
In Vitro Antimicrobial and Antibiofilm Activity of S-(-)-Limonene and R-(+)-Limonene against Fish Bacteria

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phytochemicals
2.2. Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Assays
2.3. Checkerboard Assay
2.4. Effect on Biofilm Formation
2.5. Effect on Biofilm Formation in Nylon Cubes
2.6. Atomic Force Microscopy
2.7. Statistical Analysis
3. Results
3.1. Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Assays
3.2. Checkerboard Assay
3.3. Effect of Biofilm Formation by Optical Microscopy
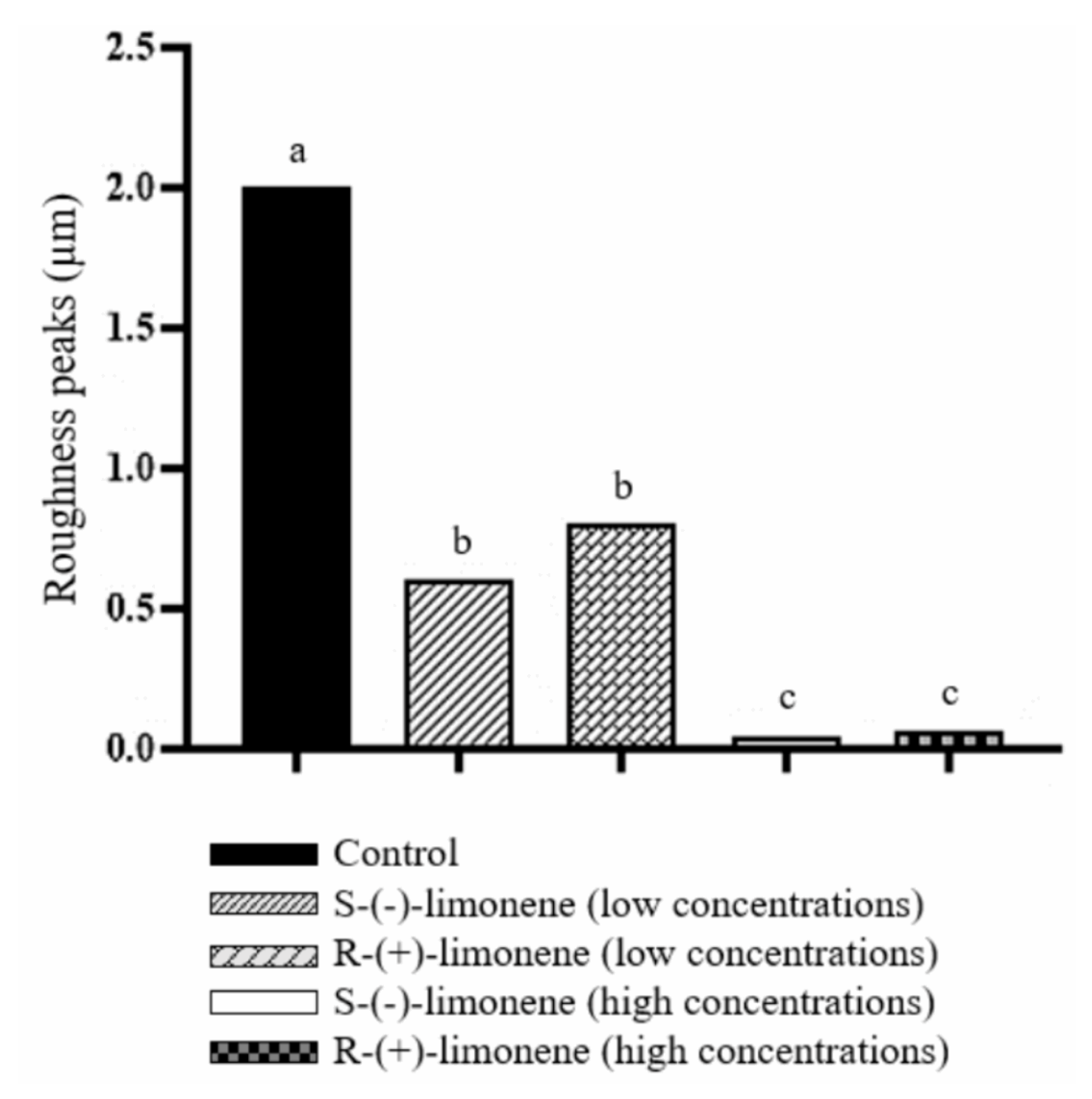
3.4. Effect of Biofilm Formation by Atomic Force Microscopy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterman, M.A.; Posadas, B.C. Direct Economic Impact of Fish Diseases on the East Mississippi Catfish Industry. N. Am. J. Aquac. 2018, 81, 222–229. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária. Pesca e Aquicultura; Embrapa: Palmas, Brazil, 2017. [Google Scholar]
- Pessoa, R.B.G.; Oliveira, W.F.; Marques, D.S.C.; Correia, M.T.S.; Carvalho, E.V.M.M.; Luana Coelho, C.B.B. The genus Aeromonas: A general approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcellos, L.J.G.; Kreutz, L.C.; Rodrigues, L.B.; Santos, L.R.; Motta, A.C.; Ritter, F.; Bedin, A.C.; Silva, L.B. Aeromonas hydrophila in Rhamdia quelen: Macroscopic and microscopic aspect of the lesions and antibiotic resistence profiles. Bol. Inst. Pesca. 2008, 34, 355–363. [Google Scholar]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Diseases of Farmed and Wild Fish, 2nd ed.; Springer-Praxis Publishing, Ltd.: Chichester, UK, 2007. [Google Scholar]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens, Disease of Farmed and Wild Fish, 6th ed.; Springer International Publishing: Basel, Switzerland, 2016. [Google Scholar]
- Ghatak, S.; Blom, J.; Das, S.; Sanjukta, R.; Puro, K.; Mawlong, M.; Shakuntala, I.; Sen, A.; Goesmann, A.; Kumar, A.; et al. Pan-genome analysis of Aeromonas hydrophila, Aeromonas veronii and Aeromonas caviae indicates phylogenomic diversity and greater pathogenic potential for Aeromonas hydrophila. Antonie Van Leeuwenhoek 2016, 109, 945–956. [Google Scholar] [CrossRef]
- Song, Y.; Hu, X.; Lü, A.; Sun, J.; Yiksung, Y.; Pei, C.; Zhang, C.; Li, L. Isolation and characterization of Aeromonas veronii from ornamental fish species in China. Isr. J. Aquac. 2017, 69, 1446. [Google Scholar] [CrossRef]
- Cai, S.H.; Wu, Z.H.; Jian, J.C.; Lu, Y.S.; Tang, J.F. Characterization of pathogenic Aeromonas veronii bv. veronii associated with ulcerative syndrome from Chinese longsnout catfish (Leiocassis longirostris Gunther). Braz. J. Microbiol. 2012, 43, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Seng, P.; Boushab, B.M.; Romain, F.; Gouriet, F.; Bruder, N.; Martin, C.; Paganelli, F.; Bernit, E.; Le Treut, Y.P.; Thomas, P.; et al. Emerging role of Raoultella ornithinolytica in human infections: A series of cases and review of the literature. Int. J. Infect. Dis. 2016, 45, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Lü, A.; Hu, X.; Zheng, L.; Zhu, A.; Cao, C.; Jiang, J. Isolation and characterization of Citrobacter spp. from the intestine of grass carp Ctenopharyngodon idellus. Aquaculture 2011, 313, 156–160. [Google Scholar] [CrossRef]
