
Differences in Polyamine Content between Human Milk and Infant Formulas

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.1.1. Human Milk
2.1.2. Infant Formulas
2.2. Polyamine Analysis
2.3. Statistical Analysis
3. Results and Discussion
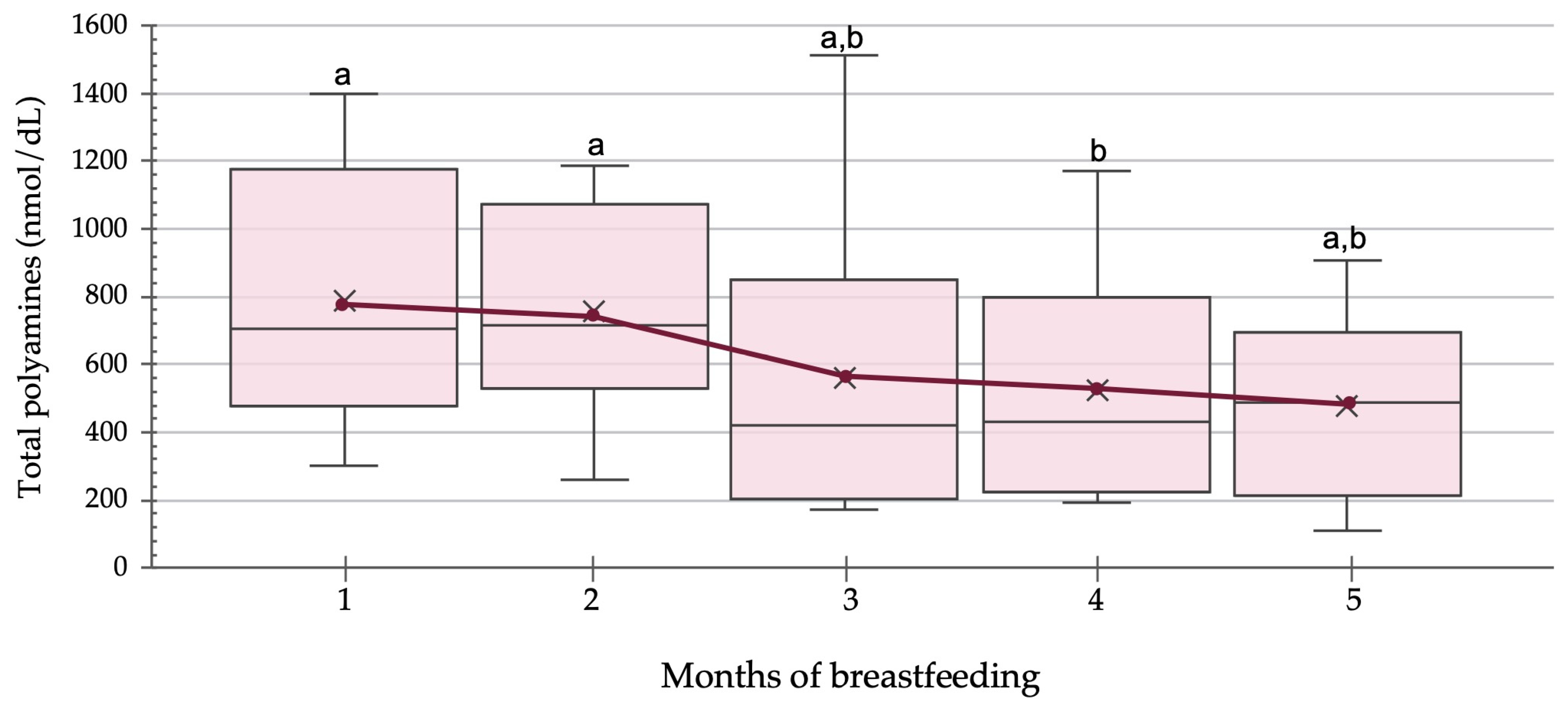
3.1. Polyamines in Human Milk
3.2. Polyamines in Infant Formulas
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mosca, F.; Giannì, M.L. Human milk: Composition and health benefits. Pediatr. Med. Chir. 2017, 39, 155. [Google Scholar] [CrossRef] [Green Version]
- Garwolińska, D.; Namieśnik, J.; Kot-Wasik, A.; Hewelt-Belka, W. Chemistry of Human Breast Milk—A Comprehensive Review of the Composition and Role of Milk Metabolites in Child Development. J. Agric. Food Chem. 2018, 66, 11881–11896. [Google Scholar] [CrossRef] [PubMed]
- WHO. Infant and young feeding. In Model Chapter for Textbooks for Medical Students and allied Health Professionals; WHO: Geneva, Switzerland, 2009. [Google Scholar]
- Gallego, C.G.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 2016, 21, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Andreas, N.J.; Kampmann, B.; Le-Doare, K.M. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gallego, C.; Kumar, H.; García-Mantrana, I.; Du Toit, E.; Suomela, J.-P.; Linderborg, K.M.; Zhang, Y.; Isolauri, E.; Yang, B.; Salminen, S.; et al. Breast Milk Polyamines and Microbiota Interactions: Impact of Mode of Delivery and Geographical Location. Ann. Nutr. Metab. 2017, 70, 184–190. [Google Scholar] [CrossRef]
- Romain, N.; Dandrifosse, G.; Jeusette, F.; Forget, P. Polyamine Concentration in Rat Milk and Food, Human Milk, and Infant Formulas1. Pediatr. Res. 1992, 32, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Pollack, P.F.; Koldovskỳ, O.; Nishioka, K. Polyamines in human and rat milk and in infant formulas. Am. J. Clin. Nutr. 1992, 56, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buts, J.-P.; De Keyser, N.; De Raedemaeker, L.; Collette, E.; Sokal, E. Polyamine Profiles in Human Milk, Infant Artificial Formulas, and Semi-elemental Diets. J. Pediatr. Gastroenterol. Nutr. 1995, 21, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Strandvik, B.; Palme-Kilander, C.; Yngve, A. Lower polyamine levels in breast milk of obese mothers compared to mothers with normal body weight. J. Hum. Nutr. Diet. 2013, 26, 164–170. [Google Scholar] [CrossRef]
