
Effects of Single or Combined Administration of Dietary Synbiotic and Sodium Propionate on Humoral Immunity and Oxidative Defense, Digestive Enzymes and Growth Performances of African Cichlid (Labidochromis lividus) Challenged with Aeromonas hydrophila


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Feeding Trial
2.3. Growth Indices
2.4. Immunological and Antioxidant Parameter Analyses
2.5. Digestive Enzyme Analyses
2.6. Microbiota Assays
2.7. Proximate Analysis
2.8. Bacterial Exposure Challenge
2.9. Statistical Analysis
3. Results
3.1. Growth Performance and Survival Rate
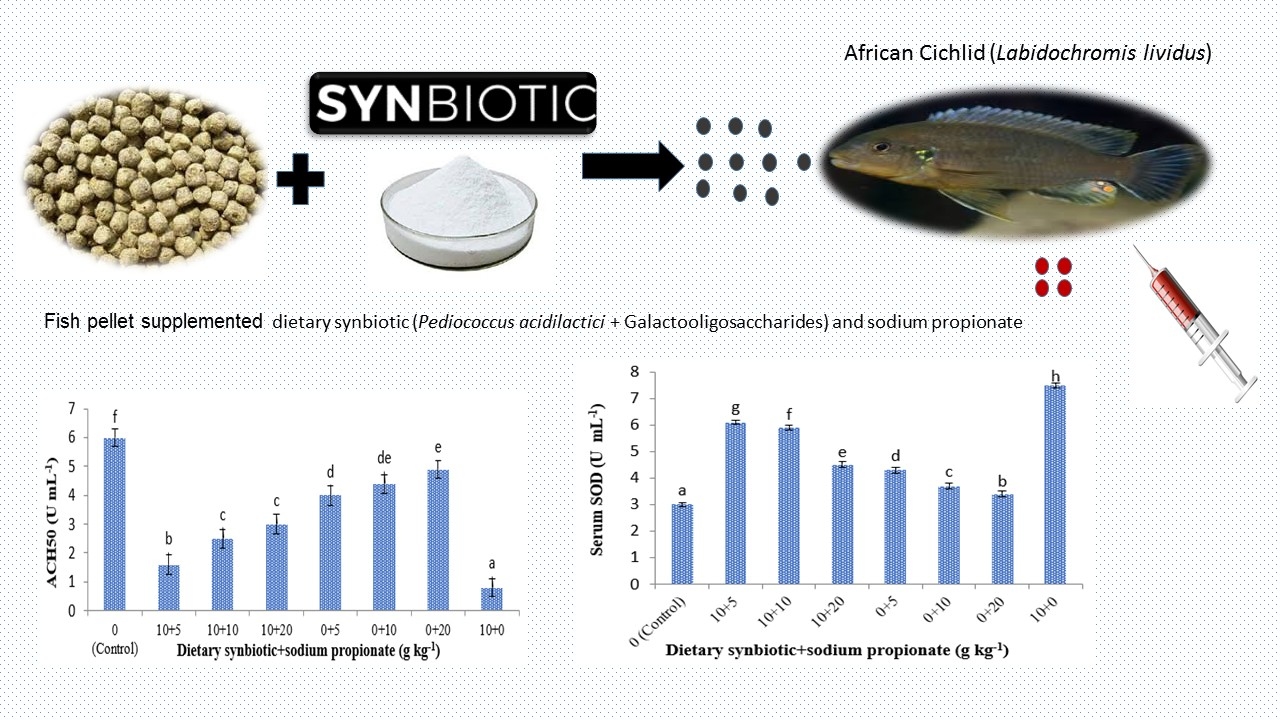
3.2. Immunological Assay
3.3. Antioxidant Enzyme Assay
3.4. Digestive Enzymes Activity
3.5. Microbiota Assay
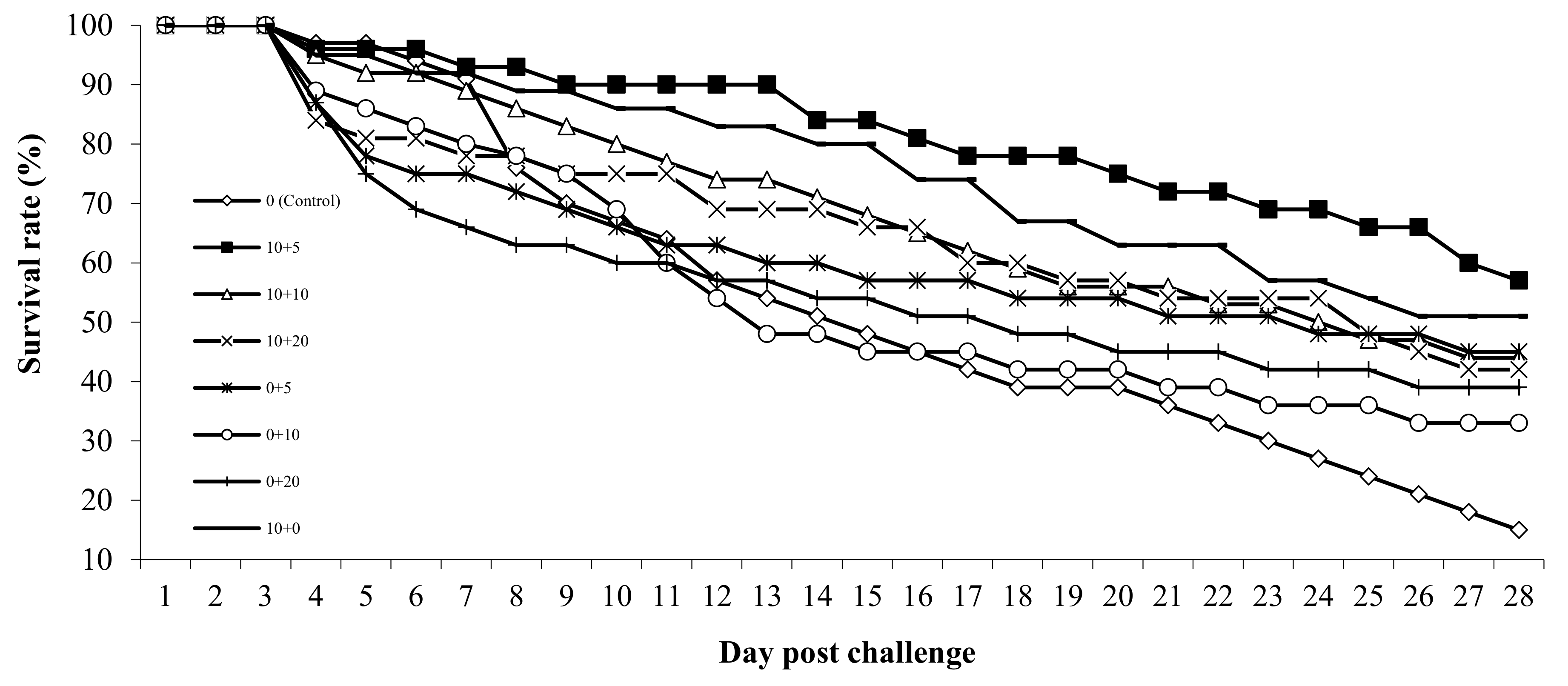
3.6. Pathogen Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ladisa, C.; Bruni, M.; Lovatelli, A. Overview of Ornamental Species Aquaculture. FAO Aquac. Newsl. 2017, 56, 39–40. [Google Scholar]
- Ngo, V.H. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture 2015, 446, 88–96. [Google Scholar]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish. Shellfish. Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef]
- Safari, O.; Paolucci, M.; Ahmadniaye Motlagh, H.R. Effects of synbiotics on immunity and disease resistance of narrow-clawed crayfish, Astacus leptodactylus leptodactylus (Eschscholtz, 1823). Fish. Shellfish Immunol. 2017, 64, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Safari, O.; Paolucci, M. Effect of in vitro selected synbiotics (galactooligosaccharide and mannanoligosaccharide with or without Enterococcus faecalis) on growth performance, immune responses and intestinal microbiota of juvenile narrow clawed crayfish, Astacus leptodactylus leptodactylus Eschscholtz, 1823. Aquac. Nutr. 2018, 24, 247–259. [Google Scholar]
- Safari, O.; Sarkheil, M. Dietary administration of eryngii mushroom (Pleurotus eryngii) powder on haemato-immunological responses, bactericidal activity of skin mucus and growth performance of koi carp fingerlings (Cyprinus carpio koi). Fish. Shellfish Immunol. 2018, 80, 505–513. [Google Scholar] [CrossRef]
