
Lactic Acid Bacteria as Antimicrobial Agents: Food Safety and Microbial Food Spoilage Prevention

 ,
,  ,
,  ,
,  , and
, and
Abstract
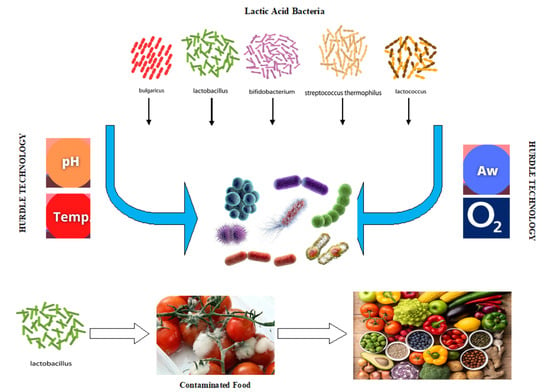
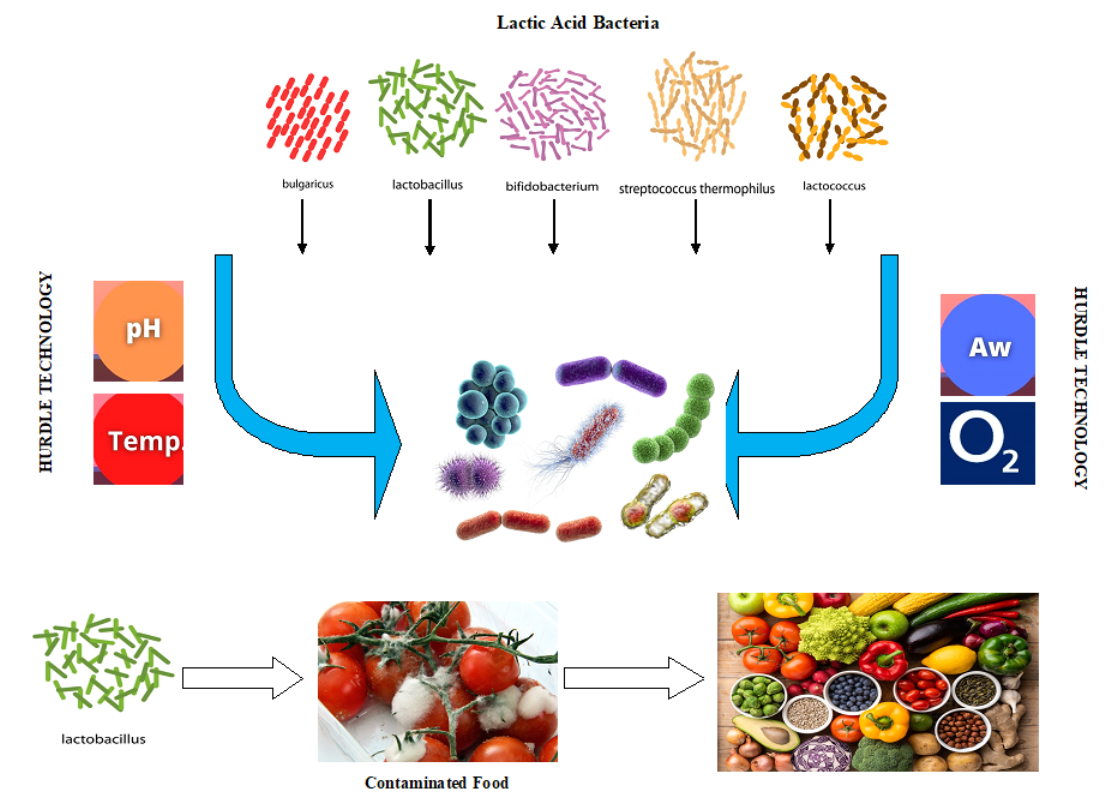
:
1. Introduction
2. The Microbial Ecosystem
3. LAB Affect the Growth of Microorganisms

{kind=link}
| Small Molecules | Example of Prominent LAB Producer | Example of Food Pathogen Application | Application in Food |
|---|---|---|---|
| Hydrogen Peroxide | Lactobacillus johnsonii [37] | Escherichia coli O157:H7 [38] Salmonella enterica [38] Listeria monocytogenes [38] | Lettuce [38] |
| Reuterin | Limosilactobacillus reuteri [40] | Campylobacter spp. [40] Escherichia coli O157:H7 [41] | Meat [40] |
| Diacetyl | Streptococcus diacetyl lactis [42] | Escherichia coli O157:H7 [43] Salmonella typhimurium [43] | Meat [43] |
4. Prevention of Foodborne Pathogens and Elimination of Food Spoilage Bacteria
5. Bacteriocin-Antimicrobial Synergy
6. Competitive Growth Interactions between LAB and Other Microorganisms
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiernasz, N.; Cornet, J.; Cardinal, M.; Pilet, M.-F.; Passerini, D.; Leroi, F. Lactic Acid Bacteria Selection for Biopreservation as a Part of Hurdle Technology Approach Applied on Seafood. Front. Mar. Sci. 2017, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, T.; Siddiqui, S.; Bischoff, W.; Ibrahim, S. Tackling Airborne Virus Threats in the Food Industry: A Proactive Approach. Int. J. Environ. Res. Public Health 2021, 18, 4335. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Wang, J.; Liu, M.; Li, P.; Gu, Q. Purification and characterization of a novel bacteriocin from Lactobacillus paracasei ZFM54. LWT 2021, 143, 111125. [Google Scholar] [CrossRef]
- Prosser, J.I.; Bohannan, B.J.M.; Curtis, T.P.; Ellis, R.J.; Firestone, M.K.; Freckleton, R.P.; Green, J.L.; Green, L.E.; Killham, K.; Lennon, J.J.; et al. The role of ecological theory in microbial ecology. Nat. Rev. Genet. 2007, 5, 384–392. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awaisheh, S.; Ibrahim, S.A. Screening of Antibacterial Activity of Lactic Acid Bacteria Against Different Pathogens Found in Vacuum-Packaged Meat Products. Foodborne Pathog. Dis. 2009, 6, 1125–1132. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually stimulating interactions between lactic acid bacteria and Saccharomyces cerevisiae in sourdough fermentation. LWT 2018, 90, 201–206. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2017, 49, 23–28. [Google Scholar] [CrossRef]
- Awaisheh, S.S.; Al-Nabulsi, A.A.; Osaili, T.M.; Ibrahim, S.; Holley, R. Inhibition of Cronobacter sakazakii by Heat Labile Bacteriocins Produced by Probiotic LAB Isolated from Healthy Infants. J. Food Sci. 2013, 78, M1416–M1420. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drug Administration. Fed Register Nisin Preparation: Affirmation of GRAS Status as a Direct Human Food In-Gredient; US Food and Drug Administration: Silver Spring, MD, USA, 1988; pp. 11247–11251.
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Factories 2014, 13, S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negash, A.W.; Tsehai, B.A. Current Applications of Bacteriocin. Int. J. Microbiol. 2020, 2020, 4374891. [Google Scholar] [CrossRef] [PubMed]
