1. Introduction
The grey mullet,
Mugil cephalus, is an economically valuable euryhaline and eurythermal species in many countries, contributing to large fisheries in estuarine and coastal areas [
1,
2,
3]. Because of its strong tolerance to captivity, fast development, omnivorous feeding habits, and high market price, this fish species has been identified as a potential species for aquaculture diversification in the Mediterranean region, as well as in other parts of the world [
2,
4,
5]. This reality necessitates the advancement of breeding technologies, as well as a suitable and cost-effective grow-out diet. However, the majority of commercial grey mullet aquaculture production also relies on wild-caught fry, which is less expensive but not sustainable [
2,
6,
7]. Mullets are characterized as omnivorous, opportunistic feeders which thrive on any food that comes their way [
8]. However, knowledge is scarce about how to formulate realistic feed for cultured mullets [
9,
10].
Microcrustaceans, Copepods, zooplankton, detritus, and benthic microalgae are the main food sources for grey mullet larvae [
11]. This suggests that on-growing diets for this species could include high levels of fishmeal (FM) replacement with alternate protein sources [
12,
13,
14,
15]. This is also significant because the global supply of FM remains largely stable, resulting in high processing costs and a decline in its availability for large-scale use in high-quality fishmeal-based diets [
16,
17,
18]. More research is required to find more cost-effective alternative protein-rich feed ingredients that are appropriate for omnivorous fish species [
19,
20].
Alternative protein sources may be the main factor that helps in lowering feed costs and increasing the industry’s economic viability. Zooplankton has been deemed “nutritionally superior live feed” for fish larvae and juveniles of cultivable species, demonstrating that it has a higher nutritional benefit than Artemia; therefore, it plays an important role in initial feeding for survival and growth [
21,
22,
23]. Freshwater zooplankton biomass consists of three main species, Rotifera, Cladoceran, and Copepoda, which have been successfully used as live feed in the aquaculture industry [
24,
25,
26,
27]. According to biochemical research, zooplankton is high in proteins (45–52%), lipids (9–12%), essential amino acids, and essential fatty acids; it is a natural food source for many aquatic species and has high reproductive performance, as well as producing high biomass under the right culture conditions, enabling it to be used as an ingredient in the aqua-feed [
28,
29,
30].
Zooplankton meal succeeded in replacing 100% of FM in the diet of
Dicentrarchus labrax fingerlings without compromising growth performance and feed utilization [
31]. The study of Sharahi, et al. [
32] revealed that the replacement of FM with gammarus meal up to 20% significantly improved different growth performance criteria and feed utilization of Siberian sturgeon,
Acipenser baerii. In addition, the use of
Dahpnia magna meal as an FM alternative in the diet of grey mullet showed a quadratic regression trend in terms of growth and feed utilization, with the best replacement level at 75% of FM [
33]. The previous studies showed that the total zooplankton meal could replace 100% of FM; however, using individual types of zooplankton (daphnia or gammarus) partially succeeded in replacing FM in the aquatic diet. Accordingly, the use of total zooplankton meal as feed ingredient to replace FM in the diet of grey mullet could be a promising alternative. Therefore, this study was conducted to evaluate the growth performance, feed utilization, carcass traits, histological status, and economic evaluation of grey mullet fed diets with different FM replacement levels with zooplankton biomass meal (ZBM).
2. Materials and Methods
The fish rearing, handling, and sampling procedures were approved by the institutional animal care and use committee of Alexandria University with approval No. AU:19/21/05/27/3/18.