- Wang, Y.B.; Tian, Z.; Yao, J.; Li, W. Effect of probiotics, Enterococcus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture 2008, 277, 203–207. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bogwald, J. The current status and future focus of probiotic and prebiotic applications for salmo¬nids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Ghasempour Dehaghani, P.; Javaheri Baboli, M.; Taghavi Moghadam, A.; Ziaei-Nejad, S.; Pourfarhadi5, M. Effect of synbiotic dietary supplementation on survival, growth performance, and digestive enzyme activities of common carp (Cyprinus carpio) fingerlings. Czech. J. Anim. Sci. 2015, 60, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Azimirad, M.; Meshkini, S.; Ahmadifard, N.; Hoseinifar, S.H. The effects of feeding with synbiotic (Pediococcus acidilactici and fructooligosaccharide) enriched adult Artemia on skin mucus immune responses, stress resistance, intestinal microbiota and performance of angelfish (Pterophyllum scalare). Fish. Shellfish Immunol. 2016, 54, 516–522. [Google Scholar] [CrossRef]
- Safari, R.; Hoseinifar, S.H.; Nejadmoghadam, S.; Khalili, M. Non-specific immune parameters, immune, antioxidant and growth-related genes expression of common carp (Cyprinus carpio L.) fed sodium propionate. Aquac. Res. 2017, 48, 1–9. [Google Scholar] [CrossRef]
- Fuller, R. Probiotic in man and animals. J. Appl. Microbiol. 1989, 66, 365–378. [Google Scholar]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Lactobacillus sakei BK19 enriched diet enhances the immunity status and disease resistance to streptococcosis infection in kelp grouper, Epinephelus bruneus. Fish. Shellfish Immunol. 2010, 29, 1037–1043. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspec¬tive. Fish. Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Bradley, G.; Harper, G.M.; Baker, R.T.M.; Munn, C.B.; Davies, S.J. Assessment of the effects of vegetative and lyophilized Pediococcus acidilactici on growth, feed utilization, intestinal colonization and health parameters of rainbow trout (Oncorhynchus mykiss Walbaum). Aquac. Nutr. 2011, 17, 73–79. [Google Scholar] [CrossRef]
- Neissi, A.; Rafiee, G.; Nematollahi, M.; Safari, O. The effect of Pediococcus acidilactici bacteria used as probiotic supplement on the growth and non-specific immune responses of green terror, Aequidens rivulatus. Fish. Shellfish Immunol. 2013, 35, 1976–1980. [Google Scholar] [CrossRef]
- Safari, R.; Hoseinifar, S.H.; Kavandi, M. Modulation of antioxidant defence and immune response in zebra fish (Danio rerio) using dietary sodium propionate. Fish. Physiol. Biochem. 2016, 42, 1733–1739. [Google Scholar] [CrossRef] [PubMed]