- Oscáriz, J.C.; Pisabarro, A.G. Classification and mode of action of membrane-active bacteriocins produced by gram-positive bacteria. Int. Microbiol. 2001, 4, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Dang, S.; Gupta, S.; Gabrani, R. Antibacterial activity, cytotoxicity, and the mechanism of action of bacteriocin from Bacillus subtilis GAS101. Med. Princ. Pract. 2018, 27, 186–192. [Google Scholar] [CrossRef]
- Ng, Z.J.; Abu Zarin, M.; Lee, C.K.; Tan, J.S. Application of bacteriocins in food preservation and infectious disease treatment for humans and livestock: A review. RSC Adv. 2020, 10, 38937–38964. [Google Scholar] [CrossRef]
- Mitra, S.; Mukhopadhyay, B.; Biswas, S.R. Potential application of the nisin Z preparation of Lactococcus lactis W8 in preservation of milk. Lett. Appl. Microbiol. 2011, 53, 98–105. [Google Scholar] [CrossRef]
- Budu-Amoako, E.; Ablett, R.F.; Harris, J.; Delves-Broughton, J. Combined effect of nisin and moderate heat on destruction of Listeria monocytogenes in cold-pack lobster meat. J. Food Prot. 1999, 62, 46–50. [Google Scholar] [CrossRef]
- Nykänen, A.; Weckman, K.; Lapveteläinen, A. Synergistic inhibition of Listeria monocytogenes on cold-smoked rainbow trout by nisin and sodium lactate. Int. J. Food Microbiol. 2000, 61, 63–72. [Google Scholar] [CrossRef]
- Ukuku, D.O.; Zhang, H.; Huang, L. Growth parameters of Escherichia coli O157:H7, Salmonella spp., Listeria monocytogenes, and aerobic mesophilic bacteria of apple cider amended with Nisin–EDTA. Foodborne Pathog. Dis. 2009, 6, 487–494. [Google Scholar] [CrossRef]
- von Staszewski, M.; Jagus, R.J. Natural antimicrobials: Effect of Microgard™ and nisin against Listeria innocua in liquid cheese whey. Int. Dairy J. 2008, 18, 255–259. [Google Scholar] [CrossRef]
- Morgan, S.; Galvin, M.; Ross, R.; Hill, C. Evaluation of a spray-dried lacticin 3147 powder for the control of Listeria monocytogenes and Bacillus cereus in a range of food systems. Lett. Appl. Microbiol. 2001, 33, 387–391. [Google Scholar] [CrossRef]
- Scannell, A.G.M.; Ross, R.; Hill, C.; Arendt, E.K. An effective lacticin biopreservative in fresh pork sausage. J. Food Prot. 2000, 63, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Arqués, J.L.; Rodríguez, E.; Nuñez, M.; Medina, M. Combined effect of reuterin and lactic acid bacteria bacteriocins on the inactivation of food-borne pathogens in milk. Food Control 2011, 22, 457–461. [Google Scholar] [CrossRef]
- Arakawa, K.; Kawai, Y.; Iioka, H.; Tanioka, M.; Nishimura, J.; Kitazawa, H.; Tsurumi, K.; Saito, T. Effects of gassericins A and T, bacteriocins produced by Lactobacillus gasseri, with glycine on custard cream preservation. J. Dairy Sci. 2009, 92, 2365–2372. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, Z.; Öncül, N.; Karabiyikli, Ş.; Yildirim, M. Application of lactococcin BZ and enterocin KP against Listeria monocytogenes in milk as biopreservation agents. Acta Aliment. 2016, 45, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Molinos, A.C.; Abriouel, H.; López, R.L.; Ben Omar, N.; Valdivia, E.; Gálvez, A. Enhanced bactericidal activity of enterocin AS-48 in combination with essential oils, natural bioactive compounds and chemical preservatives against Listeria monocytogenes in ready-to-eat salad. Food Chem. Toxicol. 2009, 47, 2216–2223. [Google Scholar] [CrossRef]