- Svetlana, J.; Dobrila, J.D.; Veljović, L.J. Citrobacter freundii as a cause of disease in fish. Acta Vet. 2003, 53, 399–410. [Google Scholar] [CrossRef]
- Hajjar, R.; Ambaraghassi, G.; Sebajang, H.; Schwenter, F.; Su, S.-H. Raoultella ornithinolytica: Emergence and Resistance. Infect. Drug Resist. 2020, 13, 1091–1104. [Google Scholar] [CrossRef] [Green Version]
- Gajdács, M.; Urbán, E. Prevalence and antibiotic resistance of Stenotrophomonas maltophilia in respiratory tract samples: A 10-Year Epidemiological Snapshot. Health Serv. Res. Manag. Epidemiol. 2019, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Wang, K.; Chen, D.; Huang, X.; He, M.; Yin, Z. Stenotrophomonas maltophilia, an emerging opportunist pathogen for cultured channel catfish, Ictalurus punctatus, in China. Aquaculture 2010, 308, 132–135. [Google Scholar] [CrossRef]
- Kapetanović, D. Gill disease in a gilthead sea bream (Sparus aurata L.). Med. Weter. 2006, 11, 62. [Google Scholar]
- Bandeira Junior, G.; Sutili, F.J.; Gressler, L.T.; Ely, V.L.; Silveira, B.P.; Tasca, C.; Reghelin, M.; Matter, L.B.; Vargas, A.P.C.; Baldisserotto, B. Antibacterial potential of phytochemicals alone or in combination with antimicrobials against fish pathogenic bactéria. J. Appl. Microbiol. 2018, 125, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Sebastião, F.D.A.; Furlan, L.; Hashimoto, D.T.; Pilarski, F. Identification of bacterial fish pathogens in Brazil by direct colony PCR and 16S rRNA gene sequencing. Adv. Microbiol. 2015, 5, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Sutili, F.J.; Kreutz, L.C.; Noro, M.; Gressler, L.T.; Heinzmann, B.M.; de Vargas, A.C.; Baldisserotto, B. The use of eugenol against Aeromonas hydrophila and its effect on hematological and immunological parameters in silver catfish (Rhamdia quelen). Vet. Immunol. Immunopathol. 2014, 157, 142–148. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Totsika, M.; Kathryn, E.; Fairfull-Smith, K.E. Bacterial Biofilm eradication agents: A Current Review. Front. Chem. 2019, 28, 824. [Google Scholar] [CrossRef] [Green Version]
- Grenni, P.; Ancona, V.; Caraccioloa, A.B. Ecological effects of antibiotics on natural ecossystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Rasul, M.G.; Majumdar, B.C. Abuse of antibiotics in aquaculture and it’s effects on human, aquatic animal and environment. Haya: Saudi J. Life Sci. 2017, 2, 81–88. [Google Scholar] [CrossRef]
