
Bee Pollen Diet Alters the Bacterial Flora and Antimicrobial Peptides in the Oral Cavities of Mice

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
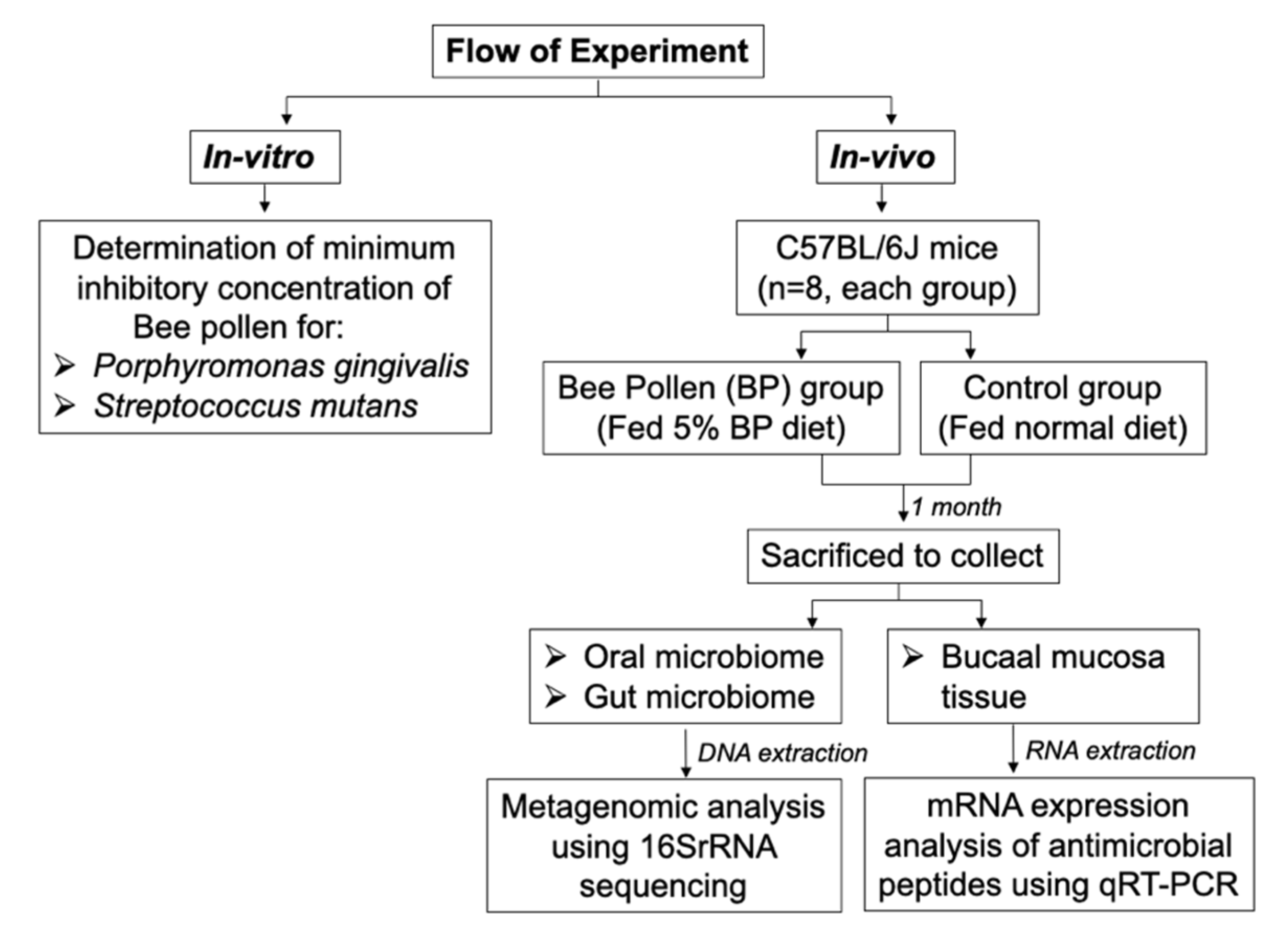
2. Materials and Methods
2.1. Animals
2.2. Bacteria Collection and DNA Extraction
2.3. Sequencing and Library Preparation
2.4. Analysis of Sequencing Data
2.5. Tissue Collection and RNA Extraction and Quantitative Reverse Transcriptase-PCR
3. Results
3.1. Minimum Inhibitory Concentration
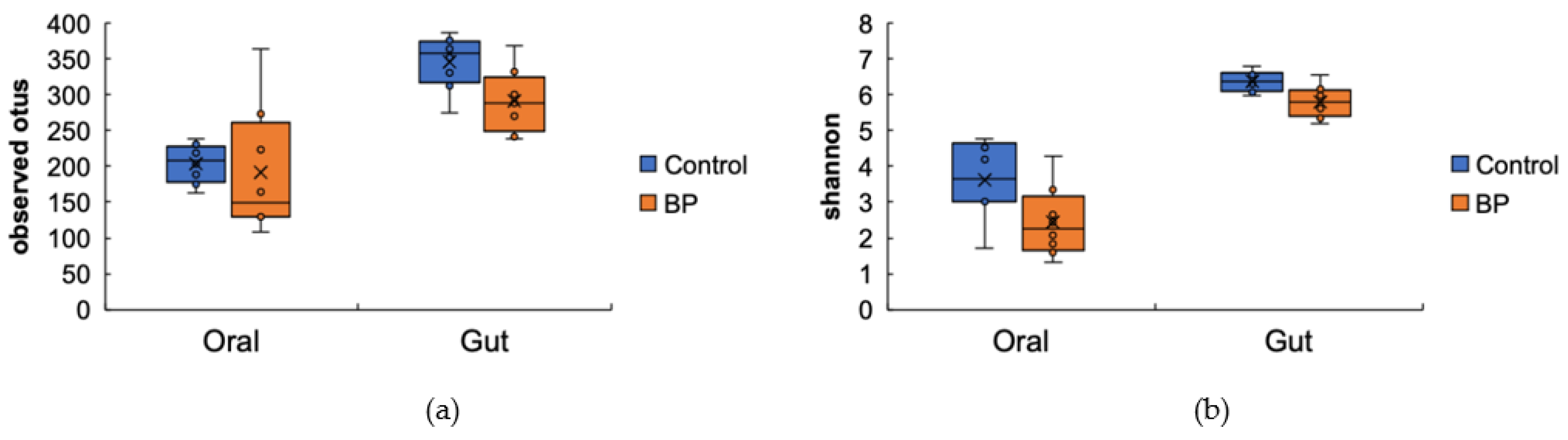
3.2. Oral and Gut Bacterial Species Richness and Diversity (Alpha Diversity)
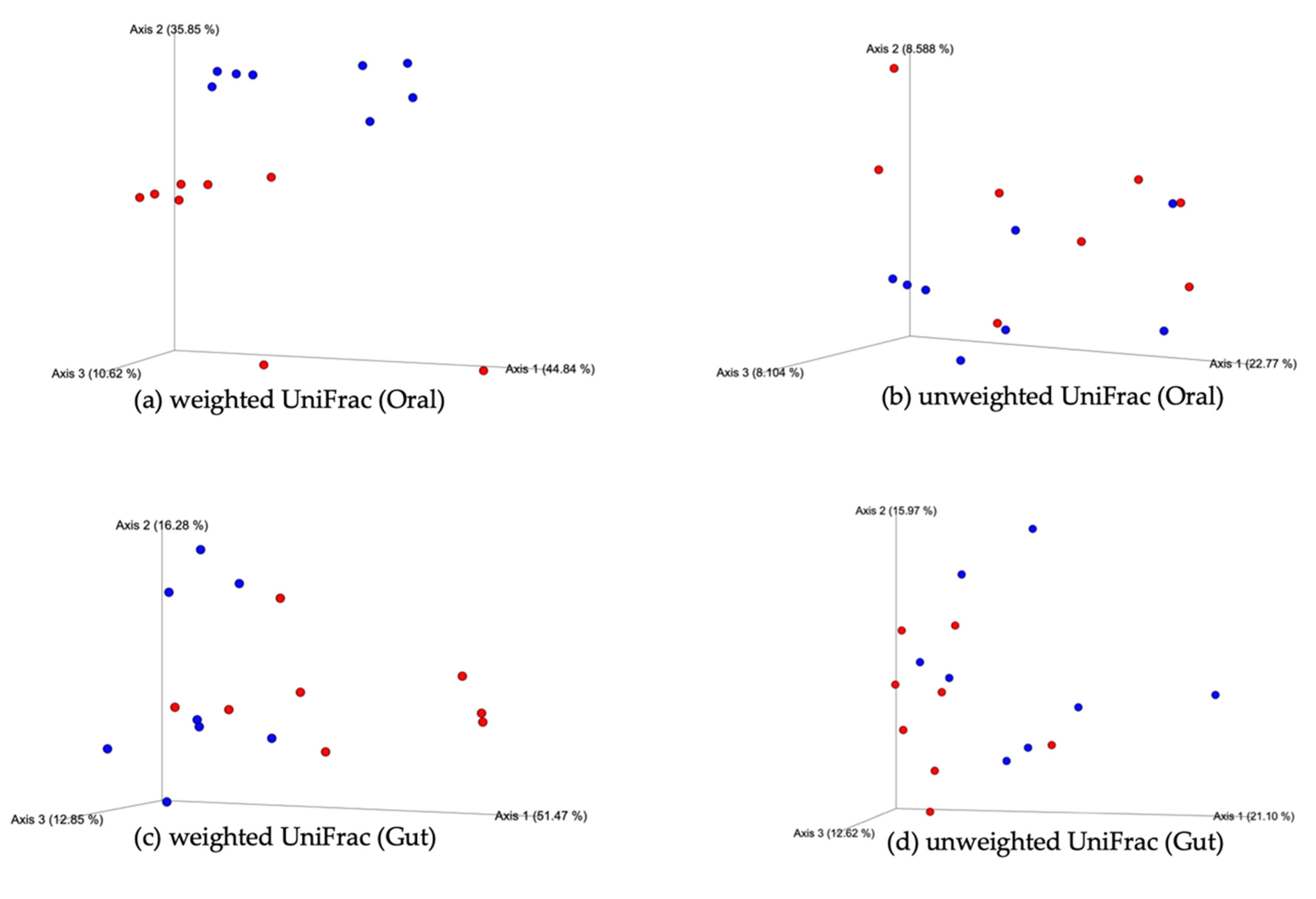
3.3. Oral and Gut Bacterial PCoA of the Weighted and Unweighted UniFrac (Beta Diversity)
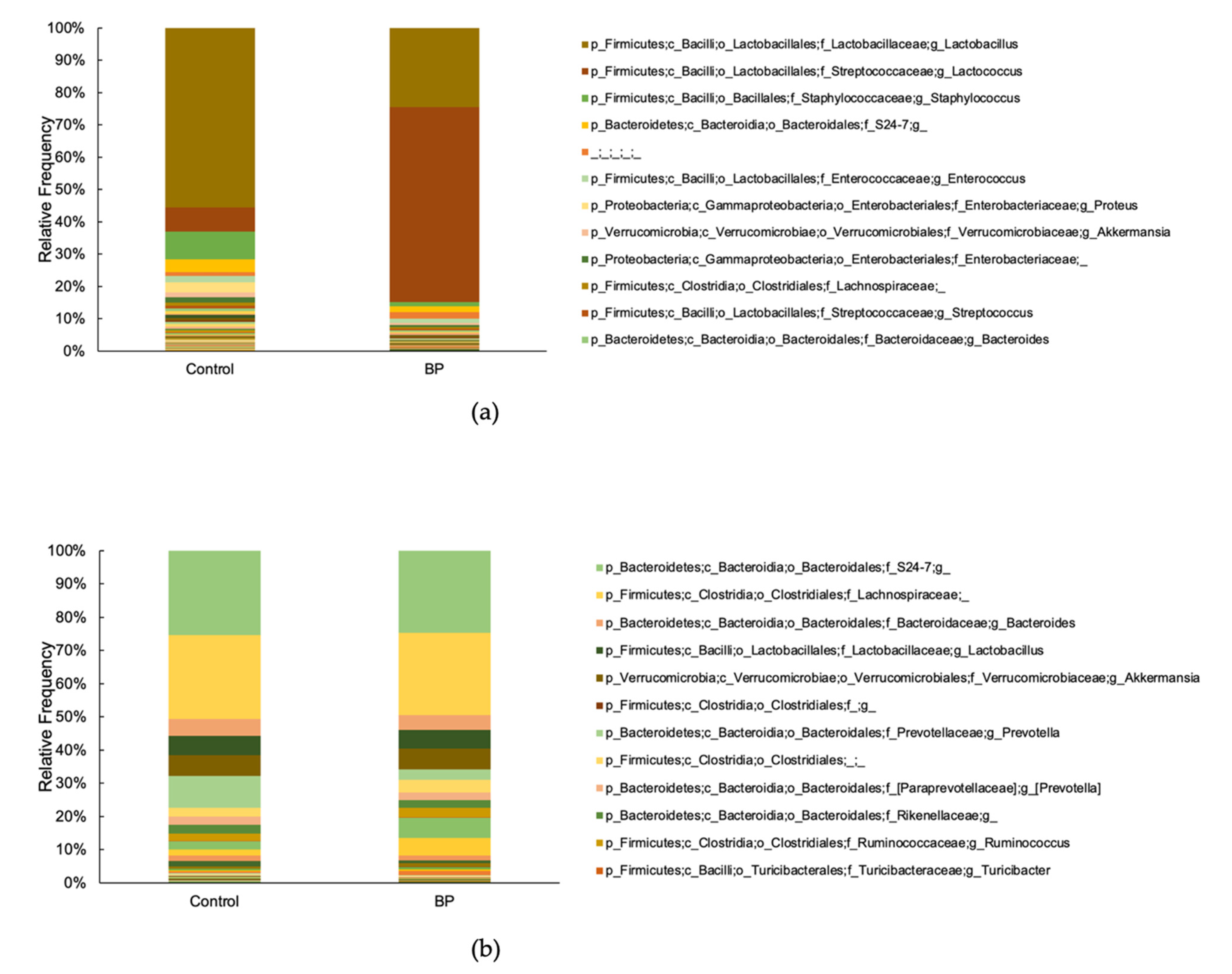
3.4. Oral and Gut Bacterial Taxonomy
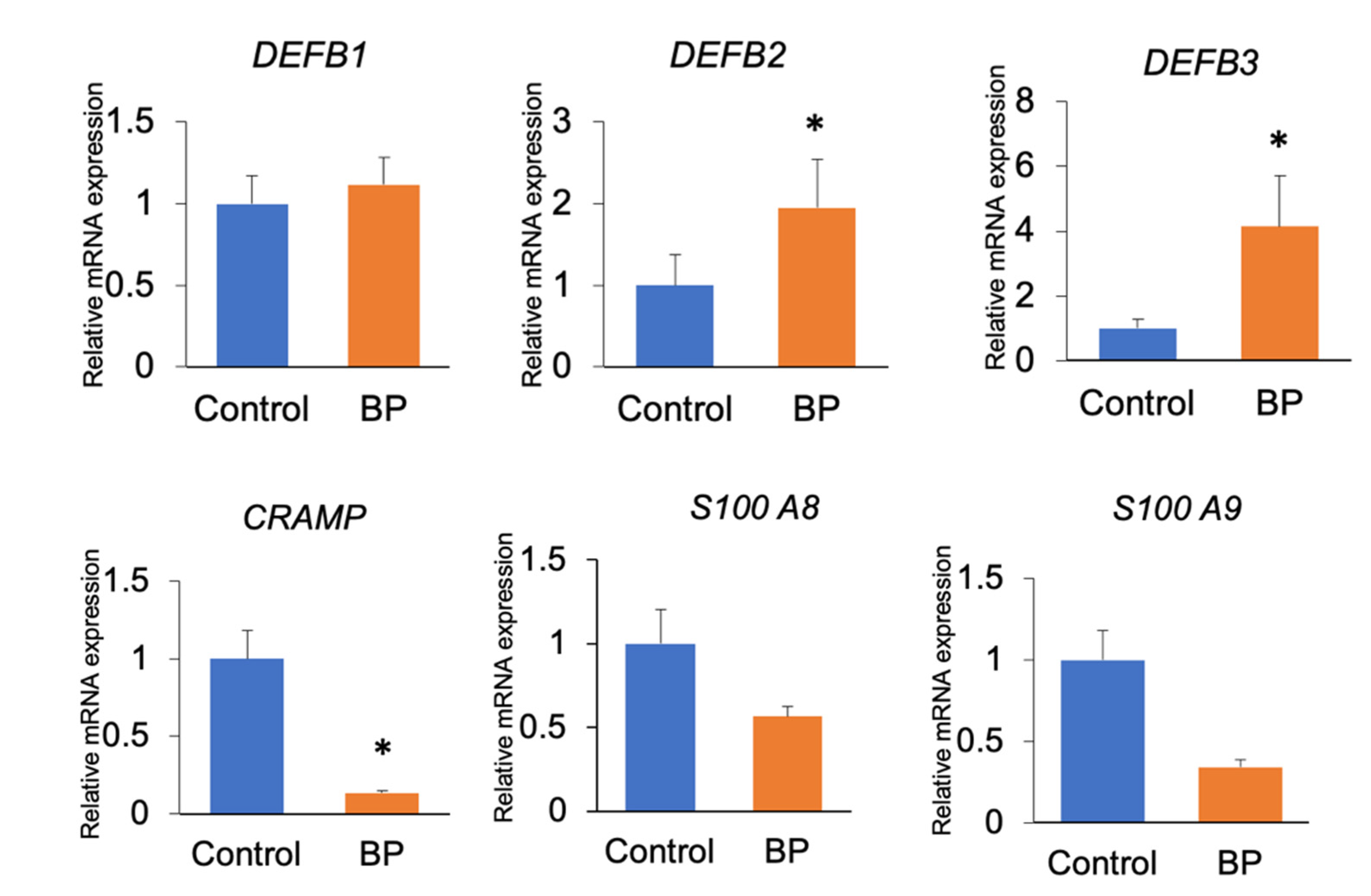
3.5. Gene Expression Levels in the Oral and Gut Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Martinello, M.; Mutinelli, F. Antioxidant Activity in Bee Products: A Review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Kaźmierczak, J.; Mencner, L.; Olczyk, K. Bee Pollen: Chemical Composition and Therapeutic Application. Evid.-Based Complement. Altern. Med. 2015, 2015, 297425. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Muradian, L.B.; Pamplona, L.C.; Coimbra, S.; Barth, O.M. Chemical composition and botanical evaluation of dried bee pollen pellets. J. Food Compos. Anal. 2005, 18, 105–111. [Google Scholar] [CrossRef]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 2018, 1–29. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Escapa, I.F.; Chen, T.; Huang, Y.; Gajare, P.; Dewhirst, F.E.; Lemon, K.P. New Insights into Human Nostril Microbiome from the Expanded Human Oral Microbiome Database (eHOMD): A Resource for the Microbiome of the Human Aerodigestive Tract. mSystems 2018, 3, e00187-18. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Olsen, I. Systemic Diseases Caused by Oral Infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Ling, K.H.; Wang, M.F.; El-Nezami, H. Green tea polyphenol epigallocatechin-3-gallate improves epithelial barrier function by inducing the production of antimicrobial peptide pBD-1 and pBD-2 in monolayers of porcine intestinal epithelial IPEC-J2 cells. Mol. Nutr. Food Res. 2016, 60, 1048–1058. [Google Scholar] [CrossRef]
- Lombardo Bedran, T.B.; Feghali, K.; Zhao, L.; Palomari Spolidorio, D.M.; Grenier, D. Green tea extract and its major constituent, epigallocatechin-3-gallate, induce epithelial beta-defensin secretion and prevent beta-defensin degradation by Porphyromonas gingivalis. J. Periodontal Res. 2014, 49, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Zhou, L.; Li, J.; Kuang, R.; Lin, Y.; Ni, L. An in vitro investigation of Lactococcus lactis antagonizing cariogenic bacterium Streptococcus mutans. Arch. Oral Biol. 2012, 57, 376–382. [Google Scholar] [CrossRef]