- Zamora, J.P.; Sabater-Molina, M.; Rodríguez-Palmero, M.; Rivero, M.; Bosch, V.; Nadal, J.M.; Zamora, S.; Larque, E. Polyamines in human breast milk for preterm and term infants. Br. J. Nutr. 2013, 110, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Strandvik, B.; Sabel, K.-G.; Kilander, C.P.; Stromberg, R.; Yngve, A. Polyamine levels in breast milk are associated with mothers’ dietary intake and are higher in preterm than full-term human milk and formulas. J. Hum. Nutr. Diet. 2013, 27, 459–467. [Google Scholar] [CrossRef]
- Muñoz-Esparza, N.C.; Vásquez-Garibay, E.M.; Guzmán-Mercado, E.; Larrosa-Haro, A.; Comas-Basté, O.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Influence of Breastfeeding Factors on Polyamine Content in Human Milk. Nutrients 2021, 13, 3016. [Google Scholar] [CrossRef] [PubMed]
- Löser, C. Polyamines in human and animal milk. Br. J. Nutr. 2000, 84, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Dorhout, B.; Van Beusekom, C.M.; Huisman, M.; Kingma, A.W.; De Hoog, E.; Boersma, E.R.; Muskiet, F.A.J. Estimation of 24-h Polyamine Intake from Mature Human Milk. J. Pediatr. Gastroenterol. Nutr. 1996, 23, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.; Ladero, V.; Redruello, B.; Del Rio, B.; Fernández, L.; Rodríguez, J.; Martín, M.C.; Fernández, M.; Alvarez, M.A. Mastitis Modifies the Biogenic Amines Profile in Human Milk, with Significant Changes in the Presence of Histamine, Putrescine and Spermine. PLoS ONE 2016, 11, e0162426. [Google Scholar] [CrossRef]
- Koletzko, B.; Baker, S.; Cleghorn, G.; Neto, U.F.; Gopalan, S.; Hernell, O.; Hock, Q.S.; Jirapinyo, P.; Lonnerdal, B.; Pencharz, P.; et al. Global Standard for the Composition of Infant Formula: Recommendations of an ESPGHAN Coordinated International Expert Group. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 584–599. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Villares, J.M. Actualización en fórmulas infantiles. An. Pediatría Contin. 2011, 9, 31–40. [Google Scholar] [CrossRef]
- Lönnerdal, B. Infant formula and infant nutrition: Bioactive proteins of human milk and implications for composition of infant formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef] [Green Version]