- Cunha, J.A.; Heinzmann, B.M.; Baldisserotto, B. The effects of essential oils and their major compounds on fish bacterial pathogens—A review. J. Appl. Microbiol. 2018, 125, 328–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutili, F.J.; Silva, L.d.L.; Gressler, L.T.; Battisti, E.K.; Heinzmann, B.M.; de Vargas, A.C.; Baldisserotto, B. Plant essential oils against Aeromonas hydrophila: In vitro activity and their use in experimentally infected fish. J. Appl. Microbiol. 2015, 119, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Debbarma, J.; Kishore, P.; Nayak, B.B.; Kannuchamy, N.; Gudipati, N. Antibacterial activity of ginger, eucalyptus and sweet orange peel essential oils on fish-borne bacteria. J. Food Process. Technol. 2012, 37, 1022–1030. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A. Chemistry and bioactivity of essential oils. In Lipids and Essential Oils as Antimicrobial Agents; Thormar, H., Ed.; Wiley: Chichester, UK, 2011; pp. 203–238. [Google Scholar] [CrossRef]
- Vieira, A.J.; Beserra, F.P.; Souza, M.C.; Totti, B.M.; Rozza, A.L. Limonene: Aroma of innovation in health and disease. Chem. Biol. Interact. 2018, 283, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhard, T.J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Wissing, S.A.; Müller, R.H. Cosmetic applications for solid lipid nanoparticles (SLN). Int. J. Pharm. 2003, 254, 65–68. [Google Scholar] [CrossRef]
- Tongnuanchan, P.; Benjakul, S.; Prodpran, T. Properties and antioxidant activity of fish skin gelatin film incorporated with citrus essential oils. Food Chem. 2012, 134, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Pirjo, K.; Aflatuni, A.; Tiilikkala, K.; Holopainen, J.K. Insecticidal, repellent, antimicrobial activity and phytotoxicity of essential oils: With special reference to limonene and its suitability for control of insect pests. Agric. Food. Sci. Finl. 2001, 10, 243–259. [Google Scholar] [CrossRef]
- Song, X.; Liu, T.; Wang, L.; Liu, L.; Li, X.; Wu, X. Antibacterial effects and mechanism of mandarin (Citrus reticulata L.) essential oil against Staphylococcus aureus. Molecules 2020, 25, 4956. [Google Scholar] [CrossRef]
- Haraoui, N.; Allem, R.; Chaouche, T.M.; Belouazni, A. In-vitro antioxidant and antimicrobial activities of some varieties citrus grown in Algeria. Orient Pharm. Exp. Med. 2019, 20, 23–24. [Google Scholar] [CrossRef]
- CLSI. Methods for Broth Dilution Susceptibility Testing of Bacteria Isolated from Aquatic Animals, Approved Guideline, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Rand, K.H.; Houck, H.J.; Brown, P.; Bennett, D. Reproducibility of the microdilution checkerboard method for antibiotic synergy. Antimicrob. Agents Chemother. 1993, 37, 613–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, L.S.; Isenberg, H.D. Clinical Microbiology Procedures Handbook, 3rd ed.; ASM Press: Washington, DC, USA, 2010. [Google Scholar]