- Al-Hisnawi, A.; Rodiles, A.; Rawling, M.D.; Castex, M.; Waines, P.; Gioacchini, G.; Carnevali, O. Dietary robiotic Pediococcus acidilactici MA18/5M modulates the intestinal microbiota and stimulates intestinal immunity in rainbow trout (Oncorhynchus mykiss). J. World Aquacult Soc. 2019, 50, 1–19. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the colonic microbiota: Introducing the concept of prebiotics. Nutr. J. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Guerreiro, I.; Couto, A.; Machado, M.; Castro, C.; Pousão-Ferreira, P.; Oliva-Teles, A. Prebiotics effect on immune and hepatic oxidative status and gut morphology of white sea bream (Diplodus sargus). Fish. Shellfish Immunol. 2016, 50, 168–174. [Google Scholar] [CrossRef]
- Ringo, E.; Song, S.K. Application of dietary supplements (synbiotics and probiotics in combination with plant products and β-glucans) in aquaculture. Aquac. Nutr. 2016, 22, 4–24. [Google Scholar] [CrossRef]
- Yousefi, S.; Hoseinifar, S.H.; Paknejad, H.; Hajimoradloo, A. The effects of dietary supplement of galactooligosaccharide on innate immunity, immune related genes expression and growth performance in zebrafish (Danio rerio). Fish. Shellfish Immunol. 2018, 73, 192–196. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Doan, H.V.; Ashouri, G. Galactooligosaccharide effects as prebiotic on intestinal microbiota of different fish species. J. Agron. Anim. Ind. 2019, 14, 266–278. [Google Scholar] [CrossRef]
- Ng, W.K.; Koh, C.B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquacult. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Ahmadniaye Motlagh, H.; Sarkhei, M.; Safari, O.; Paolucci, M. Supplementation of dietary apple cider vinegar as an organic acidifier on the growth performance, digestive enzymes and mucosal immunity of green terror (Andinoacara rivulatus). Aquac. Res. 2019, 51, 1–9. [Google Scholar] [CrossRef]
- Silva, B.C.; Vieira, F.N.; Mourino, J.L.P.; Ferreira, G.S.; Seiffert, W.Q. Salts of organic acids selection by multiple characteristics for marine shrimp nutrition. Aquaculture 2013, 384–387, 104–110. [Google Scholar] [CrossRef]
- Erdogan, F.; Erdogan, M.; Gümüş, E. Effects of dietary protein and lipid levels on growth performances of two African Cichlids (Pseudotropheus socolofi and Haplochromis ahli). Turk. J. Fish. Aquat. Sci. 2012, 12, 635–640. [Google Scholar]