- Ayivi, R.; Gyawali, R.; Krastanov, A.; Aljaloud, S.; Worku, M.; Tahergorabi, R.; Da Silva, R.; Ibrahim, S. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 15. [Google Scholar] [CrossRef]
- Sallam, K.I. Antimicrobial and antioxidant effects of sodium acetate, sodium lactate, and sodium citrate in refrigerated sliced salmon. Food Control 2007, 18, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özcelik, S.; Kuley, E.; Özogul, F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. LWT 2016, 73, 536–542. [Google Scholar] [CrossRef]
- Ricke, S.C.; Dittoe, D.K.; Richardson, K.E. Formic Acid as an Antimicrobial for Poultry Production: A Review. Front. Vet. Sci. 2020, 7, 563. [Google Scholar] [CrossRef]
- Gómez-García, M.; Sol, C.; De Nova, P.J.G.; Puyalto, M.; Mesas, L.; Puente, H.; Mencía-Ares, Ó.; Miranda, R.; Argüello, H.; Rubio, P.; et al. Antimicrobial activity of a selection of organic acids, their salts and essential oils against swine enteropathogenic bacteria. Porc. Health Manag. 2019, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Radkowski, M.; Zdrodowska, B.; Gomółka-Pawlicka, M. Effect of Succinic Acid on Elimination of Salmonella in Chicken Meat. J. Food Prot. 2018, 81, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Ghoul, M.; Mitri, S. The Ecology and Evolution of Microbial Competition. Trends Microbiol. 2016, 24, 833–845. [Google Scholar] [CrossRef]
- Peh, E.; Kittler, S.; Reich, F.; Kehrenberg, C. Antimicrobial activity of organic acids against Campylobacter spp. and development of combinations—A synergistic effect? PLoS ONE 2020, 15, e0239312. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.N.; Chowdhury, R.; Islam, K.; Akbar, M.A. Propionic Acid Is An Alternative To Antibiotics In Poultry Diet. Bangladesh J. Anim. Sci. 1970, 38, 115–122. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Russell, J.B.; Flythe, M.; Gantois, I.; Timbermont, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. The use of organic acids to combatSalmonellain poultry: A mechanistic explanation of the efficacy. Avian Pathol. 2006, 35, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Hertzberger, R.; Arents, J.C.; Dekker, H.L.; Pridmore, R.D.; Gysler, C.; Kleerebezem, M.; De Mattos, M.J.T. H2O2Production in Species of the Lactobacillus acidophilus Group: A Central Role for a Novel NADH-Dependent Flavin Reductase. Appl. Environ. Microbiol. 2014, 80, 2229–2239. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-M.; Moon, S.S.; Doyle, M.P.; McWatters, K.H. Inactivation of Escherichia coli O157:H7, Salmonella enterica Serotype Enteritidis, and Listeria monocytogenes on Lettuce by Hydrogen Peroxide and Lactic Acid and by Hydrogen Peroxide with Mild Heat. J. Food Prot. 2002, 65, 1215–1220. [Google Scholar] [CrossRef]
- Nakajima, H.; Toba, T.; Ito, A.; Kudo, S.; Sato, S.; Sato, Y. The Screening of Hydrogen Peroxide-Producing Lactic Acid Bacteria and Their Application to Inactivating Psychrotrophic Food-Borne Pathogens. Curr. Microbiol. 2003, 47, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Asare, P.T.; Zurfluh, K.; Greppi, A.; Lynch, D.; Schwab, C.; Stephan, R.; Lacroix, C. Reuterin Demonstrates Potent Antimicrobial Activity Against a Broad Panel of Human and Poultry Meat Campylobacter spp. Isolates. Microorganisms 2020, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Bertin, Y.; Habouzit, C.; Dunière, L.; Laurier, M.; Durand, A.; Duchez, D.; Segura, A.; Thévenot-Sergentet, D.; Baruzzi, F.; Chaucheyras-Durand, F.; et al. Lactobacillus reuteri suppresses E. coli O157:H7 in bovine ruminal fluid: Toward a pre-slaughter strategy to improve food safety? PLoS ONE 2017, 12, e0187229. [Google Scholar] [CrossRef] [Green Version]
