Interactions between L. monocytogenes and P. fluorescens in Dual-Species Biofilms under Simulated Dairy Processing Conditions


 , ,
, ,  , , ,
, , ,
Abstract
:
1. Introduction
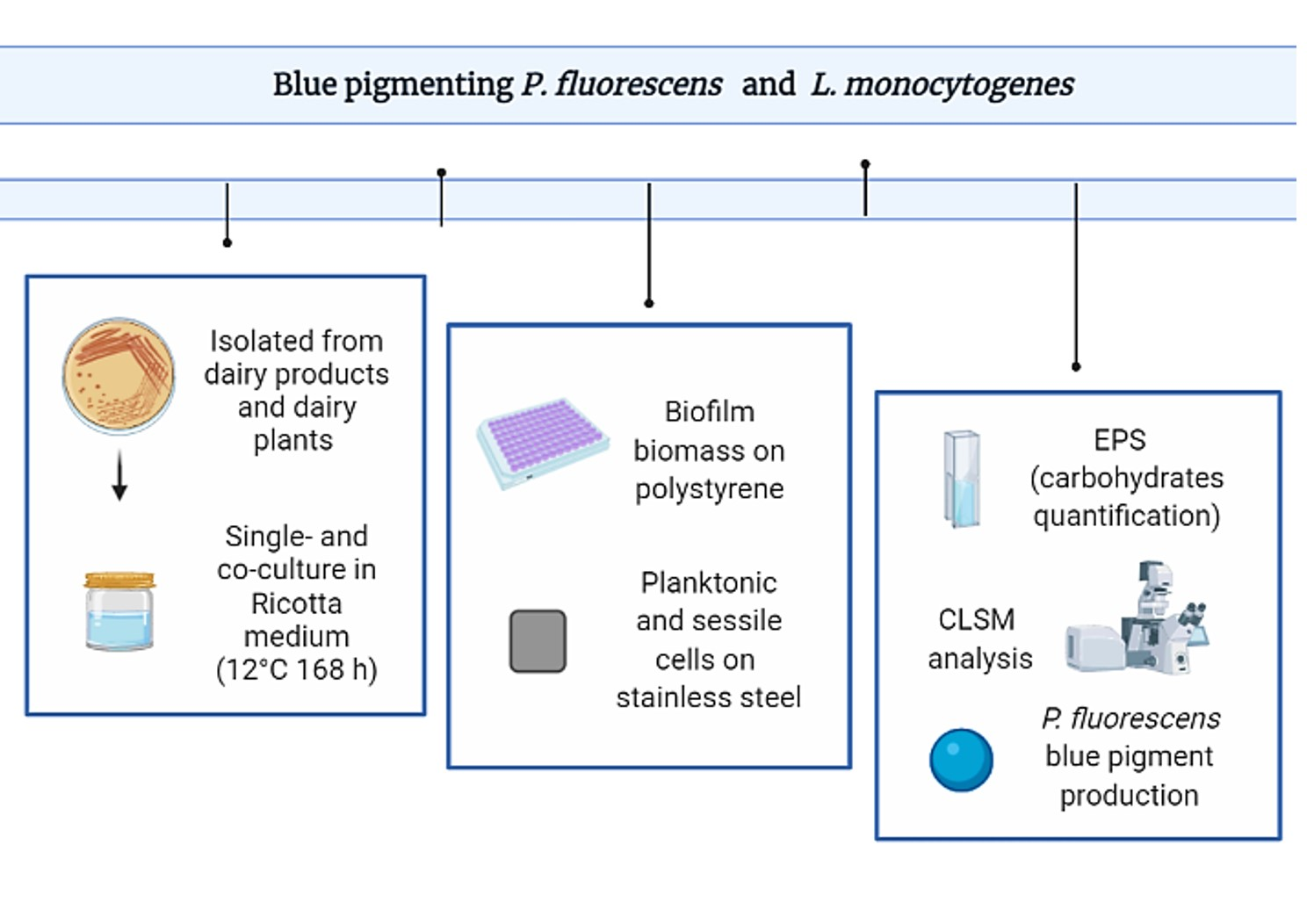
2. Materials and Methods
2.1. Bacterial Strains
2.2. Characterization and Typing of L. monocytogenes Strains
2.3. Inoculum
2.4. Ricotta-Based Medium Preparation
2.5. Biofilm Formation on Polystyrene Microplates
2.6. Biofilm Formation on Stainless Steel and Enumeration of Planktonic and Sessile Cells
2.7. EPS Extraction from Biofilms and Total Carbohydrates Quantification
2.8. Confocal Laser Scanning Microscopy (CLSM) Analysis
2.9. Statistical Analysis
3. Results
3.1. Serotype and Pulsotype of L. monocytogenes Strains
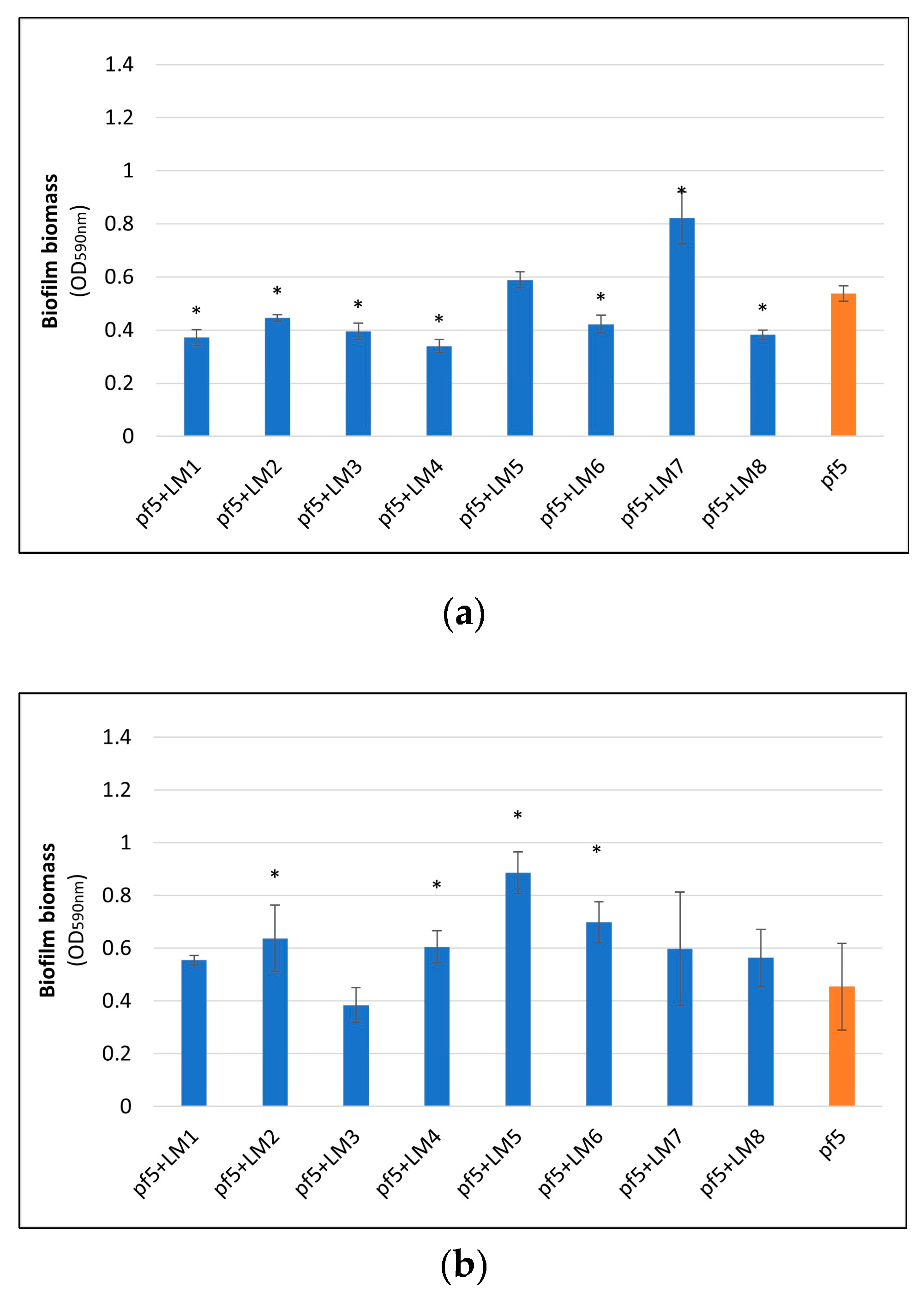
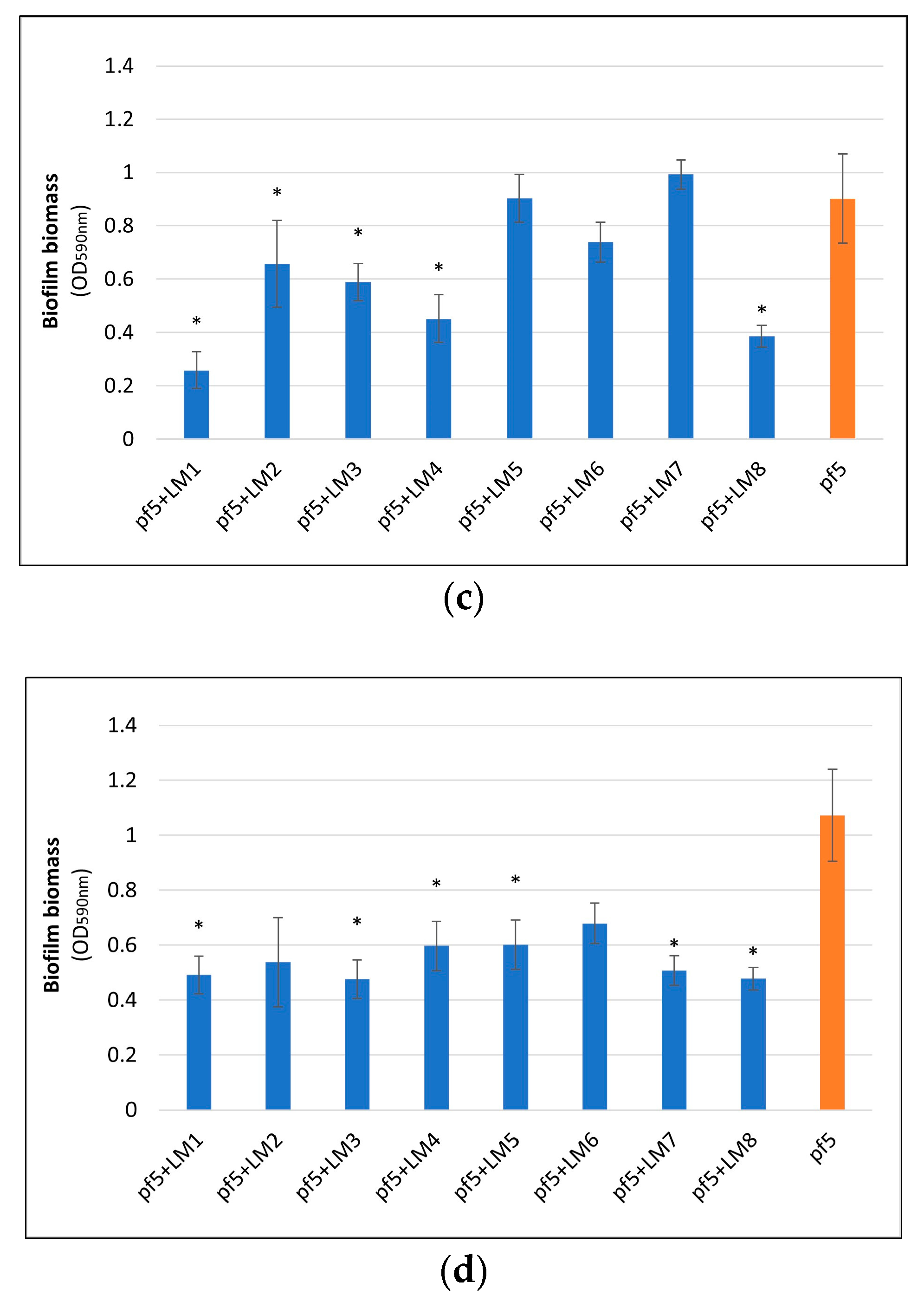
3.2. Biofilm Formation on Polystyrene Surface
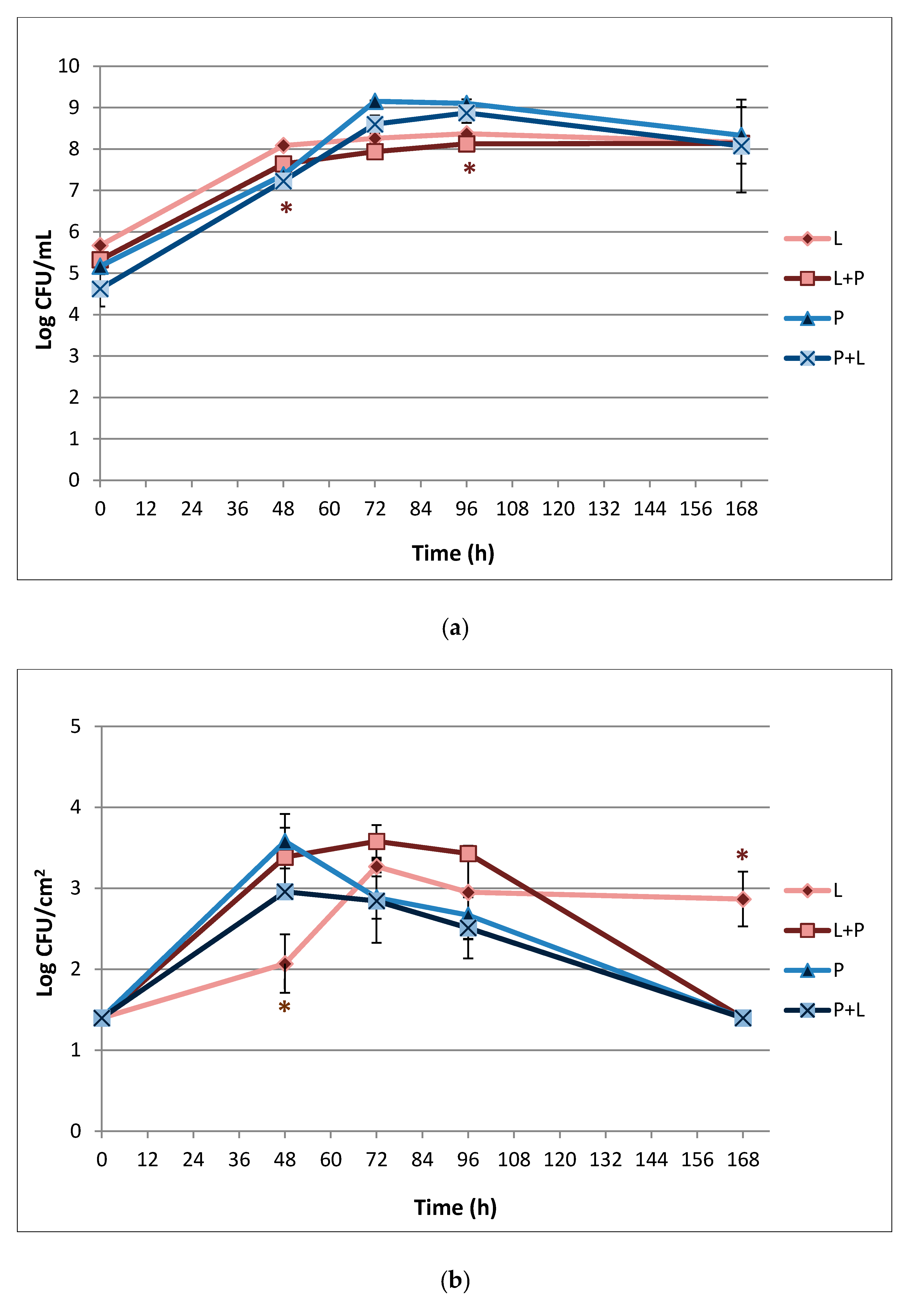
3.3. Biofilm Formation on Stainless Steel Surface and Enumeration of Planktonic and Sessile Cells
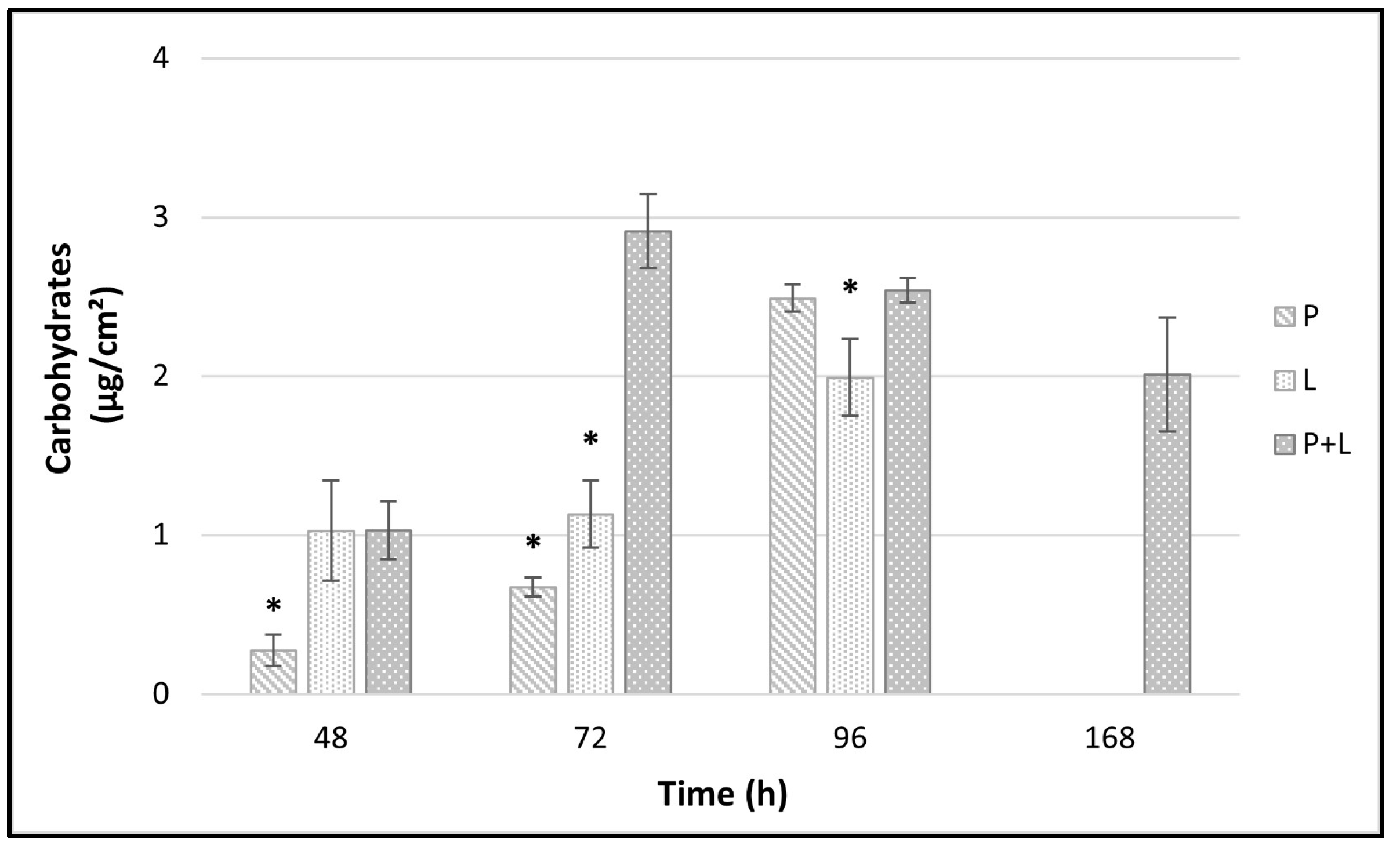
3.4. EPS Analysis by Carbohydrates Quantification
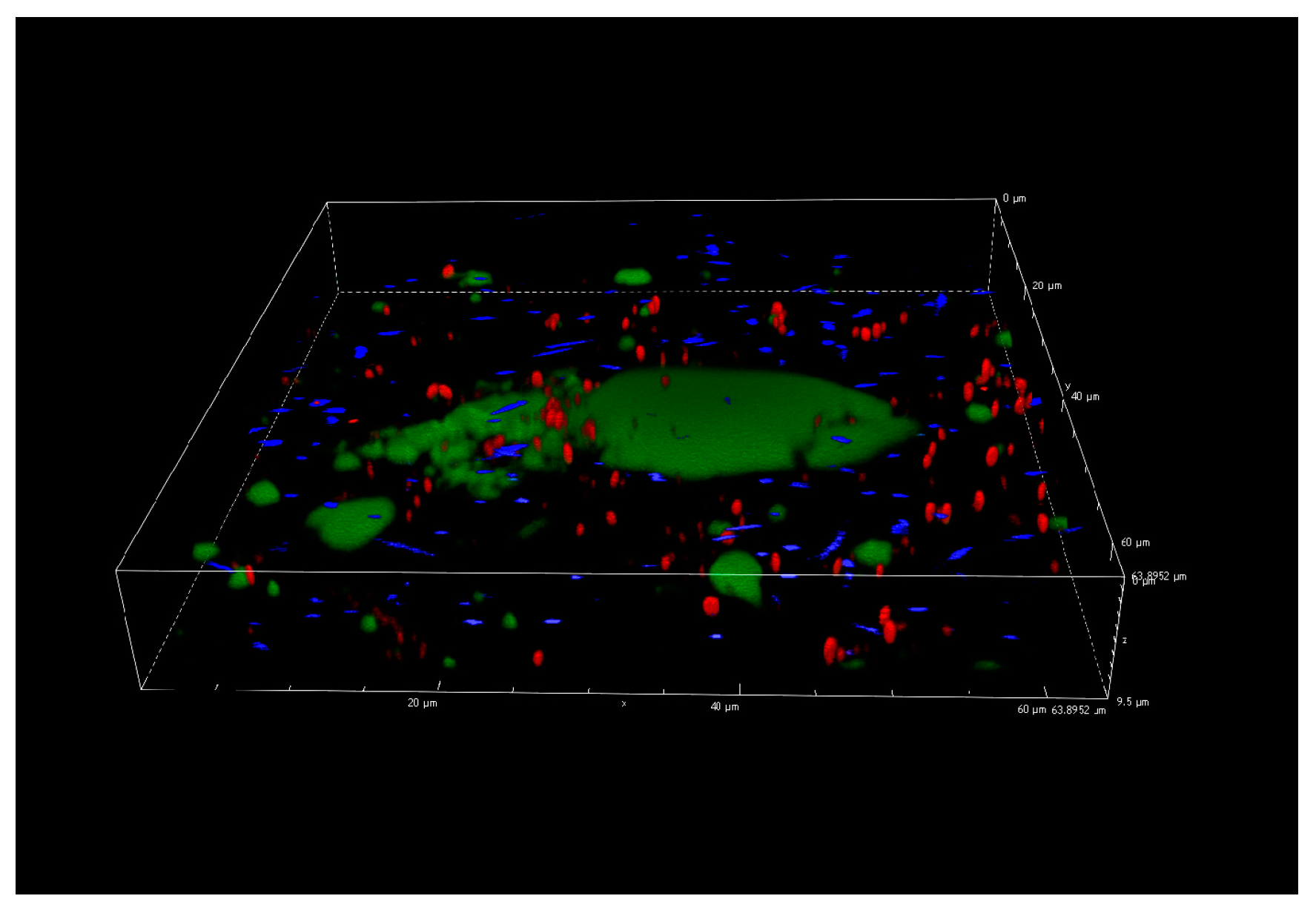
3.5. Confocal Laser Scanning Microscopy Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kassinger, S.J.; Van Hoek, M.L. Biofilm architecture: An emerging synthetic biology target. Synth. Syst. Biotechnol. 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Chaves-López, C.; Serio, A.; Goffredo, E.; Goga, B.T.C.; Paparella, A. Influence of incubation conditions on biofilm formation by Pseudomonas fluorescens isolated from dairy products and dairy manufacturing plants. Ital. J. Food Saf. 2016, 5, 5793. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.S.; Lopes, D.R.G.; Andre, C.; Silva, C.C.; Baglinière, F.; Vanetti, M.C.D. Multispecies biofilm formation by the contaminating microbiota in raw milk. Biofouling 2019, 35, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Di Ciccio, P.; Rubiola, S.; Grassi, M.A.; Civera, T.; Abbate, F.; Chiesa, F. Fate of Listeria monocytogenes in the presence of resident cheese microbiota on common packaging materials. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Melo, J.; Andrew, P.W.; Faleiro, M.I. Listeria monocytogenes in cheese and the dairy environment remains a food safety challenge: The role of stress responses. Food Res. Int. 2015, 67, 75–90. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control, The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500.