- Souza, A.M.; Armstrong, L.; Merino, F.J.Z.; Cogo, L.L.; Monteiro, C.L.B.; Duarte, M.R.; Miguel, O.G.; Miguel, M.D. In vitro effects of Eugenia pyriformis Cambess, Myrtaceae: Antimicrobial activity and synergistic interactions with vancomycin and fluconazole. Afr. J. Pharm. Pharmacol. 2014, 8, 862–867. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Stepanović, S.; Cirković, I.; Ranin, L.; Svabić-Vlahović, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef]
- Peixe, B.R. Anuário Peixe BR da Psicultura 2020; Associação Brasileira da Piscicultura: Pinheiros, Brasil, 2020. [Google Scholar]
- Miyake, D.; Kasahara, Y.; Morisaki, H. Distribution and characterization of antibiotic resistant bacteria in the sediment of southern basin of Lake Biwa. Microbes Environ. 2003, 18, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Lis-Balchin, M.; Ochocka, R.J.; Deans, S.; Asztemborska, M.; Hart, S. Bioactivity of the enantiomers of limonene. Med. Sci. Res. 1996, 24, 309–310. [Google Scholar]
- Lee, G.W.; Chung, M.S.; Kang, M.; Chung, Y.B.; Lee, S. Direct suppression of a rice bacterial blight (Xanthomonas oryzae pv. oryzae) by monoterpene (S)-limonene. Protoplasma 2016, 253, 683–690. [Google Scholar] [CrossRef]
- Omran, S.M.; Moodi, M.A.; Amiri, S.M.B.N.; Mosavi, S.J.; Saeed, S.A.M.G.M.; Shiade, S.M.J.; Kheradi, E.; Salehi, M. The effects of limonene and orange peel extracts on some spoilage fungi. Int. J. Mol. Clin. Microbiol. 2011, 1, 82–86. [Google Scholar]
- Pathirana, H.N.K.S.; Wimalasena, S.H.M.P.; De Silva, B.C.J.; Hossain, S.; Heo, G.-J. Antibacterial activity of lime (Citrus aurantifolia) essential oil and limonene against fish pathogenic bacteria isolated from cultured olive flounder (Paralichthys olivaceus). Fish. Aquat. Life 2018, 26, 131–139. [Google Scholar] [CrossRef]
- Ríos, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef] [PubMed]
- SINDAN. Compêndio de Produtos Veterinários; SINDAN: Vila Olímpia, Brazil, 2018. [Google Scholar]
- Botelho, R.G.; Christofoletti, C.A.; Correia, J.E.; Ansoar, Y.; Olinda, R.A.; Tornisielo, V.L. Genotoxic responses of juvenile tilapia (Oreochromis niloticus) exposed to florfenicol and oxytetracycline. Chemosphere 2015, 132, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Bowser, P.R. Pharmacokinetics of oxytetracycline in Nile tilapia Oreochromis niloticus challenged with Streptococcus iniae and Vibrio vulnificus. J. World Aquac. Soc. 2005, 36, 262–270. [Google Scholar] [CrossRef]
- Liu, X.; Steele, J.C.; Meng, X.Z. Usage, residue, and human health risk of antibiotics in Chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef]
- Yoshimizu, M.; Kasai, H.; Aoki, T.; Ototake, M.; Sakai, M.; Jung, T.-S.; Hikima, J.; Okamoto, N.; Sakamoto, T.; Ozaki, A.; et al. Fish Diseases—Prevention and Treatment of Diseases Caused by Fish Pathogens; Eolss Publishers/UNESCO: Oxford, UK, 2016. [Google Scholar]