- Sarkheil, M.; Ameri, M.; Safari, O. Application of alginate-immobilized microalgae beads as biosorbent for removal of total ammonia and phosphorus from water of African cichlid (Labidochromis lividus) recirculating aquaculture system. Environ. Sci. Pollut. Res. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Siwicki, A.K.; Anderson, D.P.; Rumsey, G.L. Dietary intake of immunostimulants by rainbow trout affects non-specific immunity and protection against furunculosis. Vet. Immunol. Immunopathol. 1994, 41, 125–139. [Google Scholar] [CrossRef]
- Kumari, J.; Sahoo, P.K.; Swain, T.; Sahoo, S.K.; Sahu, A.K.; Mohanty, B.R. Seasonal variation in the innate immune parameters of the Asian catfish Clarias batrachus. Aquaculture 2006, 252, 121–127. [Google Scholar] [CrossRef]
- Yano, T. Assay of Hemolytic Complement Activity. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Hattari, S.C., Rowley, A.F., Eds.; SOS Publications: Trenton, NJ, USA, 1992; pp. 131–141. [Google Scholar]
- Yanbo, W.; Zirong, X. Effect of probiotics for common carp (Cyprinus carpio) based on growth performance and digestive enzyme activities. Anim. Feed Sci. Technol. 2006, 127, 283–292. [Google Scholar] [CrossRef]
- Gawlicka, A.; Parent, B.; Horn, M.H.; Ross, N.; Opstad, I.; Torrissen, O.J. Activity of digestive enzymes in yolk-sac larvae of Atlantic halibut (Hippoglossus hippoglossus): Indication of eadiness for first feeding. Aquaculture 2000, 184, 303–314. [Google Scholar] [CrossRef]
- Worthington, C. Worthington Enzyme Manual Related Biochemical. Freehold. N. J. 1991, 34, 212–215. [Google Scholar]
- Hidalgo, M.C.; Urea, E.; Sanz, A. Comparative study of digestive enzymes in fish with different nutritional habits: Proteolytic and amylase activities. Aquaculture 1999, 170, 267–283. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists Arlington: Washington, DC, USA, 2005. [Google Scholar]
- Hoseinifar, S.H.; Zoheiri, F.; Caipang, C.M. Dietary sodium propionate improved performance, mucosal and humoral immune responses in Caspian white fish (Rutilus frisii kutum) fry. Fish. Shellfish Immunol. 2016, 55, 523–528. [Google Scholar] [CrossRef]
- Sewaka, M.; Trullas, C.; Chotiko, A.; Rodkhum, C.; Chansue, N.; Boonanuntanasarn, S.; Pirarat, N. Efficacy of synbiotic Jerusalem artichoke and Lactobacillus rhamnosus GG-supplemented diets on growth performance, serum biochemical parameters, intestinal morphology, immune parameters and protection against Aeromonas veronii in juvenile red tilapia (Oreochromis spp.). Fish. Shellfish Immunol. 2018, 86, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Eslamloo, K.; Falahatkar, B.; Yokoyama, S. Effects of dietary bovine lactoferrin on growth, physiological performance, iron metabolism and non-specific immune responses of Siberian sturgeon Acipenser baeri. Fish. Shellfish Immunol. 2012, 32, 976–985. [Google Scholar] [CrossRef]