- Herrador, Z.; Gherasim, A.; López-Vélez, R.; Benito, A. Listeriosis in Spain based on hospitalisation records, 1997 to 2015: Need for greater awareness. Euro Surveill. 2019, 24, 1800271. [Google Scholar] [CrossRef]
- Carrascosa, C.; Millán, R.; Jaber, J.R.; Lupiola, P.; del Rosario-Quintana, C.; Mauricio, C.; Sanjuán, E. Blue pigment in fresh cheese produced by Pseudomonas fluorescens. Food Control 2015, 54, 95–102. [Google Scholar] [CrossRef]
- Martins, M.; Uelinton, M.P.; Katharina, R.; Vanetti, M.C.D. Milk-deteriorating exoenzymes from Pseudomonas fluorescens 041 isolated from refrigerated raw milk. Braz. J. Microbiol. 2015, 46, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Zarei, M.; Yousefvand, A.; Maktabi, S.; Borujeni, M.P.; Mohammadpour, H. Identification, phylogenetic characterisation and proteolytic activity quantification of high biofilm-forming Pseudomonas fluorescens group bacterial strains isolated from cold raw milk. Int. Dairy J. 2020, 109, 104787. [Google Scholar] [CrossRef]
- Reichler, S.J.; Martin, N.H.; Evanowski, R.L.; Kovac, J.; Wiedmann, M.; Orsi, R.H. A century of gray: A genomic locus found in 2 distinct Pseudomonas spp. is associated with historical and contemporary color defects in dairy products worldwide. Int. J. Dairy Sci. 2019, 102, 5979–6000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Olmo, A.; Calzada, J.; Nuñez, M. The blue discoloration of fresh cheeses: A worldwide defect associated to specific contamination by Pseudomonas fluorescens. Food Control 2018, 86, 359–366. [Google Scholar] [CrossRef]
- Ringel, M.T.; Brüser, T. The biosynthesis of pyoverdines. Microb. Cell. 2018, 5, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Andreani, N.A.; Carraro, L.; Martino, M.E.; Fondi, M.; Fasolato, L.; Miotto, G.; Cardazzo, B. A genomic and transcriptomic approach to investigate the blue pigment phenotype in Pseudomonas fluorescens. Int. J. Food Microbiol. 2015, 213, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Quintieri, L.; Fanelli, F.; Zühlke, D.; Caputo, L.; Logrieco, A.F.; Albrecht, D.; Riedel, K. Biofilm and pathogenesis-related proteins in the foodborne P. fluorescens ITEM 17298 with distinctive phenotypes during cold storage. Front. Microbiol. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Rossi, C.; Serio, A.; Chaves-López, C.; Anniballi, F.; Auricchio, B.; Goffredo, E.; Cenci-Goga, B.T.; Lista, F.; Fillo, S.; Paparella, A. Biofilm formation, pigment production and motility in Pseudomonas spp. isolated from the dairy industry. Food Control 2018, 86, 241–248. [Google Scholar] [CrossRef]
- Paz-Méndez, A.M.; Lamas, A.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Effect of food residues in biofilm formation on stainless steel and polystyrene surfaces by Salmonella enterica strains isolated from poultry houses. Foods 2017, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoët, A.S.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]
- Rossi, C.; Chaves-López, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2020, 1–20. [Google Scholar] [CrossRef]
- Røder, H.L.; Sørensen, S.J.; Burmølle, M. Studying bacterial multispecies biofilms: Where to start? Trends Microbiol. 2016, 24, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, C.; Deschênes, L.; Timothy, C.E.; Bisaillon, Y.; Savard, T. Interactions between spoilage bacteria in tri-species biofilms developed under simulated meat processing conditions. Food Microbiol. 2019, 82, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Shan, L.; Li, X.; Hu, F.; Yuan, Y.; Zhong, D.; Zhang, J. Effects of interspecific interactions on biofilm formation potential and chlorine resistance: Evaluation of dual-species biofilm observed in drinking water distribution systems. J. Water Process. Eng. 2020, 38, 101564. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal biofilm development: Structure, regulation, and treatment strategies. Microbiol. Mol Biol. Rev. 2020, 84, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Dygico, L.K.; Cormac, G.M.; Grogan, G.H.; Burgess, C.M. The ability of Listeria monocytogenes to form biofilm on surfaces relevant to the mushroom production environment. Int. J. Food Microbiol. 2020, 317, 108085. [Google Scholar] [CrossRef] [PubMed]
- Handorf, O.; Pauker, V.I.; Schnabel, U.; Weihe, T.; Freund, E.; Bekeschus, S.; Riedel, K.; Ehlbeck, J. Characterization of antimicrobial effects of plasma-treated water (PTW) produced by microwave-induced plasma (MidiPLexc) on Pseudomonas fluorescens biofilms. Appl. Sci. 2020, 10, 118. [Google Scholar] [CrossRef]