- Radaic, A.; Ye, C.; Parks, B.; Gao, L.; Kuraji, R.; Malone, E.; Kamarajan, P.; Zhan, L.; Kapila, Y.L. Modulation of pathogenic oral biofilms towards health with nisin probiotic. J. Oral Microbiol. 2020, 12, 1809302. [Google Scholar] [CrossRef] [PubMed]
- Grahn, E.; Holm, S.E.; Roos, K. Penicillin Tolerance in Beta-Streptococci Isolated from Patients with Tonsillitis. Scand. J. Infect. Dis. 1987, 19, 421–426. [Google Scholar] [CrossRef]
- Kimoto, H.; Nomura, M.; Kobayashi, M.; Mizumachi, K.; Okamoto, T. Survival of lactococci during passage through mouse digestive tract. Can. J. Microbiol. 2003, 49, 707–711. [Google Scholar] [CrossRef]
- Kimoto-Nira, H. New lactic acid bacteria for skin health via oral intake of heat-killed or live cells. Anim. Sci. J. 2018, 89, 835–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, F.; Murakami, Y.; Nishikawa, K.; Hasegawa, Y.; Kawaminami, S. Surface components of Porphyromonas gingivalis. J. Periodontal Res. 2009, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gursoy, U.K.; Könönen, E. Understanding the roles of gingival beta-defensins. J. Oral Microbiol. 2012, 4, 15127. [Google Scholar] [CrossRef] [PubMed]
- Ouhara, K.; Komatsuzawa, H.; Yamada, S.; Shiba, H.; Fujiwara, T.; Ohara, M.; Sayama, K.; Hashimoto, K.; Kurihara, H.; Sugai, M. Susceptibilities of periodontopathogenic and cariogenic bacteria to antibacterial peptides, β-defensins and LL37, produced by human epithelial cells. J. Antimicrob. Chemother. 2005, 55, 888–896. [Google Scholar] [CrossRef]
