Methods, and Types of Drugs. ILAR J. 2009, 50, 343–360. [Google Scholar] [CrossRef] [Green Version]
- Fredricks, K.T.; Hubert, T.D.; Amberg, J.J.; Cupp, A.R.; Dawson, V.K. Chemical Controls for an Integrated Pest Management Program. N. Am. J. Fish. Manag. 2019. Available online: https://0-afspubs-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/abs/10.1002/nafm.10339 (accessed on 2 June 2020). [CrossRef]
- Schneider, E.V.; Hasler, C.T.; Suski, C.D. Swimming Performance of a Freshwater Fish during Exposure to High Carbon Dioxide. Environ. Sci. Pollut. Res. 2018, 26, 3447–3454. [Google Scholar] [CrossRef]
- Ruebush, B.; Sass, G.; Chick, J.; Stafford, J. In-Situ Tests of Sound-Bubble-Strobe Light Barrier Technologies to Prevent Range Expansions of Asian Carp. Aquat. Invasions 2012, 7, 37–48. [Google Scholar] [CrossRef]
- Hasler, C.T.; Midway, S.R.; Jeffrey, J.D.; Tix, J.A.; Sullivan, C.; Suski, C.D. Exposure to Elevated pCO2 Alters Post-Treatment Diel Movement Patterns of Largemouth Bass over Short Time Scales. Freshw. Boil. 2016, 61, 1590–1600. [Google Scholar] [CrossRef]
- Hasler, C.T.; Jeffrey, J.D.; Butman, D.; Suski, C. Freshwater Biota and Rising pCO2? Ecol. Lett. 2016, 19, 98–108. [Google Scholar] [CrossRef]
- Jeffrey, J.D.; Hannan, K.D.; Hasler, C.T.; Suski, C.D. Hot and Bothered: Effects of Elevated pCO2 and Temperature on Juvenile Freshwater Mussels. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R115–R127. [Google Scholar] [CrossRef] [PubMed]
- Hannan, K.D.; Jeffrey, J.D.; Hasler, C.T.; Suski, C.D. Physiological Responses of Three Species of Unionid Mussels to Intermittent Exposure to Elevated Carbon Dioxide. Conserv. Physiol. 2016, 4, cow066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasler, C.T.; Jeffrey, J.D.; Schneider, E.V.; Hannan, K.D.; Tix, J.A.; Suski, C.D. Biological Consequences of Weak Acidification Caused by Elevated Carbon Dioxide in Freshwater Ecosystems. Hydrobiologia 2017, 806, 1–12. [Google Scholar] [CrossRef]
- Midway, S.R.; Hasler, C.T.; Wagner, T.; Suski, C.D. Predation of Freshwater Fish in Elevated Carbon Dioxide Environments. Mar. Freshwater Res. 2017, 68, 1585–1592. [Google Scholar] [CrossRef]
- Tix, J.A.; Hasler, C.T.; Sullivan, C.; Jeffrey, J.D.; Suski, C.D. Elevated Carbon Dioxide Has the Potential to Impact Alarm Cue Responses in Some Freshwater Fishes. Aquat. Ecol. 2016, 51, 59–72. [Google Scholar] [CrossRef]
- Robertson, M.; Hernandez, M.F.; Midway, S.R.; Hasler, C.T.; Suski, C.D. Shelter-Seeking Behavior of Crayfish, Procambarus Clarkii, in Elevated Carbon Dioxide. Aquat. Ecol. 2018, 52, 225–233. [Google Scholar] [CrossRef]
Figure 1.
Flow chart showing the possible reactions of fishes to a potentially adverse environmental stimulus, such as an area of elevated carbon dioxide [73].
Figure 1.
Flow chart showing the possible reactions of fishes to a potentially adverse environmental stimulus, such as an area of elevated carbon dioxide [73].

Figure 2.
Relative expression of c-fos mRNA extracted from the gill tissue of juvenile bighead carp (a), silver carp (b), bluegill (c), and largemouth bass (d) exposed to a two hypercarbic treatments. Relative mRNA expression of juvenile fish that had an exposure duration of 30 min are shown in black bars, while white bars show the mRNA expression of juvenile fish exposed for 60 min. Horizontal lines denote a significant CO2 concentration effect across exposure durations within a species. Dissimilar letters indicate significant differences between bars within a species. Data are mean ± SE, calculated relative to the expression of the reference gene (i.e., either 18s or ef1-a). For clarity, data are expressed relative to the mean of juvenile fish exposed to ambient water conditions [84].
Figure 2.
Relative expression of c-fos mRNA extracted from the gill tissue of juvenile bighead carp (a), silver carp (b), bluegill (c), and largemouth bass (d) exposed to a two hypercarbic treatments. Relative mRNA expression of juvenile fish that had an exposure duration of 30 min are shown in black bars, while white bars show the mRNA expression of juvenile fish exposed for 60 min. Horizontal lines denote a significant CO2 concentration effect across exposure durations within a species. Dissimilar letters indicate significant differences between bars within a species. Data are mean ± SE, calculated relative to the expression of the reference gene (i.e., either 18s or ef1-a). For clarity, data are expressed relative to the mean of juvenile fish exposed to ambient water conditions [84].

Figure 3.
Concentration of CO2 at which largemouth bass, bluegill, and silver carp displayed either an agitated activity (surface ventilations, twitching, or elevated swimming activity) (a) or movement out of high CO2 environment to a lower CO2 environment (b) during the course of avoidance trials [66].
Figure 3.
Concentration of CO2 at which largemouth bass, bluegill, and silver carp displayed either an agitated activity (surface ventilations, twitching, or elevated swimming activity) (a) or movement out of high CO2 environment to a lower CO2 environment (b) during the course of avoidance trials [66].