- Haddad, S.; Elliot, M.; Savard, T.; Deschênes, L.; Smith, T.; Ells, T. Variations in biofilms harbouring Listeria monocytogenes in dual and triplex cultures with Pseudomonas fluorescens and Lactobacillus plantarum produced under a model system of simulated meat processing conditions, and their resistance to benzalkonium chloride. Food Control 2020, 107720, in press. [Google Scholar]
- Wang, Y.; Hong, X.; Liu, J.; Zhu, J.; Chen, J. Interactions between fish isolates Pseudomonas fluorescens and Staphylococcus aureus in dual-species biofilms and sensitivity to carvacrol. Food Microbiol. 2020, 91, 1–11. [Google Scholar] [CrossRef]
- Ma, Z.; Stanford, K.; Xiao, M.B.; Yan, D.N.; McAllister, T.A. Effects of beef juice on biofilm formation by Shiga toxin–producing Escherichia coli on stainless steel. Foodborne Pathog. Dis. 2020, 17, 235–242. [Google Scholar] [CrossRef]
- Chen, J.Q.; Regan, P.; Laksanalamai, P.; Healey, S.; Hu, Z. Prevalence and methodologies for detection, characterization and subtyping of Listeria monocytogenes and L. ivanovii in foods and environmental sources. Food Sci. Hum. Well. 2017, 6, 97–120. [Google Scholar] [CrossRef]
- Londero, A.; Costa, M.; Gallia, L.; Brusa, V.; Linaresa, L.; Prieto, M.; Leotta, G. Characterization and subtyping of Listeria monocytogenes strains from butcher shops. LWT Food Sci. Technol. 2019, 113, 108363. [Google Scholar] [CrossRef]
- Donnelly, C.W.; Diez-Gonzalez, F. Guide to foodborne pathogens. In Listeria monocytogenes, 2nd ed.; Labbé, R.G., Garcìa, S., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; pp. 45–74. [Google Scholar]
- Paparella, A.; Mazzarrino, G.; Chaves-López, C.; Rossi, C.; Sacchetti, G.; Guerrieri, O.; Serio, A. Chitosan boosts the antimicrobial activity of Origanum vulgare essential oil in modified atmosphere packaged pork. Food Microbiol. 2016, 59, 23–31. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, R.J.; de Souza, G.T.; Honório, V.G.; de Sousa, J.P.; da Conceição, M.L.; Maganani, M.; de Souza, E.L. Comparative inhibitory effects of Thymus vulgaris L. essential oil against Staphylococcus aureus, Listeria monocytogenes and mesophilic starter co-culture in cheese-mimicking models. Food Microbiol. 