- Hans, M.; Madaan Hans, V. Epithelial Antimicrobial Peptides: Guardian of the Oral Cavity. Int. J. Pept. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Schröder, J.-M.; Harder, J. Human beta-defensin-2. Int. J. Biochem. Cell Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef]
- Tonetti, M.S.; Imboden, M.A.; Lang, N.P. Neutrophil Migration Into the Gingival Sulcus Is Associated With Transepithelial Gradients of Interleukin-8 and ICAM-1. J. Periodontol. 1998, 69, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.A.R.; Libério, S.A.; Guerra, R.N.M.; Ribeiro, M.N.S.; Nascimento, F.R.F. Mechanisms of action underlying the anti-inflammatory and immunomodulatory effects of propolis: A brief review. Rev. Bras. Farmacogn. 2012, 22, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi-Ishii, Y.; Nagaoka, I. NF-kappa B-mediated transcriptional regulation of human beta-defensin-2 gene following lipopolysaccharide stimulation. J. Leukoc. Biol. 2002, 71, 154–162. [Google Scholar] [PubMed]
- Hausmann, C.; Hertz-Kleptow, D.; Zoschke, C.; Wanjiku, B.; Wentzien-Odenthal, A.; Kerscher, M.; Schäfer-Korting, M. Reconstructed Human Epidermis Predicts Barrier-Improving Effects of Lactococcus lactis Emulsion in Humans. Skin Pharmacol. Physiol. 2019, 32, 72–80. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEFB1 | F: CCAGATGGAGCCAGGTGTTG |