- Oberman, H.H.; Piątkiewicz, A.; Libudzisz, Z. Production of diacetyl and acetoin by lactic acid bacteria. Food/Nahrung 1982, 26, 615–623. [Google Scholar] [CrossRef]
- Kang, D.-H.; Fung, D.Y.C. Effect of Diacetyl on Controlling Escherichia coli O157:H7 and Salmonella Typhimurium in the Presence of Starter Culture in a Laboratory Medium and during Meat Fermentation. J. Food Prot. 1999, 62, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Hathout, A.S.; Aly, S.E. Role of lactic acid bacteria as a biopreservative agent of Talbina. J. Am. Sci. 2010, 6, 889–898. [Google Scholar]
- Heredia-Castro, P.Y.; Méndez-Romero, J.I.; Hernandez-Mendoza, A.; Acedo-Félix, E.; González-Córdova, A.F.; Vallejo-Cordoba, B. Antimicrobial activity and partial characterization of bacteriocin-like inhibitory substances produced by Lactobacillus spp. isolated from artisanal Mexican cheese. J. Dairy Sci. 2015, 98, 8285–8293. [Google Scholar] [CrossRef] [Green Version]
- Fhoula, I.; Najjari, A.; Turki, Y.; Jaballah, S.; Boudabous, A.; Ouzari, H. Diversity and Antimicrobial Properties of Lactic Acid Bacteria Isolated from Rhizosphere of Olive Trees and Desert Truffles of Tunisia. BioMed Res. Int. 2013, 2013, 405708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, C.; Almendárez, B.E.G.; Martin, S.E.; Regalado, C. Food-associated lactic acid bacteria with antimicrobial potential from traditional Mexican foods. Rev. Latinoam. Microbiol. 2008, 48, 260–268. [Google Scholar]
- Hernandez, D.; Cardell, E.; Zarate, V. Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese: Initial characterization of plantaricin TF711, a bacteriocin-like substance produced byLactobacillus plantarumTF711. J. Appl. Microbiol. 2005, 99, 77–84. [Google Scholar] [CrossRef]
- Bian, X.; Evivie, S.E.; Muhammad, Z.; Luo, G.-W.; Liang, H.-Z.; Wang, N.-N.; Huo, G.-C. In vitro assessment of the antimicrobial potentials of Lactobacillus helveticus strains isolated from traditional cheese in Sinkiang China against food-borne pathogens. Food Funct. 2015, 7, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Langa, S.; Martín-Cabrejas, I.; Montiel, R.; Landete, J.; Medina, M.; Arqués, J. Short communication: Combined antimicrobial activity of reuterin and diacetyl against foodborne pathogens. J. Dairy Sci. 2014, 97, 6116–6121. [Google Scholar] [CrossRef] [Green Version]
- Rahmeh, R.; Akbar, A.; Kishk, M.; Al-Onaizi, T.; Al-Azmi, A.; Al-Shatti, A.; Shajan, A.; Al-Mutairi, S.; Akbar, B. Distribution and antimicrobial activity of lactic acid bacteria from raw camel milk. New Microbes New Infect. 2019, 30, 100560. [Google Scholar] [CrossRef]
- Coman, M.M.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Boyko, N.; Cresci, A. In vitro evaluation of antimicrobial activity of Lactobacillus rhamnosus IMC 501®, Lactobacillus paracasei IMC 502 and SYNBIO against pathogens. J. Appl. Microbiol. 2014, 117, 518–527. [Google Scholar] [CrossRef] [PubMed]