- Tamayo López, G.; De Sáenz Urturi, A.; Hernández Sáez, M.R.; Pedrón Giner, C.; Garcia Novo, M.D. Formulas infantiles especiales. An. Esp. Pediatr. 1997, 47, 455–465. [Google Scholar]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the essential composition of infant and follow-on formulae. EFSA J. 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gallego, C.; Recio, I.; Gómez-Gómez, V.; Ortuño, I.; Bernal, M.; Ros, G.; Periago, M. Effect of processing on polyamine content and bioactive peptides released after in vitro gastrointestinal digestion of infant formulas. J. Dairy Sci. 2016, 99, 924–932. [Google Scholar] [CrossRef] [Green Version]
- Spizzirri, U.G.; Puoci, F.; Iemma, F.; Restuccia, D. Biogenic amines profile and concentration in commercial milks for infants and young children. Food Addit. Contam. Part A 2019, 36, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Esparza, N.C.; Latorre-Moratalla, M.L.; Comas-Basté, O.; Toro-Funes, N.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Polyamines in Food. Front. Nutr. 2019, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Biol-N’Garagba, M.-C.; Greco, S.; George, P.; Hugueny, I.; Louisot, P. Polyamine Participation in the Maturation of Glycoprotein Fucosylation, but not Sialylation, in Rat Small Intestine. Pediatr. Res. 2002, 51, 625–634. [Google Scholar] [CrossRef]
- Peulen, O.; Gharbi, M.; Powroznik, B.; Dandrifosse, G. Differential effect of dietary spermine on alkaline phosphatase activity in jejunum and ileum of unweaned rats. Biochimie 2004, 86, 487–493. [Google Scholar] [CrossRef]
- Perez-Cano, F.J.; González-Castro, A.; Castellote, C.; Franch, A.; Castell, M. Influence of breast milk polyamines on suckling rat immune system maturation. Dev. Comp. Immunol. 2010, 34, 210–218. [Google Scholar] [CrossRef]
- Dandrifosse, G.; Peulen, O.; El Khefif, N.; Deloyer, P.; Dandrifosse, A.C.; Grandfils, C. Are milk polyamines preventive agents against food allergy? Proc. Nutr. Soc. 2000, 59, 81–86. [Google Scholar] [CrossRef]
- Almeida, C.C.; Pereira, B.F.M.; Leandro, K.C.; Costa, M.P.; Spisso, B.F.; Conte-Junior, C.A. Bioactive Compounds in Infant Formula and Their Effects on Infant Nutrition and Health: A Systematic Literature Review. Int. J. Food Sci. 2021, 2021, 1–31. [Google Scholar] [CrossRef]
- Gómez-Gallego, C.; Frias, R.; Martinez, G.P.; Bernal, M.J.; Periago, M.J.; Salminen, S.; Ros, G.; Collado, M.C. Polyamine supplementation in infant formula: Influence on lymphocyte populations and immune system-related gene expression in a Balb/cOlaHsd mouse model. Food Res. Int. 2014, 59, 8–15. [Google Scholar] [CrossRef]