2015, 52, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campana, R.; Casettari, L.; Fagioli, L.; Cespi, M.; Bonacucina, G.; Baffone, W. Activity of essential oil-based microemulsions against Staphylococcus aureus biofilms developed on stainless steel surface in different culture media and growth conditions. Int. J. Food Microbiol. 2017, 241, 132–140. [Google Scholar] [CrossRef]
- Dos Santos Rodrigues, J.B.; de Souza, N.T.; Scarano, J.O.A.; de Sousa, J.M.; Lira, M.C.; de Figueiredo, R.C.B.Q.; Magnani, M. Efficacy of using oregano essential oil and carvacrol to remove young and mature Staphylococcus aureus biofilms on food-contact surfaces of stainless steel. LWT 2018, 93, 293–299. [Google Scholar] [CrossRef]
- Abdallah, H.M.I.; Asaad, G.F.M.; Nada, S.A.; Taha, H.S.; Seif El-Nasr, M.M. Influence of extract derived in-vitro cell suspension cultures of Echinacea purpurea against some immunosuppressive effects. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1136–1143. [Google Scholar]
- Harimawan, A.; Ting, Y.P. Investigation of extracellular polymeric substances (EPS) properties of P. aeruginosa and B. subtilis and their role in bacterial adhesion. Colloids Surf. B. 2016, 146, 459–467. [Google Scholar] [CrossRef]
- Rossi, C.; Chaves-López, C.; Serio, A.; Anniballi, F.; Valbonetti, L.; Paparella, A. Effect of Origanum vulgare essential oil on biofilm formation and motility capacity of Pseudomonas fluorescens strains isolated from discoloured Mozzarella cheese. J. Appl. Microbiol. 2018, 124, 1220–1231. [Google Scholar] [CrossRef]
- Martin, N.H.; Murphy, S.C.; Ralyea, R.D.; Wiedmann, M.; Boor, K.J. When cheese gets the blues: Pseudomonas fluorescens as the causative agent of cheese spoilage. Int. J. Dairy Sci. 2011, 94, 3176–3183. [Google Scholar] [CrossRef]
- Braga, V.; Vázquez, S.; Vico, V.; Pastorino, V.; Mota, M.I.; Legnani, M.; Schelotto, F.; Lancibidad, G.; Varela, G. Prevalence and serotype distribution of Listeria monocytogenes isolated from foods in Montevideo-Uruguay. Braz. J. Microbiol. 2017, 48, 689–694. [Google Scholar] [CrossRef]