2.1. Fish and Feeding Management
Grey mullet larvae were obtained from the Mediterranean Sea, Rosita Fry Collection Center, (RFCC), Al-Behira, Egypt. Before the growth trial, all fish were adapted to laboratory conditions for two weeks. The fish were fed 50% Artemia nauplii (15–20 nauplii mL−1) and 50% control diet for 7 days, then gradually weaned to the control diet during the second 7 days. After that, 1500 apparent healthy juveniles (initial body weight (BW) 0.10 ± 0.01 g and standard length (SL) 1.36 ± 0.05 cm) were randomly distributed into 15 glass aquariums (100 L). All aquaria were assigned to five groups, each group with three replicates (n = 100 fry per aquarium), and the fish were fed five times daily (9:00, 12:00, 15:00, 18:00, and 21:00) for 60 days. The daily feeding rate was approximately 15% of the fish body weight. The husbandry system was supplied with sand-filtered, chlorinated, and UV-disinfected water and about 70% of the water was exchanged daily.
Water quality conditions during the acclimation and experimental periods were as follows: temperature of around 17.6–20.7 °C; dissolved oxygen, in a range from 5.40 to 6.45 mg L−1; NH4+, 0.16–0.29 mg L−1; the photoperiod was 10L:14D (light: darkness).
2.2. Preparation of Diets and Zooplankton Biomass Meal (ZBM)
Zooplankton biomass was collected in May 2019 from El-Mahmoudia Canal (31°3007′ N and 29°4166′ E), using a standard plankton net (No. 25) of 55 μm mesh size with a diameter of 55 cm as described by Abo-Taleb, et al. [
23], Manickam, et al. [
34]. The collected freshwater zooplankton biomass consisted of 48.70% Rotifera, 46.10% Copepoda, 3.09% Cladocera, 1.21% Ostracoda and 0.91% Protozoa. The obtained biomass was cleaned, washed with distilled water, dried at 50 °C for 24 h, and finely ground using an electric mill. Approximate composition of ZBM and ingredients of the experiment diets are presented in
Table 1. The control diet (Z
0) was formulated with using FM as animal protein source (200 g/kg), then the FM was replaced by ZBM at the ratios of 25%, 50%, 75%, and 100% (Z
25, Z
50, Z
75, and Z
100), respectively, to form five iso-energetic and iso-nitrogenous experimental diets.
All the ingredients were finely mashed and sieved through a 60-mesh sieve before being completely combined. To form a dough, water and oil were progressively added to the dry ingredients and the mixture was pelleted using a meat grinder. Finally, pellets were dried and crushed at 50 °C for 24 h, sieved to generate feed particles with an average size of 0.5 mm, and stored at −20 °C. The diet formulation and approximate composition are shown in
Table 2.
2.3. Growth Performance and Survival Determinations
Prior to the start of the growth trial, 50 fish were randomly collected to measure the initial weight and length. After 60-days, all the fish (after having fasted overnight) were taken from the aquarium with a dip net and gently anesthetized with 50 mg of clove oil l
−1; then, their wet BW (g) and SL (cm) determined to the nearest 1.0 mg and 1.0 mm, respectively. Ten fish from each replicate per treatment were directly stored at −20 °C to analyze whole-body composition. In addition, 3 specimens per replicate were sacrificed with an overdose of anesthetic for assessing the histological changes in the liver and intestine. Survival rate (SR), weight gain (WG), daily growth index (DGI), thermal growth coefficient (TGC), length gain (LG), feed conversion ratio (FCR), protein efficiency ratio (PER), lipid efficiency ratio (LER), and condition factor (CF) were calculated using the following formulae:
2.4. Proximate Chemical Analysis
All chemical analyses of ingredients, experimental diets, and fish were examined by the standard methods of AOAC [
35]. The whole fish that had been sampled before and after the experiment were ground and pooled. The moisture content of the samples was evaluated by drying them at 105 °C for 24 h. Before the chemical analysis, frozen body samples were freeze-dried. The ash content of feed and whole-body fish was determined using a muffle furnace at 550 °C for 5 h (Nabertherm B150, Bremen, Germany). Crude protein (N × 6.25) was determined using the Kjeldahl system method (VELP Scientifica, UDK 149, Usmate Velate, Italy). Crude lipid content was determined by petroleum ether extraction (40–60 °C) using a Soxhlet System (VELP Scientifica, SER 148, Usmate Velate, Italy).