- El-Ziney, M.G.; Debevere, J.M. The Effect of Reuterin on Listeria monocytogenes and Escherichia coli O157:H7 in Milk and Cottage Cheese. J. Food Prot. 1998, 61, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Nayab, H.; Rehman, T.U.; Williamson, M.P.; Haq, K.U.; Shafi, N.; Shafique, F. Characterisation of Bacteriocins Produced by Lactobacillus spp. Isolated from the Traditional Pakistani Yoghurt and Their Antimicrobial Activity against Common Foodborne Pathogens. BioMed Res. Int. 2020, 2020, 8281623. [Google Scholar] [CrossRef] [PubMed]
- Cizeikiene, D.; Juodeikiene, G.; Paskevicius, A.; Bartkiene, E. Antimicrobial activity of lactic acid bacteria against pathogenic and spoilage microorganism isolated from food and their control in wheat bread. Food Control 2013, 31, 539–545. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic Acid Bacteria as Antibacterial Agents to Extend the Shelf Life of Fresh and Minimally Processed Fruits and Vegetables: Quality and Safety Aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef]
- Ross, A.I.; Griffiths, M.W.; Mittal, G.S.; Deeth, H.C. Combining nonthermal technologies to control foodborne microorganisms. Int. J. Food Microbiol. 2003, 89, 125–138. [Google Scholar] [CrossRef]
- Ušković, J.; Kos, B.; Beganović, J.; Leboš Pavunc, A.; Habjanič, K.; Matošić, S. Antimicrobial activity–the most important property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, R. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Fang, T.J.; Tsai, H.C. Growth patterns of Escherichia coli O157: H7 in ground beef treated with nisin, chelators, organic acids and their combinations immobilized in calcium alginate gels. Food Microbiol. 2003, 20, 243–253. [Google Scholar] [CrossRef]
- Cotter, P.; Hill, C.; Ross, R. Bacterial Lantibiotics: Strategies to Improve Therapeutic Potential. Curr. Protein Pept. Sci. 2005, 6, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.; Hill, C.; Ross, R.P. Bacteriocin-Antimicrobial Synergy: A Medical and Food Perspective. Front. Microbiol. 2017, 8, 1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivarooban, T.; Hettiarachchy, N.S.; Johnson, M.G. Transmission Electron Microscopy Study of Listeria monocytogenes Treated with Nisin in Combination with either Grape Seed or Green Tea Extract. J. Food Prot. 2008, 71, 2105–2109. [Google Scholar] [CrossRef] [PubMed]
- Norhana, M.W.; Poole, S.E.; Deeth, H.C.; Dykes, G.A. Effects of nisin, EDTA and salts of organic acids on Listeria monocytogenes, Salmonella and native microflora on fresh vacuum packaged shrimps stored at 4 C. Food Microbiol. 2012, 31, 43–50. [Google Scholar] [CrossRef] [PubMed]
- McManamon, O.; Kaupper, T.; Scollard, J.; Schmalenberger, A. Nisin application delays growth of Listeria monocytogenes on fresh-cut iceberg lettuce in modified atmosphere packaging, while the bacterial community structure changes within one week of storage. Postharvest Biol. Technol. 2018, 147, 185–195. [Google Scholar] [CrossRef]
- Branen, J.K.; Davidson, P. Enhancement of nisin, lysozyme, and monolaurin antimicrobial activities by ethylenediaminetetraacetic acid and lactoferrin. Int. J. Food Microbiol. 2003, 90, 63–74. [Google Scholar] [CrossRef]
- Moon, G.S.; Kim, W.J.; Kim, M.H. Synergistic effects of bacteriocin-producing Pediococcus acidilactici K10 and organic acids on inhibiting Escherichia coli O157: H7 and applications in ground beef. J. Microbiol. Biotech. 2002, 12, 936–942. [Google Scholar]