- Giguère, S.; Prescott, J.F.; Dowling, P.M. Antimicrobial Therapy in Veterinary Medicine, 5th ed.; Wiley: Blackwell, UK, 2013. [Google Scholar]
- Assane, I.M.; Gozi, K.S.; Valladão, G.M.R.; Pilarski, F. Combination of antimicrobials as an approach to reduce their application in aquaculture: Emphasis on the use of thiamphenicol/florfenicol against Aeromonas hydrophila. Aquacullture 2019, 507, 238–245. [Google Scholar] [CrossRef]
- Milatovic, D.; Braveny, I. Development of resistance during antibiotic therapy. Eur. J. Clin. Microbiol. 1987, 6, 234–244. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ chemical characterization and investigation of some biological activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Misra, G.; Pavlostathis, S.G.; Perdue, E.M.; Araujo, R. Aerobic biodegradation of selected monoterpenes. Appl. Microbiol. Biotechnol. 1996, 45, 831–838. [Google Scholar] [CrossRef]
- Nguyen, L.A.; He, H.; Pham-Huy, C. Chiral Drugs: An Overview. Int. J. Biomed. Sci. 2006, 2, 85–100. [Google Scholar]
- Chotsaeng, N.; Laosinwattana, C.; Charoenying, P. Enantioselective and synergistic herbicidal activities of common amino acids against Amaranthus tricolor and Echinochloa crus-galli. Molecules 2021, 26, 2071. [Google Scholar] [CrossRef]
- Vuuren, S.F.; Viljoen, A.M. Antimicrobial activity of limonene enantiomers and 1,8-cineole alone and in combination. Flavour Fragr. J. 2007, 22, 540–544. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of Biofilm Resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Millezi, A.F.; Cardoso, M.G.; Alves, E.; Piccoli, R.H. Reduction of Aeromonas hidrophyla biofilm on stainless stell surface by essential oils. Braz. J. Microbiol. 2013, 44, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Aparna, M.S.D.; Yadav, S. Biofilms: Microbes and Disease. Braz. J. Infect. Dis. 2008, 12, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramenium, G.A.; Vijayakumar, K.; Pandian, S.K. Limonene inhibits streptococcal biofilm formation by targeting surface-associated virulence factors. J. Med. Microbiol. 2015, 64, 879–890. [Google Scholar] [CrossRef]
- Lopes, L.Q.S.; De Almeida Vaucher, R.; Giongo, J.L.; Gündel, A.; Santos, R.C.V. Characterisation and anti-biofilm activity of glycerol monolaurate nanocapsules against Pseudomonas aeruginosa. Microb. Pathog. 2019, 130, 178–185. [Google Scholar] [CrossRef]
- Jacques, M.; Aragon, V.; Tremblay, Y.D.N. Biofilm pathogens of veterinary importance. Anim. Health Res. Rev. 2010, 11, 97–121. [Google Scholar] [CrossRef]
- Anand, S.; Singh, D.; Avadhanula, M.; Marka, S. Development and control of bacterial biofilms on dairy processing membranes. Compr. Rev. Food Sci. Food Saf. 2014, 13, 18–33. [Google Scholar] [CrossRef]