2.5. Histological Examination of the Liver and Midgut
Hepatopancreas and midgut samples of six fish per treatment were selected randomly, then dissected and fixed in 10% buffered formalin solution for 24 h; then, they were dehydrated in ethanol, treated with xylene, embedded in paraffin wax, sectioned into 5 µm sections using a microtome, and stained with haematoxylin and eosin (H&E), according to conventional histological techniques [
36]. The examination was carried out using an Olympus IX71 light microscope and a digital camera (C-4000 zoom) (Olympus Scientific Solutions Americas Inc., Waltham, MA, USA). Villus height, villus width, muscle thickness, and crypt depth were all measured in sections of the intestine according to Hamidian, et al. [
37].
2.6. Economic Evaluation
An economic evaluation of the five experimented diets was calculated, including the cost of feed and the price of fish. The cost of experimental diets was calculated in Egyptian pounds (LE) according to international and local market prices, per kg, as follows: fishmeal, LE 40.00; zooplankton biomass meal, LE 20.00; soybean meal, LE 8.50; corn meal, LE 3.70; rice polishing, LE 4.00; fish oil, LE 27.00; vitamin and mineral premix, LE 30.00. As the zooplankton biomass meal was collected from nature, its cost was calculated based on the cost of collection, cleaning, drying, grinding, and sieving.
2.7. Statistical Analysis
The SPSS version 25.0 software was used to analyze the data using one-way analysis of variance, followed by Tukey’s honestly significant difference test. If there were significant differences between the groups, Tukey’s multiple comparison test was performed. Mean differences were considered significant at a
p-value equal to or less than 0.05. All data are presented as mean ± standard error of the mean. In addition, to determine the fit regression model between increasing ZBM levels and different measurements, orthogonal quadratic contrasts were performed [
38].
4. Discussion
The world production of zooplankton biomass represents hundreds of millions of tones. Only a small portion of this biomass is harvested and the remainder is not used in commercial products [
17,
39,
40]. However, ZBM contains a high percentage of protein, reaching 45–52% crude protein on dry matter basis [
41,
42,
43]. Therefore, the high protein content of ZBM indicates that it may be used as an alternative animal protein source in the diet of farmed fish [