- Mendez, E.; Walker, D.K.; Vipham, J.; Trinetta, V. The use of a CDC biofilm reactor to grow multi-strain Listeria monocytogenes biofilm. Food Microbiol. 2020, 92, 103592. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Brożek, K.; Łukasik, M.; Wiktorczyk, N.; Korkus, J.; Gospodarek-Komkowska, E. Assessment of drug susceptibility and biofilm formation ability by clinical strains of Listeria monocytogenes. Disaster Emerg. Med. J. 2020, 5, 12–18. [Google Scholar] [CrossRef]
- Hossain, M.I.; Mizana, M.F.R.; Ashrafudoulla, M.; Nahara, S.; Joo, H.J.; Jahid, I.K.; Parkc, S.H.; Kim, K.S.; Ha, S.D. Inhibitory effects of probiotic potential lactic acid bacteria isolated from kimchi against Listeria monocytogenes biofilm on lettuce, stainless-steel surfaces, and MBEC™ biofilm device. LWT Food Sci. Technol. 2020, 118, 108864. [Google Scholar] [CrossRef]
- Puga, C.H.; Dahdouh, E.; SanJose, C.; Orgaz, B. Listeria monocytogenes colonizes Pseudomonas fluorescens biofilms and induces matrix over-production. Front. Microbiol. 2018, 9, 1706. [Google Scholar] [CrossRef] [PubMed]
- Circella, E.; Schiavone, A.; Barrasso, R.; Camarda, A.; Pugliese, N.; Bozzo, G. Pseudomonas azotoformans belonging to Pseudomonas fluorescens Group as causative agent of blue coloration in carcasses of slaughterhouse rabbits. Animals 2020, 10, 256. [Google Scholar] [CrossRef] [Green Version]
- Caputo, L.; Quintieri, L.; Bianchi, D.M.; Decastelli, L.; Monaci, L.; Visconti, A.; Baruzzi, F. Pepsin-digested bovine lactoferrin prevents Mozzarella cheese blue discoloration caused by Pseudomonas fluorescens. Food Microbiol. 2015, 46, 15–24. [Google Scholar] [CrossRef]
- Pang, X.; Yuk, H.G. Effects of the colonization sequence of Listeria monocytogenes and Pseudomonas fluorescens on survival of biofilm cells under food-related stresses and transfer to salmon. Food Microbiol. 2019, 82, 142–150. [Google Scholar] [CrossRef]
- Pang, X.; Wong, C.; Chung, H.J.; Yuk, H.G. Biofilm formation of Listeria monocytogenes and its resistance to quaternary ammonium compounds in a simulated salmon processing environment. Food Control 2019, 98, 200–208. [Google Scholar] [CrossRef]
- Papaioannou, E.; Giaouris, E.D.; Berillis, P.; Boziaris, I.S. Dynamics of biofilm formation by Listeria monocytogenes on stainless steel under mono-species and mixed-culture simulated fish processing conditions and chemical disinfection challenges. Int. J. Food Microbiol. 2018, 267, 9–19. [Google Scholar] [CrossRef]