- Latorre-Moratalla, M.; Bosch-Fuste, J.; Lavizzari, T.; Bover-Cid, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Validation of an ultra high pressure liquid chromatographic method for the determination of biologically active amines in food. J. Chromatogr. A 2009, 1216, 7715–7720. [Google Scholar] [CrossRef] [PubMed]
- Bover-Cid, S.; Latorre-Moratalla, M.; Veciana-Nogués, M.; Vidal-Carou, M. Processing Contaminants: Biogenic Amines. Encycl. Food Saf. 2014, 381–391. [Google Scholar] [CrossRef]
- Kalač, P. Health effects and occurrence of dietary polyamines: A review for the period 2005–mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Esparza, N.; Costa-Catala, J.; Comas-Basté, O.; Toro-Funes, N.; Latorre-Moratalla, M.; Veciana-Nogués, M.; Vidal-Carou, M. Occurrence of Polyamines in Foods and the Influence of Cooking Processes. Foods 2021, 10, 1752. [Google Scholar] [CrossRef]
- Ruiz-Cano, D. Implicaciones de las poliaminas en la salud infantil. Arch. Argent. Pediatr. 2012, 110, 244–250. [Google Scholar] [CrossRef]
- Bardócz, S.; Duguid, T.J.; Brown, D.S.; Grant, G.; Pusztai, A.; White, A.; Ralph, A. The importance of dietary polyamines in cell regeneration and growth. Br. J. Nutr. 1995, 73, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, K.; Shiina, R.; Kashiwagi, K.; Igarashi, K. Decrease in Polyamines with Aging and Their Ingestion from Food and Drink. J. Biochem. 2006, 139, 81–90. [Google Scholar] [CrossRef]
- Glória, M.B.A.; Tavares-Neto, J.; Labanca, R.A.; Carvalho, M.S. Influence of Cultivar and Germination on Bioactive Amines in Soybeans (Glycine max L. Merril). J. Agric. Food Chem. 2005, 53, 7480–7485. [Google Scholar] [CrossRef]
- Ponce De León, C.; Torija, E.; Matallana, M.C. Utilidad en la alimentación de algunas semillas germinadas: Brotes de soja y trigo. Bol. R. Soc. Esp. Hist. Nat. Sec. Biol 2013, 107, 47–55. [Google Scholar]
- Menéndez, A.B.; Calzadilla, P.I.; Sansberro, P.A.; Espasandin, F.D.; Gázquez, A.; Bordenave, C.D.; Maiale, S.; Rodríguez, A.; Maguire, V.G.; Campestre, M.P.; et al. Polyamines and Legumes: Joint Stories of Stress, Nitrogen Fixation and Environment. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
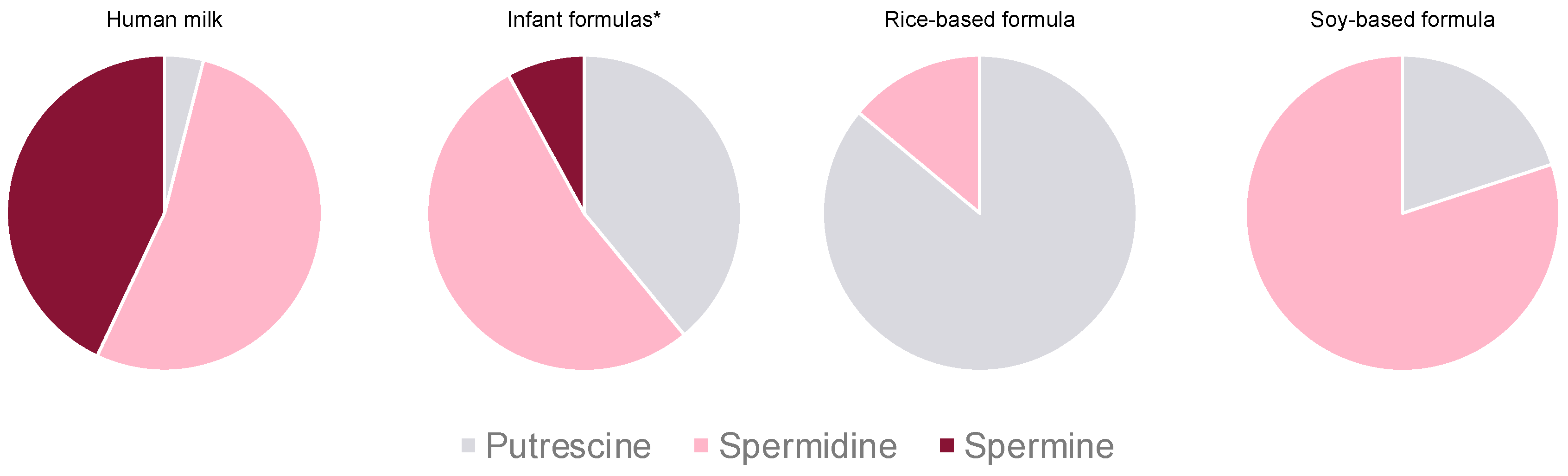{kind=link}