44], if we farm zooplankton. According to our knowledge, until now, there is no practical diet for grey mullet juveniles that contains ZBM as a total or partial replacement of FM.
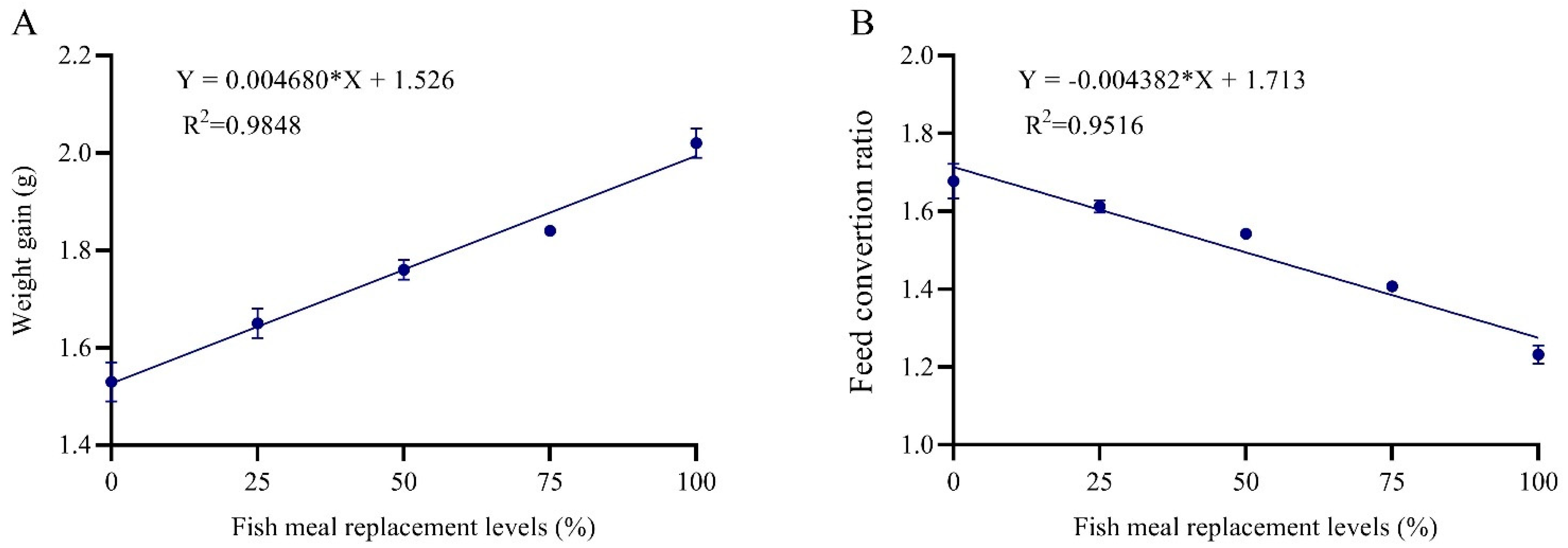
The current study clearly confirmed that ZBM can totally replace FM in a practical diet for grey mullet juveniles. When compared to fish fed the FM-based diet, fish fed increasing levels of ZBM based-diets (Z
25, Z
50, Z
75, and Z
100) showed a trend toward better growth performance, feed consumption, carcass composition, and lower diet preparation expense (Z0). The results of the present study concluded that fish fed Z
100 showed the highest WG, LG, DGI, FER, and survival, while achieved the lowest significant FCR value, compared with Z
0 and other treatments. This data clearly confirms that 100% substitution of FM by ZBM is the ideal replacement of FM for grey mullet juveniles. The result was similar to the observation of Hongxia, et al. [
45], who found that a 60% replacement level of FM by daphnia magna meal significantly increased the weight gain, specific growth rate, and protein efficiency of
Pelteobagrus Fulvidraco fingerlings. Other studies have also found that replacing FM (control diet contained 35% FM) with zooplankton meal up to 100% significantly improved growth performance, PER, and FCR of seabass,
Dicentrarchus labrax, fingerlings [
31]. In addition,
Dahpnia magna meal as an FM alternative in the diet (control diet contained 18% FM) of grey mullet larvae could replace up to 75% of FM, which recorded the best growth and feed utilization parameters [
33]. The barramundi,
Lates calcarifer, fingerlings fed diets containing 5–10% daphnia meal have high immune surveillance and disease resistance compared to the control group [
46]. The study of Sharahi, et al. [
32] revealed that the replacement of FM with gammarus meal up to 20% significantly improved different growth performance criteria and feed utilization of Siberian sturgeon,
Acipenser baerii, juveniles.
In addition, Saravanan, et al. [
47] found that, when feeding young yellow-tail damselfish,
Neopomocentrus nemurus, larvae on copepod, they had better survival and growth performance. A previous study investigated the effect of a mixed copepod and rotifer diet on the survival and growth performance of first-feeding larvae of the southern flounder,
Paralichthys lethostigma, larvae [
48]. In addition, growth enhancement effect of zooplankton has been shown in several fish species, such as Atlantic cod,
Gadus morhua, larvae [
43,
49] and rainbow trout,
Oncorhynchus mykiss, juvenile [
50].