| R: AGCTGGAGCGGAGACAGAATCC | |
| DEFB2 | F: AAGTATTGGATACGAAGCAG |
| R: TGGCAGAAGGAGGACAAATG | |
| DEFB3 | F: GCATTGGCAACACTCGTCAGA |
| R: CGGGATCTTGGTCTTCTCTA | |
| CRAMP | F: GGCGGTCACTATCACTGCTG |
| R: TCGGAACCTCACAGACTTGG | |
| S100A8 | F: TGCCCTCTACAAGAATGACT |
| R: AAGCTCTGCTACTCCTTGTG | |
| S100A9 | F: CGACACCTTCCATCAATACT |
| R: TCAGCATCATACACTCCTCA | |
| GAPDH | F: AGAACATCATCCCTGCATCC |
| R: CACATTGGGGGTAGGAACAC |
| Median Percentile Abundance | Max Percentile Abundance | ||||
|---|---|---|---|---|---|
| Control | BP | Control | BP | W | |
| Oral flora of genus p_Firmicutes;c_Bacilli;o_Lactobacillales;f_Streptococcaceae;g_Lactococcus | 2205.5 | 56,198 | 22,665 | 86,182 | 82 |
| Gut flora of genus p_Firmicutes;c_Bacilli;o_Turicibacterales;f_Turicibacteraceae;g_Turicibacter | 3671 | 582 | 8853 | 2999 | 18 |
| p_Firmicutes;c_Erysipelotrichi;o_Erysipelotrichales;f_Erysipelotrichaceae;g_Coprobacillus | 7.5 | 76 | 48 | 414 | 4 |
| p_Firmicutes;c_Bacilli;o_Lactobacillales;f_Streptococcaceae;g_Lactococcus | 1 | 3 | 1 | 10 | 4 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Lachnospiraceae;g_Clostridium | 12.5 | 1 | 34 | 5 | 3 |
| _;_;_;_;_ | 240 | 69.5 | 678 | 251 | 2 |
| p_Bacteroidetes;c_Flavobacteriia;o_Flavobacteriales;f_Flavobacteriaceae;g_Capnocytophaga | 1 | 1 | 1 | 3 | 2 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Veillonellaceae;g_Selenomonas | 1 | 1 | 1 | 6 | 2 |