| Infant Formulas | Batch | Putrescine | Spermidine | Spermine |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | ||
| First formula | ||||
| A1 | 1 | Nd | 10.7 ± 0.5 | Nd |
| 2 | Nd | 20.7 ± 1.0 | Nd | |
| A2 | 1 | 40.3 ± 0.8 | 26.5 ± 0.5 | Nd |
| 2 | 33.5 ± 0.8 | 18.6 ± 2.0 | Nd | |
| A3 | 1 | 36.3 ± 1.6 | 45.1 ± 1.5 | Nd |
| 2 | 37.4 ± 1.6 | 42.3 ± 1.4 | Nd | |
| Follow-on formula | ||||
| B1 | 1 | 40.3 ± 0.8 | 32.7 ± 2.4 | Nd |
| 2 | 42.5 ± 0.8 | 39.2 ± 1.0 | Nd | |
| B2 | 1 | 35.7 ± 0.8 | 32.0 ± 1.5 | Nd |
| 2 | 34.6 ± 0.8 | 32.7 ± 0.5 | Nd | |
| B3 | 1 | 59.6 ± 0.8 # | 47.2 ± 4.8 | Nd |
| 2 | 57.3 ± 0.8 # | 56.8 ± 4.3 | Nd | |
| Preterm formula | ||||
| C1 | 1 | 47.1 ± 0.8 | 29.3 ± 0.5 | 62.0 ± 1.1 |
| 2 | 38.0 ± 0.8 | 24.8 ± 1.0 | 61.7 ± 2.8 | |
| C2 | 1 | Nd | 226.9 ± 3.4 # | 63.7 ± 7.7 |
| 2 | Nd | 229.6 ± 7.8 # | 61.5 ± 3.8 | |
| C3 | 1 | 39.1 ± 0.8 | 33.4 ± 0.5 | Nd |
| 2 | 39.1 ± 0.8 | 33.4 ± 0.5 | Nd | |
| Rice-based formula | ||||
| D1 | 1 | 188.9 ± 12.0 † | 18.6 ± 2.0 | Nd |
| 2 | 191.7 ± 6.4 † | 13.8 ± 2.9 | Nd | |
| D2 | 1 | 199.7 ± 1.6 † | 43.7 ± 3.4 | Nd |
| 2 | 188.3 ± 4.8 † | 52.7 ± 0.5 | Nd | |
| D3 | 1 | 306.3 ± 28.9 † | 53.7 ± 6.8 * | Nd |
| 2 | 311.9 ± 24.1 † | 19.6 ± 1.5 * | Nd | |
| Soy-based formula | ||||
| E1 | 1 | 70.3 ± 1.6 | 337.4 ± 2.9 *† | Nd |
| 2 | 77.7 ± 0.8 | 278.5 ± 0.5 *† | Nd | |
| E2 | 1 | 43.1 ± 1.6 | 179.0 ± 1.0 *† | Nd |
| 2 | 60.7 ± 4.0 | 225.8 ± 1.9 *† | Nd | |
| E3 | 1 | 74.9 ± 1.6 | 303.6 ± 2.9 *† | Nd |
| 2 | 70.3 ± 3.2 | 258.2 ± 4.9 *† | Nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Esparza, N.C.; Comas-Basté, O.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Differences in Polyamine Content between Human Milk and Infant Formulas. Foods 2021, 10, 2866. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112866
Muñoz-Esparza NC, Comas-Basté O, Latorre-Moratalla ML, Veciana-Nogués MT, Vidal-Carou MC. Differences in Polyamine Content between Human Milk and Infant Formulas. Foods. 2021; 10(11):2866. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112866
Chicago/Turabian StyleMuñoz-Esparza, Nelly C., Oriol Comas-Basté, M. Luz Latorre-Moratalla, M. Teresa Veciana-Nogués, and M. Carmen Vidal-Carou. 2021. "Differences in Polyamine Content between Human Milk and Infant Formulas" Foods 10, no. 11: 2866. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112866