For aquatic animals, zooplankton is a good source of protein, lipids, and carbohydrates [
51]. In the present study, the results of whole-body composition indicated that feeding an increasing ZBM significantly increased crude protein and decreased lipid and ash content of grey mullet. The current findings are consistent with those of Aman, et al. [
52], Manickam, et al. [
53], who found a substantial increase in protein, lipid, and carbohydrate content in freshwater prawn,
Macrobrachium rosenbergii, fed with wild mixed zooplankton. Previously, it was confirmed that
Catla catla fry fed with different zooplankton, such as Copepoda,
Thermocyclops decipiens, Crustacea,
Moina micrura, and
Daphnia spp., had higher protein, carbohydrate, and lipid content [
20,
54]. Abo-Taleb, et al. [
33] reported that the use of
Dahpnia magna meal replacing FM at a level of 75% significantly improved the protein and lipid contents of grey mullet. Carcass protein, lipid, and nutrient digestibility were improved in the juvenile of Australian freshwater crayfish,
Cherax destructor, fed on different protein sources (meat, snail, soybean, yabby, and zooplankton) [
55].
The study of the regression trend of the present results showed a significant linear regression among increasing levels of ZBM and different growth, feed utilization, and body composition criteria. Accordingly, ZBM could totally replace FM without negative impacts. In contrast, daphia meal as an FM replacer showed a quadratic regression trend and the best growth performance criteria were observed at 75% in the diet of grey mullet [
33].
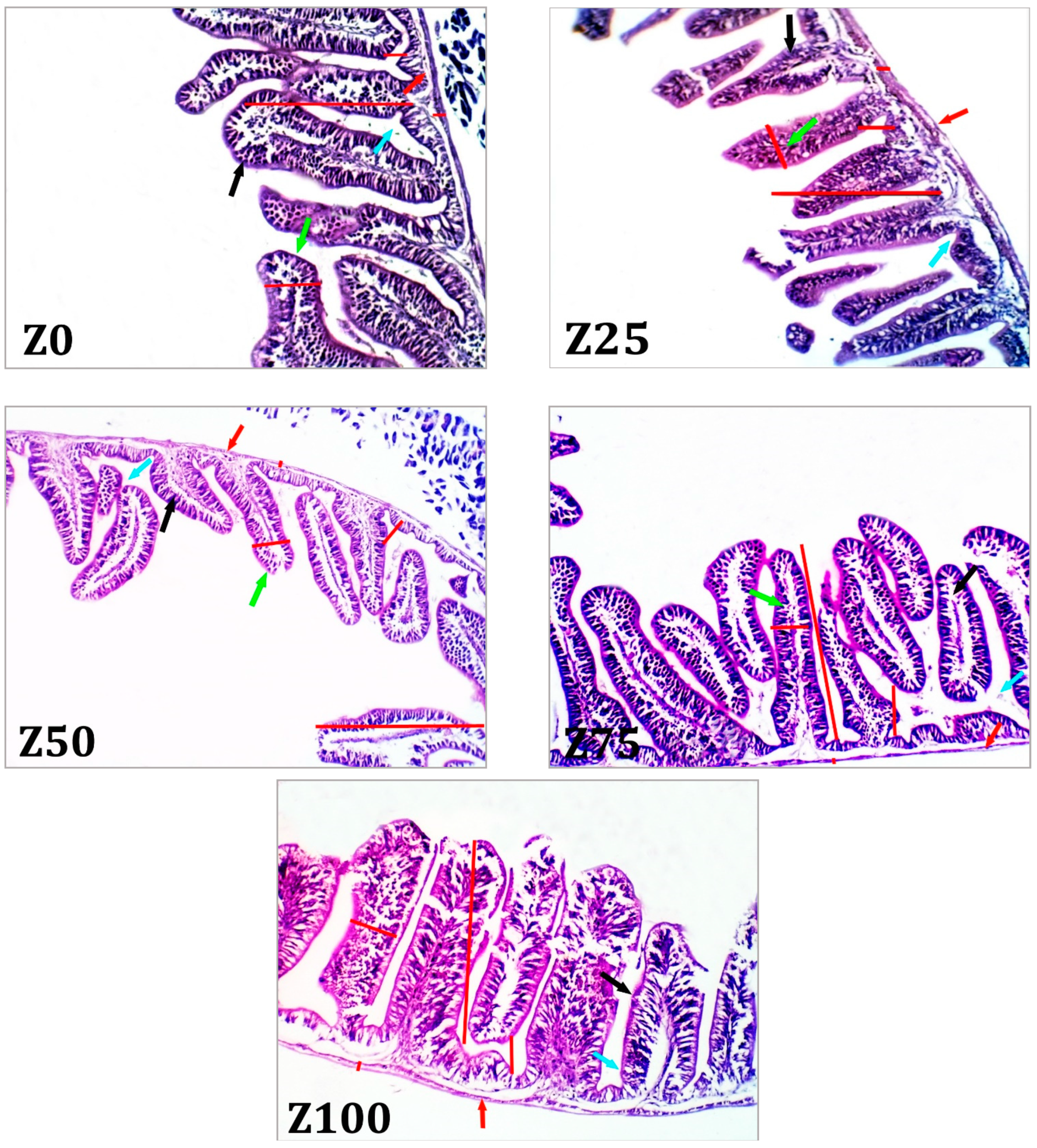
Since the GI tract is the primary site for feed digestion and nutrient absorption, its morphology can be used to assess intestinal health, feed consumption ability, and fish growth [