- Mansour, M.; Milliere, J.B. An inhibitory synergistic effect of a nisin-monolaurin combination on Bacillus spp. vegetative cells in milk. Food Microbiol. 2001, 18, 87–94. [Google Scholar] [CrossRef]
- Abo-Amer, A.E. Characterization of a bacteriocin-like inhibitory substance produced by Lactobacillus plantarum isolated from Egyptian home-made yogurt. Sci. Asia 2007, 33, 313–319. [Google Scholar] [CrossRef]
- Benech, R.-O.; Kheadr, E.E.; Laridi, R.; Lacroix, C.; Fliss, I. Inhibition of Listeria innocua in Cheddar Cheese by Addition of Nisin Z in Liposomes or by In Situ Production in Mixed Culture. Appl. Environ. Microbiol. 2002, 68, 3683–3690. [Google Scholar] [CrossRef] [Green Version]
- Bouksaim, M.; Lacroix, C.; Audet, P.; Simard, R.E. Effects of mixed starter composition on nisin Z production by Lactococcallactis subsp. lactis biovar. diacetylactis UL 719 during production and ripening of gouda cheese. Int. J. Food Microbiol. 2000, 59, 141–156. [Google Scholar] [CrossRef]
- Rilla, N.; Martinez, B.; Deldago, T.; Rodriguez, A. Inhibition of Clostridium tyrobutyricum in Vidiago cheese by Lactococcuslactis ssp. lactis IPLA 729, a nisin Z producer. Int. J. Food Microbiol. 2003, 85, 23–33. [Google Scholar] [CrossRef]
- Giraffa, G.; Neviani, E.; Torri Tarelli, G. Antilisterial activity of enterococci in a model predicting the temperature evolution of Taleggio, an Italian soft cheese. J. Dairy Sci. 1994, 77, 1176–1182. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Leroy, F.; Leontopoulou, E.; Georgalaki, M.D.; Kalantzopoulos, G.; Tsakalidou, E.; De Vuyst, L. Bacteriocin production by Enterococcus faecium FAIR-E 198 in view of its application as adjunct starter in Greek Feta cheese making. Int. J. Food Microbiol. 2002, 72, 125–136. [Google Scholar] [CrossRef]
- Maisnier-Patin, S.; Deschamps, N.; Tatini, S.R.; Richard, J. Inhibition of Listeria monocytogenes in Camembert cheese made with a nisin-producing starter. Le Lait 1992, 72, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.J.; Hussein, H.M.; Zagorec, M.; Brightwell, G.; Tagg, J.R. Isolation of lactic acid bacteria with inhibitory activity against pathogens and spoilage organisms associated with fresh meat. Food Microbiol. 2008, 25, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Sidira, M.; Karapetsas, A.; Galanis, A.; Kanellaki, M.; Kourkoutas, Y. Effective survival of immobilized Lactobacillus casei during ripening and heat treatment of probiotic dry-fermented sausages and investigation of the microbial dynamics. Meat Sci. 2014, 96, 948–955. [Google Scholar] [CrossRef]
- Joshi, V.K.; Sharma, S.; Neerja, R. Production, purification, stability and efficiency of bacteriocin from the isolate of natural lactic acid fermentation of vegetables. Food Technol. Biotechnol. 2006, 44, 435–439. [Google Scholar]
- Szutowska, J.; Gwiazdowska, D. Probiotic potential of lactic acid bacteria obtained from fermented curly kale juice. Arch. Microbiol. 2020, 203, 975–988. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, S.M.; Agun, T.F.; Ogunlusi, E.D. Antimicrobial Activity of Lactic Acid Bacteria Isolated from ‘Pupuru’: An African Fermented Staple against Food Borne- Pathogens. J. Mol. Biol. Biotech. 2018, 3, 1. [Google Scholar]