- Cai, W.; Arias, C.R. Biofilm Formation on Aquaculture Substrates by Selected Bacterial Fish Pathogens. J. Aquat. Anim. Health 2017, 29, 95–104. [Google Scholar] [CrossRef]
- Lou, Z.; Chen, J.; Yu, F.; Wang, H.; Kou, X.; Ma, C.; Zhu, S. The antioxidant, antibacterial, antibiofilm activity of essential oil from Citrus medica L. var. sarcodactylis and its nanoemulsion. LWT-Food Sci. Technol. 2017, 80, 371–377. [Google Scholar] [CrossRef]
- Avcioglu, N.H.; Sahal, G.; Bilka, S.I. Antibiofilm effects of Citrus limonum and Zingiber officinale oils on biofilm formation of Klebsiella ornithinolytica, Klebsiella oxytoca and Klebsiella terrigena species. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 61–67. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Aeromonas hydrophila ATCC® 7966 | Aeromonas hydrophila MF 372509 | Aeromonas hydrophila MF 372510 | Aeromonas veronii MH 397688 | Citrobacter freundii MF 565839 | Raoultella ornithinolytica MF 372511 | Stenotrophomonas maltophilia MT 572493 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | MIC mg mL−1 | MBC mg mL−1 | |
| S-(−)-limonene | >6.4 | >6.4 | >6.4 | >6.4 | 3.2 | 3.2 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 |
| R-(+)-limonene | >6.4 | >6.4 | >6.4 | >6.4 | 6.4 | 6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 | >6.4 |
| DRUGS | FIC | FICI |
|---|---|---|
| SL—FLF | ||
| SL | 0.125 | 2.125 b |
| FLF | 2 | |
| SL—OXT | ||
| SL | 0.125 | 4.125 c |
| OXT | 4 | |
| RL—FLF | ||
| RL | 0.125 | 2.125 b |
| FLF | 2 | |
| RL—OXT | ||
| RL | 0.125 | 4.125 c |
| OXT | 4 | |
| SL—RL | ||
| SL | 4 | 8 c |
| RL | 4 | |
| FLF—OXT | ||
| FLF | 1 | 2 b |
| OXT | 1 |
| Aeromonas hydrophila MF 372510 | ||
|---|---|---|
| S-(-)-limonene | R-(+)-limonene | |
| MIC × 8 | 0 | 1 |
| MIC × 4 | 0 | 3 |
| MIC × 2 | 1 | 3 |
| MIC | 3 | 3 |
| MIC/2 | 3 | 3 |
| MIC/4 | 3 | 3 |
| MIC/8 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, E.G.; Bandeira Junior, G.; Cargnelutti, J.F.; Santos, R.C.V.; Gündel, A.; Baldisserotto, B. In Vitro Antimicrobial and Antibiofilm Activity of S-(-)-Limonene and R-(+)-Limonene against Fish Bacteria. Fishes 2021, 6, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6030032
da Silva EG, Bandeira Junior G, Cargnelutti JF, Santos RCV, Gündel A, Baldisserotto B. In Vitro Antimicrobial and Antibiofilm Activity of S-(-)-Limonene and R-(+)-Limonene against Fish Bacteria. Fishes. 2021; 6(3):32. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6030032
Chicago/Turabian Styleda Silva, Elisia Gomes, Guerino Bandeira Junior, Juliana Felipetto Cargnelutti, Roberto Christ Vianna Santos, André Gündel, and Bernardo Baldisserotto. 2021. "In Vitro Antimicrobial and Antibiofilm Activity of S-(-)-Limonene and R-(+)-Limonene against Fish Bacteria" Fishes 6, no. 3: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6030032