56,
57,
58]. For normal intestinal functions, intestinal morphology must be healthy [
59]. Some improvements in the structure of the intestine, such as deeper crypts, number of villi and shorter villi, can result in poor nutrient preoccupation and less function [
60,
61]. In the present study, the results showed that zooplankton meal improved the intestinal villi, muscle thickness, and number of goblet cells in the intestine of grey mullet. Similar results were also reported by Mona, et al. [
62] for gilthead seabream,
Sparus aurata, fry when fed marine rotifers and copepods. In addition, Dimitroglou, et al. [
63] reported that the improvement of intestinal morphology by increasing villus length and density and increasing surface area of absorption boost the function of the intestine. In the present study, the morphological results indicate that dried zooplankton at 100% improved the gut development of grey mullet, which may also partially explain the rapid growth of fish fed this diet.
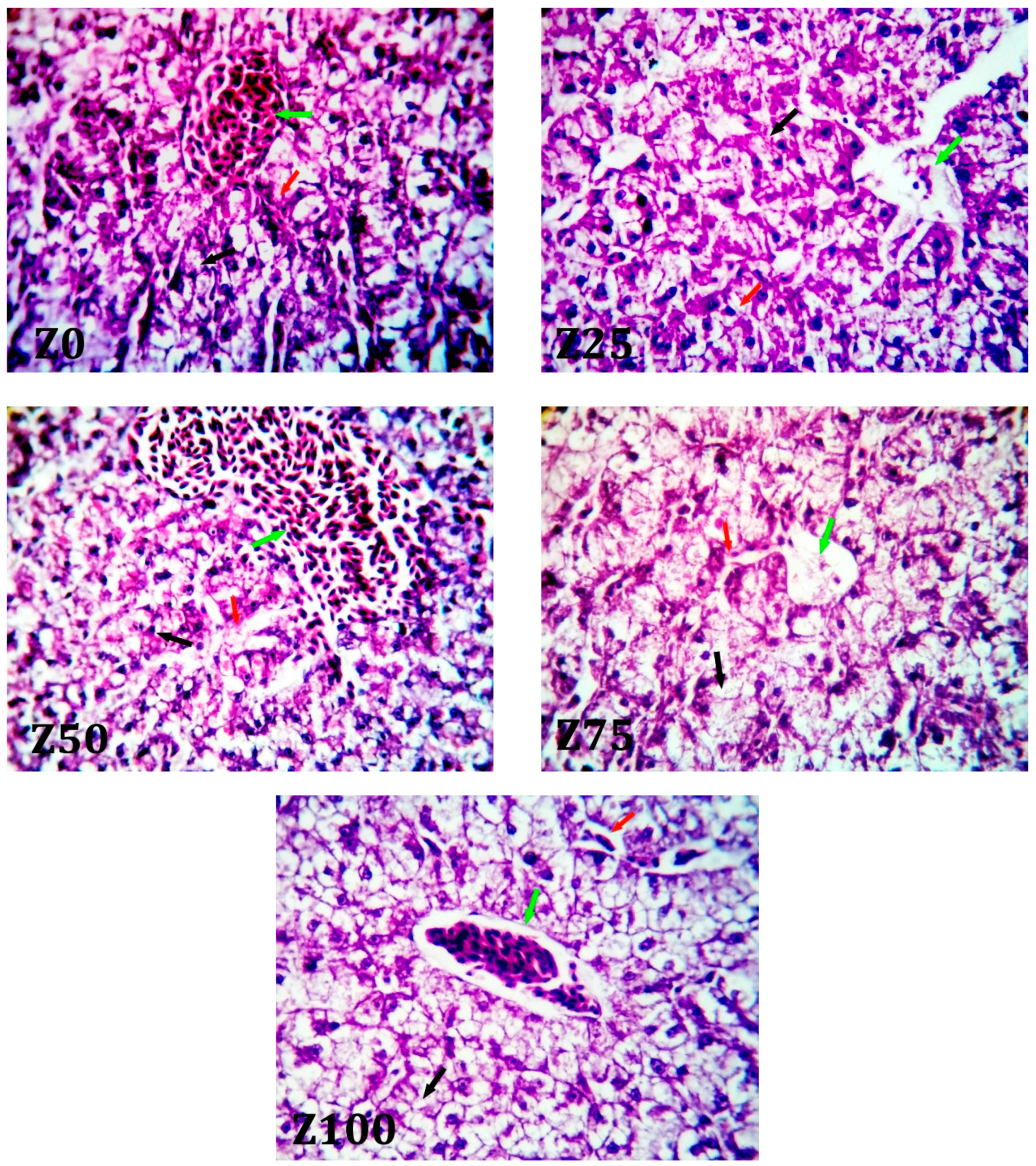
The liver of fish is a vital organ that oversees a variety of life functions, including protein, lipid, and carbohydrate metabolism, bile production, and detoxification. It also serves as a storage facility for a variety of compounds, mostly glycogen and lipids [
64]. In the current study, when grey mullet was fed various levels of ZBM, no histopathological changes were seen, when compared with the control diet (Z0). These results are in agreement with [
65], who showed that there was no effect, in control and fish oil, on liver functions.