- Stupar, J.; Holøymoen, I.G.; Hoel, S.; Lerfall, J.; Rustad, T.; Jakobsen, A.N. Diversity and Antimicrobial Activity to-wards Listeria spp. and Escherichia coli among Lactic Acid Bacteria Isolated from Ready-to-Eat Seafood. Foods 2021, 10, 271. [Google Scholar] [CrossRef] [PubMed]
- Ačai, P.; Medved’Ová, A.; Mančušková, T.; Valík, L. Growth prediction of two bacterial populations in co-culture with lactic acid bacteria. Food Sci. Technol. Int. 2019, 25, 692–700. [Google Scholar] [CrossRef] [PubMed]
| Bacteriocin | Strain | Food Applications | Reference |
|---|---|---|---|
| Nisin | Lactococcus lactis | Milk | [16] |
| Lobster | [17] | ||
| Trout | [18] | ||
| Apple Cider | [19] | ||
| Liquid Whey | [20] | ||
| Lacticin | Lactococcus lactis | Milk | [21] |
| Pork sausage | [22] | ||
| Reuterin | Limosilactobacillus reuteri | Skim milk | [23] |
| Gassericin | Lactobacillus gasseri | Custard Cream | [24] |
| Lactococcin | Lactococcus lactis | Milk | [25] |
| Enterocin | Enteroccocus spp. | Apple juice | [19] |
| Ready to eat salad | [26] |
| Organic Acid | Example of Prominent LAB Producer | Example Food Pathogen Application | Application in Food |
|---|---|---|---|
| Lactic acid | Lactobacillus delbrueckii subsp. bulgaricus [27] | Pseudomonas spp. [28] | Sliced Salmon [28] |
| Formic acid | Lactococcus lactis subsp. cremoris [29] | Esherichia coli [30] Listeria spp. [30] Salmonella spp. [30] Clostridium perfringens [31] | Poultry [30] Animal Feed [30] Pork [31] |
| Succinic acid | Lactococcus lactis subsp. lactis [29] | Salmonella spp. [30] | Chicken meat [32] |
| Malic acid | Limosilactobacillus reuteri [33] | Staphylococcus [27,33] | Meat products [33] |
| Propionic acid | Lactococcus lactis subsp. lactis [29] | Campylobacter spp. [34] | Poultry Food [35] |
| Acetic acid | Lactobacillusacidophilus [29] | Pseudomonas spp. [28] | Sliced Salmon [28] |
| Butyric Acid | Lactobacillusacidophilus [29] | Salmonella spp. [36] | Poultry [36] |
| Foodborne Pathogen | Lactic Acid Bacteria | Reference |
|---|---|---|
| Staphylococcus aureus | Lactococcus spp., Pediococcus spp., Lactobacillus spp., Weissella spp., and Enterococcus spp. Lactobacillus curvatus, Lactiplantibacillus plantarum, Lactobacillus sakei, Pediococcus acidilactici, and Pediococcus pentosaceus (industrial products) Levilactobacillus brevis, Lactobacillus coryniformis, Lactobacillusparacasei, Lactobacillus paraplantarum, Leuconostoc mesenteroides, and Weisella halotolerans (traditional products) Enterococcus faecium QPII, Lactiplantibacillus plantarum CC10, Lactiplantibacillus plantarum TF711, Lacticaseibacillus rhamnosus IMC 501, Lacticaseibacillus paracasei IMC 502, Lactobacillus sakei KTU05-6, Lactobacillus helveticus KLDS 1.8701, Pediococcus acidilactici KTU05-7, Pediococcus pentosaceus KTU05-8, KTU05-9, and KTU05-10 | [45,46,47,48,49,50] |
| Listeria innocua | Lactobacillus curvatus, Lactiplantibacillus plantarum, Lactobacillus sakei, Levilactobacillus brevis, Lactobacillus coryniformis, Lacticaseibacillus paracasei, Lactobacillus paraplantarum, Leuconostoc mesenteroides, Pediococcus acidilactici, and Pediococcus pentosaceus (industrial products), Weisella halotolerans (traditional products) | [45,47] |