- Haroun, E.; Goda, A.; Kabir, M. Effect of dietary probiotic biogen supplementation as a growth promoter on growth performance and feed utilization of Nile tilapia Oreochromis niloticus (L.). Aquac. Res. 2006, 37, 1473–1480. [Google Scholar] [CrossRef]
- Kuuhlmann, K.J.; Jintasataporn, O.; Leucksteadt, C. Dietary potassium-diformate (KDF) improves growth performance of white-leg shrimp Litopenaeus vannamei under controlled conditions. Int. Aquafeed 2011, 3, 19–22. [Google Scholar]
- Rahimnejad, S.; Guardiola, F.A.; Leclercq, E.; Esteban, M.A.; Castex, M.; Sotoudeh, E.; Lee, S.M. Effects of dietary supplementation with Pediococcus acidilactici MA18/5M, galactooligosaccharide and their synbiotic on growth, innate immunity and disease resistance of rockfish (Sebastes schlegeli). Aquaculture 2018, 482, 36–44. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Hoseini, S.M.; Bagheri, D. Effects of galactooligosaccharide and Pediococcus acidilactici on antioxidant defence and disease resistance of rainbow trout, Oncorhynchus mykiss. Ann. Anim. Sci. 2017, 17, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Mirshahpanah, M.; Salari, A.; Shahsavani, D.; Baghshani, H.; Nematy, M. Effect of Pediococcus acidilatici on intestinal microbiota and the oxidative parameters of blood and muscles in Common Carp (Cyprinus carpio). J. Nutr. Fasting Health 2019, 7, 37–43. [Google Scholar] [CrossRef]
- Daniels, C.; Merrifield, D.; Boothroyd, D.; Davies, S.; Factor, J.; Arnold, K. Effect of dietary Bacillus spp. and man¬nan oligosaccharides (MOS) on European lobster (Homarus gammarus L.) larvae growth performance, gut morphology and gut microbiota. Aquaculture 2010, 304, 49–57. [Google Scholar] [CrossRef]
- Arani, M.M.; Salati, A.P.; Safari, O.; Keyvanshokooh, S. Dietary supplementation effects of Pediococcus acidilactici as probiotic on growth performance, digestive enzyme activities and immunity response in zebrafish (Danio rerio). Aquac. Nutr. 2019, 25, 854–861. [Google Scholar] [CrossRef]