Aside from the biological and nutritional assessment of dietary FM substitution, an economic study on alternate ingredient usage is critical to ensure the viability and economics of alternative schemes. The cost of diet preparation was reduced when FM was replaced with ZBM in this study, which is a clear reason for using this ingredient in commercial diets. The cost of weight gain by the diet containing 100% ZBM (9.46 L.E per kg gain) was less costly than the control diet by 40%, resulting in a higher economic efficiency ratio. Overall, this research study found that dietary FM could be replaced by ZBM up to 100% in grey mullet juvenile diets, with positive effects on growth performance and feed consumption, thus improving economic efficiency. In the same vein, the replacement of FM with Daphnia meal improved the ECR by 14.88% and 15.91% with 75 and 100% replacement levels, respectively [
33]. One of the greatest impacts of replacement studies is the improvement of economic visibility of the diet, among other goals. Several alternatives have been used in partial replacement of FM and reduce feed costs, including high protein distillers dried grains [
66,
67], meat and bone meal [
68], silkworm (Anaphe infracta) [
69], insect meal [
70], animal by-products [
70], and whey protein [
71]. Interestingly, the findings of the present study revealed the success of total replacement of FM with ZBM regarding the zootechnical performance and economic benefit by considering the natural collection of ZBM. This economic analysis could have differed if we had farmed the zooplankton with expectation of increasing their price. It is worth mentioning that the control diet in the current study contains high plant protein source (42% soyabean); therefore, other studies could be testing the total replacement of FM with ZBM at high animal protein diets.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}