| p_Proteobacteria;c_Gammaproteobacteria;o_Pseudomonadales;f_Moraxellaceae;g_Acinetobacter | 1 | 1 | 1 | 19 | 2 |
| p_TM7;c_TM7-3;o_;f_;g_ | 1 | 1 | 1 | 7 | 2 |
| p_Firmicutes;c_Erysipelotrichi;o_Erysipelotrichales;f_Erysipelotrichaceae;g_Clostridium | 1 | 1 | 1 | 3 | 1 |
| p_Proteobacteria;c_Gammaproteobacteria;o_Pasteurellales;f_Pasteurellaceae;g_Aggregatibacter | 1 | 1 | 3 | 5 | 1 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Ruminococcaceae;g_ | 743 | 1164.5 | 1644 | 2639 | 1 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Dehalobacteriaceae;g_Dehalobacterium | 177 | 100 | 328 | 449 | 1 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Christensenellaceae;g_ | 1 | 1 | 3 | 1 | 1 |
| p_Tenericutes;c_Mollicutes;o_RF39;f_;g_ | 32.5 | 64.5 | 163 | 129 | 1 |
| p_Firmicutes;c_Bacilli;o_Lactobacillales;f_Lactobacillaceae;g_Lactobacillus | 8426.5 | 2474 | 15,887 | 8004 | 1 |
| p_Deferribacteres;c_Deferribacteres;o_Deferribacterales;f_Deferribacteraceae;g_Mucispirillum | 120.5 | 23.5 | 2403 | 55 | 1 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Peptostreptococcaceae;g_Peptostreptococcus | 1 | 1 | 19 | 13 | 1 |
| p_Bacteroidetes;_;_;_;_ | 1 | 1 | 3 | 1 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;_;_ | 1 | 1 | 1 | 3 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Bacteroidaceae;g_Bacteroides | 7508.5 | 5739.5 | 9575 | 7975 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Rikenellaceae;_ | 79 | 59.5 | 119 | 139 