| Escherichia coli | Lactobacillus curvatus, Lactiplantibacillusplantarum, Lactobacillus sakei, Levilactobacillus brevis, Lactobacillus coryniformis, Lacticaseibacillus paracasei, Lactobacillus paraplantarum, Lactobacillus helveticus KLDS 1.8701, Limosilactobacillus reuteri, Leuconostoc mesenteroides, Pediococcus spp. Acidilactici, Pediococcus pentosaceus (industrial products), Weisella halotolerans (traditional products) | [45,47,50,51] |
| Salmonella enteritidis | Limosilactobacillus reuteri | [45,50,51,52] |
| Salmonella typhimurium | Levilactobacillus brevis CM22, Lactobacillus helveticus KLDS 1.8701, Pediococcus pentosaceus CM16 | |
| Salmonella cholerae | Levilactobacillus brevis CM22, Lactobacillus helveticus KLDS 1.8701, Pediococcus pentosaceus CM16 | [45,50,52] |
| Bacillus cereus | Lactobacillus sakei KTU05-6, Lacticaseibacillus rhamnosus IMC 501, and Lacticaseibacillus paracasei IMC 502, Lactiplantibacillus plantarum TF711, Pediococcus acidilactici KTU05-7, Pediococcus pentosaceus KTU05-8, KTU05-9, and KTU05-10 | [49,53] |
| Pseudomonas | Lactobacillus sakei KTU05-6, Lactobacillus curvatus, Lactiplantibacillus plantarum, Levilactobacillus brevis, Lactobacillus coryniformis, Lacticaseibacillus paracasei, Lactobacillus paraplantarum, Leuconostoc mesenteroides, Pediococcus acidilactici KTU05-7, Pediococcus pentosaceus KTU05-8, KTU05-9, and KTU05-10, Weisella halotolerans (traditional products) | [47] |
| Enterococcus faecium DSM 13590 | Lactobacillus sakei KTU05-6, Lacticaseibacillus rhamnosus IMC 501 and Lacticaseibacillus paracasei IMC 502, Pediococcus acidilactici KTU05-7, Pediococcus pentosaceus KTU05-8, KTU05-9, and KTU05-10 | [46,47] |
| Listeria monocytogenes | Enterococcus faecium QPII, Lactobacillus sakei KTU05-6, Lacticaseibacillus rhamnosus IMC 501 and Lacticaseibacillus paracasei IMC 502, Lactobacillus curvatus, Levilactobacillus brevis, Lactobacillus coryniformis, Lacticaseibacillus paracasei, Lactobacillus paraplantarum, Lactiplantibacillus plantarum CC10, Lactobacillus helveticus KLDS 1.8701, Limosilactobacillus reuteri, Leuconostoc mesenteroides, Pediococcus acidilactici KTU05-7, Pediococcus pentosaceus KTU05-8, KTU05-9, and KTU05-10, Weisella halotolerans (traditional products) | [46,48,50,54] |
| Clostridiumsporogenes | Lactiplantibacillus plantarum TF711 | [49] |
| Shigella sonnei | Lactiplantibacillus plantarum TF711 | [49] |
| Klebsiella pneumoniae | Lactiplantibacillus plantarum TF711 | [49] |
| Acinetobacter baumannii | Lactobacillus spp. | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, S.A.; Ayivi, R.D.; Zimmerman, T.; Siddiqui, S.A.; Altemimi, A.B.; Fidan, H.; Esatbeyoglu, T.; Bakhshayesh, R.V. Lactic Acid Bacteria as Antimicrobial Agents: Food Safety and Microbial Food Spoilage Prevention. Foods 2021, 10, 3131. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123131
Ibrahim SA, Ayivi RD, Zimmerman T, Siddiqui SA, Altemimi AB, Fidan H, Esatbeyoglu T, Bakhshayesh RV. Lactic Acid Bacteria as Antimicrobial Agents: Food Safety and Microbial Food Spoilage Prevention. Foods. 2021; 10(12):3131. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123131
Chicago/Turabian StyleIbrahim, Salam A., Raphael D. Ayivi, Tahl Zimmerman, Shahida Anusha Siddiqui, Ammar B. Altemimi, Hafize Fidan, Tuba Esatbeyoglu, and Reza Vaseghi Bakhshayesh. 2021. "Lactic Acid Bacteria as Antimicrobial Agents: Food Safety and Microbial Food Spoilage Prevention" Foods 10, no. 12: 3131. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123131