- Li, J.S.; Li, J.L.; Wu, T.T. Effects of non-starch polysaccharides enzyme, phytase and citric acid on activities of endogenous digestive enzymes of tilapia (Oreochromis niloticus × Oreochromis aureus). Aquac. Nutr. 2009, 15, 415–420. [Google Scholar] [CrossRef]
- Su, X.; Li, X.; Leng, X.; Tan, C.; Liu, B.; Chai, X. The improvement of growth, digestive enzyme activity and disease resistance of white shrimp by the dietary citric acid. Aquac. Int. 2014, 22, 1823–1835. [Google Scholar] [CrossRef]
- Li, X.; Ringo, E.; Hoseinifar, S.H.; Lauzon, H.; Birkbeck, H.; Yang, D. Adherence and colonisation of microorganisms in the fish gastrointestinal tract. Rev. Aquacult. 2018, 11, 603–618. [Google Scholar] [CrossRef]
- Ringo, E.; Hoseinifar, S.H.; Ghosh, K.; Doan, H.V.; Beck, B.R.; Song, S.K. Lactic Acid Bacteria in Finfish—An Update. Front. Microbiol. 2018, 9, 1818. [Google Scholar] [CrossRef] [PubMed]
- Zare, A.; Azari-Takami, G.; Taridashti, F.; Khara, H. The effects of Pediococcus acidilactici as a probiotic on growth performance and survival rate of great sturgeon, Huso huso (Linnaeus, 1758). Iran. J. Fish. Sci. 2017, 16, 150–161. [Google Scholar]
- Wang, A.R.; Ran, C.; Ringo, E.; Zhou, Z. Progress in fish gastrointestinal microbiota research. Rev. Aquacult. 2018, 10, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Wassef, E.A.; Saleh, N.E.; Abdel-Meguid, N.E.; Barakat, K.M.; Abdel-Mohsen, H.H.; El-bermawy, N.M. Sodium propionate as a dietary acidifier for European seabass (Dicentrarchus labrax) fry: Immune competence, gut microbiome, and intestinal histology benefits. Aquac. Int. 2019, 28, 1–17. [Google Scholar] [CrossRef]
- Owen, M.A.G.; Waines, P.; Bradley, G.; Davies, S. The Effect of Dietary Supplementation of Sodium Butyrate on the Growth and Microflora of Clarias gariepinus (Burchell 1822). In Proceedings of the 12th International Symposium on Fish Nutrition and Feeding, Biarritz, France, 28 May–1 June 2006; p. 149. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feed Ingredient | Content (g kg−1) |
|---|---|
| Fishmeal 1 | 350 |
| Spirulina meal 2 | 20 |
| Soybean meal 1 | 73 |
| Corn gluten 1 | 100 |
| Wheat flour 1 | 240 |
| Fish oil 1 | 70 |
| Canola oil 1 | 70 |
| Soy lecithin 1 | 5 |
| Choline chloride (70%) 3 | 4 |
| Vitamin C (stay) 3 | 5 |
| Vitamin premix 3,* | 15 |
| Mineral premix 3,* | 15 |
| Antifungus 1 | 3 |
| Filler (Carboxymethyl cellulose) 4 | 30 |
| Chemical composition (g kg−1) | |
| Dry matter | 899.39 |
| Crude protein | 421 |
| Crude fat | 282 |
| Crude fiber | 65 |