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Rikenellaceae;g_ | 4200 | 1955 | 5724 | 5190 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_S24-7;g_ | 31,801.5 | 25,334.5 | 39,150 | 31,959 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Porphyromonadaceae;g_Parabacteroides | 921.5 | 645 | 1406 | 976 | 1 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_[Paraprevotellaceae];g_[Prevotella] | 3224 | 3911.5 | 3355 | 7706 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khurelchuluun, A.; Uehara, O.; Paudel, D.; Morikawa, T.; Kawano, Y.; Sakata, M.; Shibata, H.; Yoshida, K.; Sato, J.; Miura, H.; et al. Bee Pollen Diet Alters the Bacterial Flora and Antimicrobial Peptides in the Oral Cavities of Mice. Foods 2021, 10, 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061282
Khurelchuluun A, Uehara O, Paudel D, Morikawa T, Kawano Y, Sakata M, Shibata H, Yoshida K, Sato J, Miura H, et al. Bee Pollen Diet Alters the Bacterial Flora and Antimicrobial Peptides in the Oral Cavities of Mice. Foods. 2021; 10(6):1282. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061282
Chicago/Turabian StyleKhurelchuluun, Ariuntsetseg, Osamu Uehara, Durga Paudel, Tetsuro Morikawa, Yutaka Kawano, Mashu Sakata, Hiroshi Shibata, Koki Yoshida, Jun Sato, Hiroko Miura, and et al. 2021. "Bee Pollen Diet Alters the Bacterial Flora and Antimicrobial Peptides in the Oral Cavities of Mice" Foods 10, no. 6: 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061282