- Parkar, S.G.; Flint, S.H.; Palmer, J.S.; Brooks, J.D. Factors influencing attachment of thermophilic bacilli to stainless steel. J. Appl. Microbiol. 2001, 90, 901–908. [Google Scholar] [CrossRef]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hébraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Teh, K.H.; Flint, S.; Palmer, J.; Andrewes, P.; Bremer, P.; Lindsay, D. Biofilm− An unrecognised source of spoilage enzymes in dairy products? Int. Dairy J. 2014, 34, 32–40. [Google Scholar] [CrossRef]
- Aswathanarayan, J.B.; Vittal, R.R. Attachment and biofilm formation of Pseudomonas fluorescens PSD4 isolated from a dairy processing line. Food Sci. Biotechnol. 2014, 23, 1903–1910. [Google Scholar] [CrossRef]
- Puga, C.H.; Orgaz, B.; Muñoz, S.; SanJose, C. Cold stress and presence of Pseudomonas fluorescens affect Listeria monocytogenes biofilm structure and response to chitosan. Mol. Genet. Med. 2015, 9, 1–6. [Google Scholar]
- Hassan, A.N.; Birt, D.M.; Frank, J.F. Behavior of Listeria monocytogenes in a Pseudomonas putida biofilm on a condensate-forming surface. J. Food Prot. 2004, 67, 322–327. [Google Scholar] [CrossRef]
- Combrouse, T.; Sadovskaya, I.; Faille, C.; Kol, O.; Guérardel, Y.; Midelet-Bourdin, G. Quantification of the extracellular matrix of the Listeria monocytogenes biofilms of different phylogenic lineages with optimization of culture conditions. J. Appl. Microbiol. 2013, 114, 1120–1131. [Google Scholar] [CrossRef]
- Borucki, M.K.; Peppin, J.D.; White, D.; Loge, F.; Call, D.R. Variation in biofilm formation among strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2003, 69, 7336–7342. [Google Scholar] [CrossRef] [Green Version]
- Zameer, F.; Rukmangada, M.S.; Chauhan, J.B.; Khanum, S.A.; Kumar, P.; Devi, T.A.; Prasad, M.N.; Dhananjaya, B.L. Evaluation of adhesive and anti-adhesive properties of Pseudomonas aeruginosa biofilms and their inhibition by herbal plants. Iran. J. Microbiol. 2016, 8, 108–119. [Google Scholar]
- Simoes, M.; Pereira, M.O.; Vieira, M.J. Effect of mechanical stress on biofilms challenged by different chemicals. Water Res. 2005, 39, 5142–5152. [Google Scholar] [CrossRef] [Green Version]