| Nitrogen free extract | 131.39 |
| Gross energy (Mj kg−1) | 15.58 |
| Dietary Synbiotic + Sodium Propionate Level (g kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0 (Control) | 10 + 5 | 10 + 10 | 10 + 20 | 0 + 5 | 0 + 10 | 0 + 20 | 10 + 0 | |
| Initial weight (g) | 0.65 ± 0.05 | 0.58 ± 0.04 | 0.67 ± 0.11 | 0.69 ± 0.05 | 0.65 ± 0.03 | 0.7 ± 0.02 | 0.68 ± 0.03 | 0.65 ± 0.09 |
| Final weight (g) | 3.49 ± 0.09 a | 3.83 ± 0.09 b | 3.81 ± 0.13 b | 3.82 ± 0.07 b | 3.60 ± 0.52 b | 3.59 ± 0.06 a | 3.63 ± 0.07 a | 3.86 ± 0.04 b |
| Weight gain (g) | 2.83 ± 0.058 a | 3.25 ± 0.13 b | 3.14 ± 0.026 b | 3.13 ± 0.025 b | 2.95 ± 0.50 b | 2.89 ± 0.055 a | 2.94 ± 0.10 a | 3.20 ± 0.11 b |
| (% BW day−1) | 2.65 ± 0.10 a | 2.99 ± 0.16 d | 2.77 ± 0.22 bcd | 2.71 ± 1.02 bc | 2.69 ± 0.19 cd | 2.59 ± 0.03 a | 2.65 ± 0.09 ab | 2.82 ± 0.22 cd |
| Feed conversion ratio | 1.76 ± 0.03 c | 1.46 ± 0.06 b | 1.43 ± 0.01 b | 1.43 ± 0.02 b | 1.16 ± 0.21 a | 1.73 ± 0.03 c | 1.59 ± 0.05 c | 1.24 ± 0.04 a |
| Condition factor | 1.26 ± 0.01 | 1.43 ± 0.07 | 1.62 ± 0.09 | 1.49 ± 0.07 | 1.26 ± 0.1 | 1.26 ± 0.04 | 1.47 ± 0.04 | 1.49 ± 0.3 |
| PER | 1.3 ± 0.14 a | 2.4 ± 0.15 e | 2.2 ± 0.16 de | 2 ± 0.14 cd | 2 ± 0.15 bc | 1.7 ± 0.13 b | 1.4 ± 0.15 a | 2.8 ± 0.14 f |
| PPV | 47 ± 0.14 a | 59 ± 0.16 f | 55 ± 0.17 e | 53 ± 0.2 d | 53 ± 0.18 d | 52 ± 0.19 c | 49 ± 0.19 b | 69 ± 0.14 g |
| Survival rate (%) | 83.33 ± 3.60 a | 97.91 ± 3.60 b | 95.83 ± 7.21 b | 93.75 ± 6.25 b | 97.91 ± 3.60 b | 91.66 ± 9.54 ab | 81.25 ± 12.5 a | 100 ± 0.00 b |
| Dietary Synbiotic + Sodium Propionate Level (g kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0 (Control) | 10 + 5 | 10 + 10 | 10 + 20 | 0 + 5 | 0 + 10 | 0 + 20 | 10 + 0 | |
| Dry matter (%) | 26.88 ± 0.76 | 26.63 ± 0.86 | 26.98 ± 0.47 | 27.08 ± 0.56 | 27.00 ± 0.36 | 26.78 ± 0.56 | 27.05 ± 0.66 | 27.03 ± 0.56 |
| Crude protein (%) | 16.07 ± 0.37 a | 16.95 ± 0.32 f | 16.84 ± 0.42 e | 16.67 ± 0.23 d | 16.69 ± 0.23 d | 16.53 ± 0.23 c | 16.27 ± 0.27 b | 17.04 ± 0.22 g |
| Crude lipid (%) | 5.44 ± 0.05 e | 4.12 ± 0.08 a | 4.60 ± 0.07 c | 4.93 ± 0.13 d | 4.92 ± 0.10 d | 4.89 ± 0.09 d | 5.43 ± 0.09 e | 4.42 ± 0.11 b |
| Ash (%) | 3.16 ± 0.15 a | 3.38 ± 0.32 c | 3.41 ± 0.29 c | 3.39 ± 0.20 c | 3.18 ± 0.21 a | 3.19 ± 0.23 a | 3.25 ± 0.35 b | 3.43 ± 0.22 c |
| Dietary Synbiotic + Sodium Propionate Level (g kg−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0 (Control) | 10 + 5 | 10 + 10 | 10 + 20 | 0 + 5 | 0 + 10 | 0 + 20 | 10 + 0 | |