Figure 4.
Concentration of CO2 at which juvenile bluegill, largemouth bass, silver carp and bighead carp displayed avoidance behaviors [84].
Figure 4.
Concentration of CO2 at which juvenile bluegill, largemouth bass, silver carp and bighead carp displayed avoidance behaviors [84].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of studies quantifying CO2 thresholds that caused avoidance within the framework of generating a non-physical barrier for silver carp and bighead carp. Data have been approximated from figures where it was not clearly outlined in text of the citation. Units are left in the format that was used during publication.
Table 1.
Summary of studies quantifying CO2 thresholds that caused avoidance within the framework of generating a non-physical barrier for silver carp and bighead carp. Data have been approximated from figures where it was not clearly outlined in text of the citation. Units are left in the format that was used during publication.
| Species | Avoidance Threshold | Test Environment | Temperature | pH | Mean Fish Size (mm) | Citation |
|---|---|---|---|---|---|---|
| Silver carp | 135 mg/L | Shuttle box | 18 °C | 8.0 | 460 | [66] |
| 125 mg/L | Shuttle box | 16 °C | 7.46 | 67 | [65] | |
| 59 mg/L (29,193 μatm) | Outdoor static pond | 16 °C | 8.25 | 254 | [86] | |
| 75 mg/L (29,532–41,393 μatm) | Outdoor flowing raceway | 8–13 °C | 7.5 | 278 | [87] | |
| Bighead carp | 180 mg/L | Shuttle box | 16 °C | 7.46 | 71 | [65] |
| 59 mg/L (29,193 μatm) | Outdoor static pond | 16 °C | 8.25 | 205 | [86] | |
| 75 mg/L (29,532–41,393 μatm) | Outdoor flowing raceway | 8–13 °C | 7.5 | 212 | [87] | |
| 160,000–186,000 μatm | Indoor flowing raceway | 21 °C | 8.3 | 145 | [89] |
Table 2.
Factors influencing the thresholds of CO2 required to induce avoidance behaviors in fishes.
Table 2.
Factors influencing the thresholds of CO2 required to induce avoidance behaviors in fishes.
| Factor | Outcome | Species | Citation | |
|---|---|---|---|---|
| Factors resulting in more CO2 needed for avoidance | Temperature | Higher concentrations of CO2 needed to induce avoidance at high temperatures relative to low temperatures. | Bighead carp, silver carp, round goby | [104,105] |
| Stress | Fish with artificially-elevated cortisol levels required more CO2 to induce shuttling than non-stressed controls. | Largemouth bass | [97] | |
| Factors requiring less CO2 for avoidance | Shoals | Shoals of fish avoided CO2 at lower thresholds than did individual fish. | Bluegill | [97] |
| Factors not influencing CO2 avoidance | Social personality | Preference for associating with conspecifics did not influence thresholds for CO2 avoidance. | Bluegill | [97] |
| Personality | Activity and boldness did not influence CO2 avoidance thresholds. | Bluegill | [97] | |
| Feeding | Fish that had been deprived food for 9 days avoided CO2 at the same threshold as fed conspecifics | Largemouth bass | [101] |
Table 3.
Factors influencing the tolerance of CO2, indicated by loss of equilibrium.
| Factor | Outcome | Species | Citation | |
|---|---|---|---|---|
| Factors resulting in a higher threshold for equilibrium loss in CO2 | Size | large fish lost equilibrium sooner (were more sensitive) at a given CO2 concentration than small fish. | Rainbow trout | [81] |
| Time × concentration interaction | Equilibrium loss occurs at extended exposure to low CO2 concentration, or brief exposure to high CO2 concentration. | Several species of salmonid (steelhead, chinook) | [69,70] | |
| Temperature | When CO2 was added to a tank at a constant rate, a higher CO2 concentration was required to induce equilibrium loss at high temperatures relative to low temperatures. | Silver carp, bighead carp, round goby. | [104,105] | |
| Food deprivation | Fish that had been deprived food for 14 days required more CO2 to induce equilibrium loss than fed conspecifics. | Largemouth bass | [101] | |
| Factors resulting in a lower threshold for equilibrium loss in CO2 | Temperature | Equilibrium loss occurs faster at higher temperature when CO2 concentration is held constant. | Several species of salmonid (steelhead, chinook) | [69,110] |
| Anaerobic swimming potential | Fish that required longer to become exhausted during burst swimming required less time to lose equilibrium at high CO2. | Largemouth bass | [108] | |
| Factors not influencing equilibrium loss in high CO2 environments | Standard metabolic rate | Variation in standard metabolic rate did not predict time to equilibrium loss in high CO2. | Largemouth bass | [108] |
| Aerobic scope | Variation in aerobic scope did not predict time to equilibrium loss in high CO2. | Largemouth bass | [108] | |
| Personality (activity, boldness) | Variation in activity and boldness did not influence time to equilibrium loss in high CO2. | Bluegill | [97] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Suski, C.D. Development of Carbon Dioxide Barriers to Deter Invasive Fishes: Insights and Lessons Learned from Bigheaded Carp. Fishes 2020, 5, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030025
AMA Style
Suski CD. Development of Carbon Dioxide Barriers to Deter Invasive Fishes: Insights and Lessons Learned from Bigheaded Carp. Fishes. 2020; 5(3):25. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030025
Chicago/Turabian StyleSuski, Cory D. 2020. "Development of Carbon Dioxide Barriers to Deter Invasive Fishes: Insights and Lessons Learned from Bigheaded Carp" Fishes 5, no. 3: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5030025