- Petrova, O.E.; Sauer, K. Escaping the biofilm in more than one way: Desorption, detachment or dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Christopher, A.; Gulvik, C.A.; May, A.L.; Buchana, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine Roseobacter Phaeobacter sp. strain Y4IW. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.S.; Robins, K.J.; Ackerley, D.F. A sensitive single-enzyme assay system using the non-ribosomal peptide synthetase BpsA for measurement of L-glutamine in biological samples. Sci. Rep. 2017, 7, 41745. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Name | Source of Isolation | Serotype | Pulsotype ApaI | Pulsotype AscI |
|---|---|---|---|---|---|
| L. monocytogenes | LM1 | Gorgonzola cheese | 1/2b | GX6A12.0051 | GX6A16.0071 |
| L. monocytogenes | LM2 | Mozzarella cheese | 4b | GX6A12.0073 | GX6A16.0010 |
| L. monocytogenes | LM3 | Gorgonzola cheese | 1/2a | GX6A12.0032 | GX6A16.0029 |
| L. monocytogenes | LM4 | Caciotta cheese | 1/2a | GX6A12.0390 | GX6A16.0271 |
| L. monocytogenes | LM5 | Environmental | 1/2b | GX6A12.0349 | GX6A16.0255 |
| L. monocytogenes | LM6 | Environmental | 1/2b | GX6A12.0005 | GX6A16.0009 |
| L. monocytogenes | LM7 | Environmental | 1/2c | GX6A12.0373 | GX6A16.0261 |
| L. monocytogenes | LM8 | Environmental | 1/2c | GX6A12.0002 | GX6A16.0007 |
| P. fluorescens | pf5 | Mozzarella cheese |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggio, F.; Rossi, C.; Chaves-López, C.; Serio, A.; Valbonetti, L.; Pomilio, F.; Chiavaroli, A.P.; Paparella, A. Interactions between L. monocytogenes and P. fluorescens in Dual-Species Biofilms under Simulated Dairy Processing Conditions. Foods 2021, 10, 176. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010176
Maggio F, Rossi C, Chaves-López C, Serio A, Valbonetti L, Pomilio F, Chiavaroli AP, Paparella A. Interactions between L. monocytogenes and P. fluorescens in Dual-Species Biofilms under Simulated Dairy Processing Conditions. Foods. 2021; 10(1):176. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010176
Chicago/Turabian StyleMaggio, Francesca, Chiara Rossi, Clemencia Chaves-López, Annalisa Serio, Luca Valbonetti, Francesco Pomilio, Alessio Pio Chiavaroli, and Antonello Paparella. 2021. "Interactions between L. monocytogenes and P. fluorescens in Dual-Species Biofilms under Simulated Dairy Processing Conditions" Foods 10, no. 1: 176. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10010176