| Protease | 8.1 ± 0.14 a | 10.2 ± 0.16 d | 9.5 ± 0.17 c | 9.4 ± 0.2 c | 9.3 ± 0.18 b c | 9 ± 0.19 b | 8.4 ± 0.19 a | 10.5 ± 0.16 d |
| α-amylase | 3.9 ± 0.14 a | 7 ± 0.15 g | 6.3 ± 0.16 f | 5.5 ± 0.14 e | 5.2 ± 0.15 d | 4.9 ± 0.13 c | 4.5 ± 0.15 b | 8.2 ± 0.15 h |
| Lipase | 4.3 ± 0.19 a | 6.4 ± 0.2 e | 6.1 ± 0.22 d | 6.4 ± 0.23 c d | 6.1 ± 0.23 b c | 5.9 ± 0.2 b | 4.6 ± 0.23 a | 8.4 ± 0.15 f |
| Bacteria Count (Log CFU g−1) | Dietary Synbiotic + Sodium Propionate Level (g kg−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 (Control) | 10 + 5 | 10 + 10 | 10 + 20 | 0 + 5 | 0 + 10 | 0 + 20 | 10 + 0 | |
| Total aerobic | 6.28 ± 1.09 | 6.20 ± 1.21 | 6.48 ± 1.30 | 6.38 ± 1.47 | 6.41 ± 1.31 | 6.47 ± 1.53 | 6.39 ± 1.52 | 6.48 ± 1.42 |
| Lactobacillus | 0.42 ± 1.13 a | 3.31 ± 1.09 e | 3.12 ± 1.27 d | 2.93 ± 1.35 c | 2.89 ± 1.07 c | 1.67 ± 1.43 b | 1.71 ± 1.29 b | 3.09 ± 1.18 d |
| E. coli count | 0.51 ± 0.32 e | 0.25 ± 0.28 a | 0.43 ± 0.31 d | 0.37 ± 0.21 c | 0.38 ± 0.18 c | 0.31 ± 0.25 b | 0.32 ± 0.26 b | 0.42 ± 0.28 d |
| Fungi count | 0.42 ± 0.43 | 0.39 ± 0.24 | 0.37 ± 0.21 | 0.38 ± 0.24 | 0.41 ± 0.29 | 0.43 ± 0.13 | 0.36 ± 0.19 | 0.36 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safari, O.; Sarkheil, M.; Shahsavani, D.; Paolucci, M. Effects of Single or Combined Administration of Dietary Synbiotic and Sodium Propionate on Humoral Immunity and Oxidative Defense, Digestive Enzymes and Growth Performances of African Cichlid (Labidochromis lividus) Challenged with Aeromonas hydrophila. Fishes 2021, 6, 63. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040063
Safari O, Sarkheil M, Shahsavani D, Paolucci M. Effects of Single or Combined Administration of Dietary Synbiotic and Sodium Propionate on Humoral Immunity and Oxidative Defense, Digestive Enzymes and Growth Performances of African Cichlid (Labidochromis lividus) Challenged with Aeromonas hydrophila. Fishes. 2021; 6(4):63. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040063
Chicago/Turabian StyleSafari, Omid, Mehrdad Sarkheil, Davar Shahsavani, and Marina Paolucci. 2021. "Effects of Single or Combined Administration of Dietary Synbiotic and Sodium Propionate on Humoral Immunity and Oxidative Defense, Digestive Enzymes and Growth Performances of African Cichlid (Labidochromis lividus) Challenged with Aeromonas hydrophila" Fishes 6, no. 4: 63. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes6040063