Effect of Phenotype Selection on Genome Size Variation in Two Species of Diptera

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. macellaria Colony Development
2.2. Drosophila Body Size Selection
2.3. Species Genome Size Estimation and Calculation of Cell-Count Ratio
2.4. Statistical Analysis
2.4.1. C. macellaria
2.4.2. D. melanogaster
3. Results
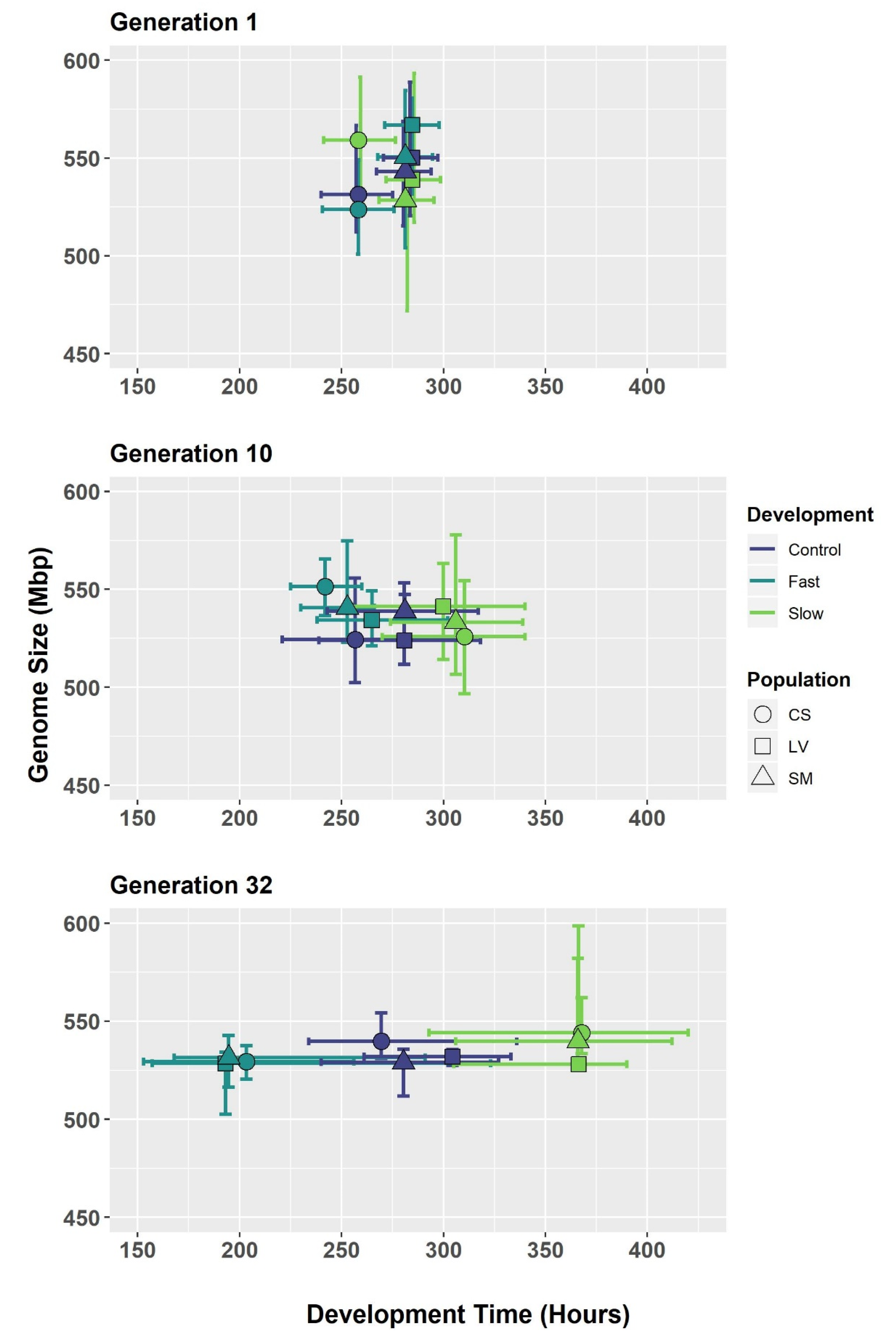
3.1. C. macellaria after 32 Generations of Selection
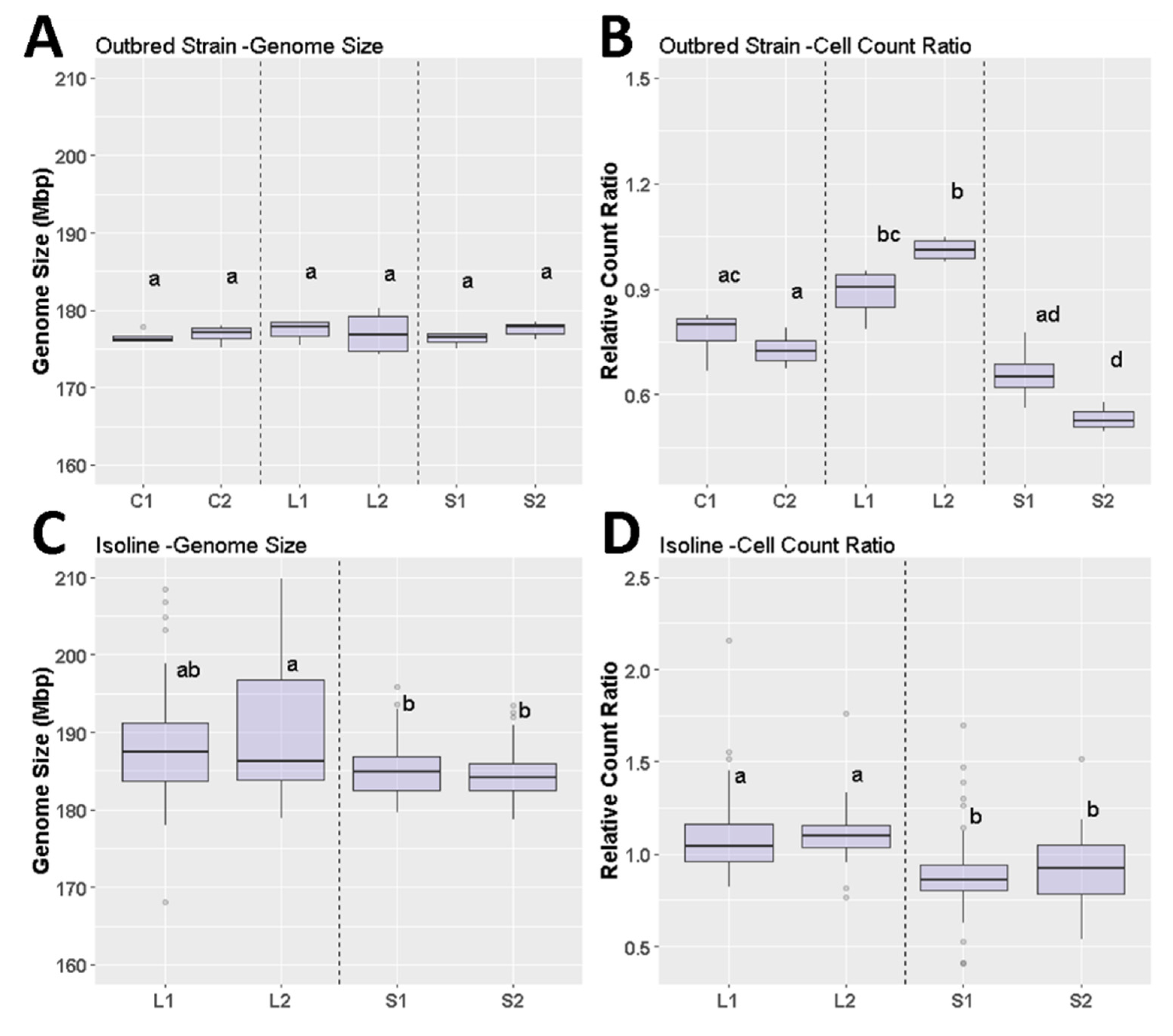
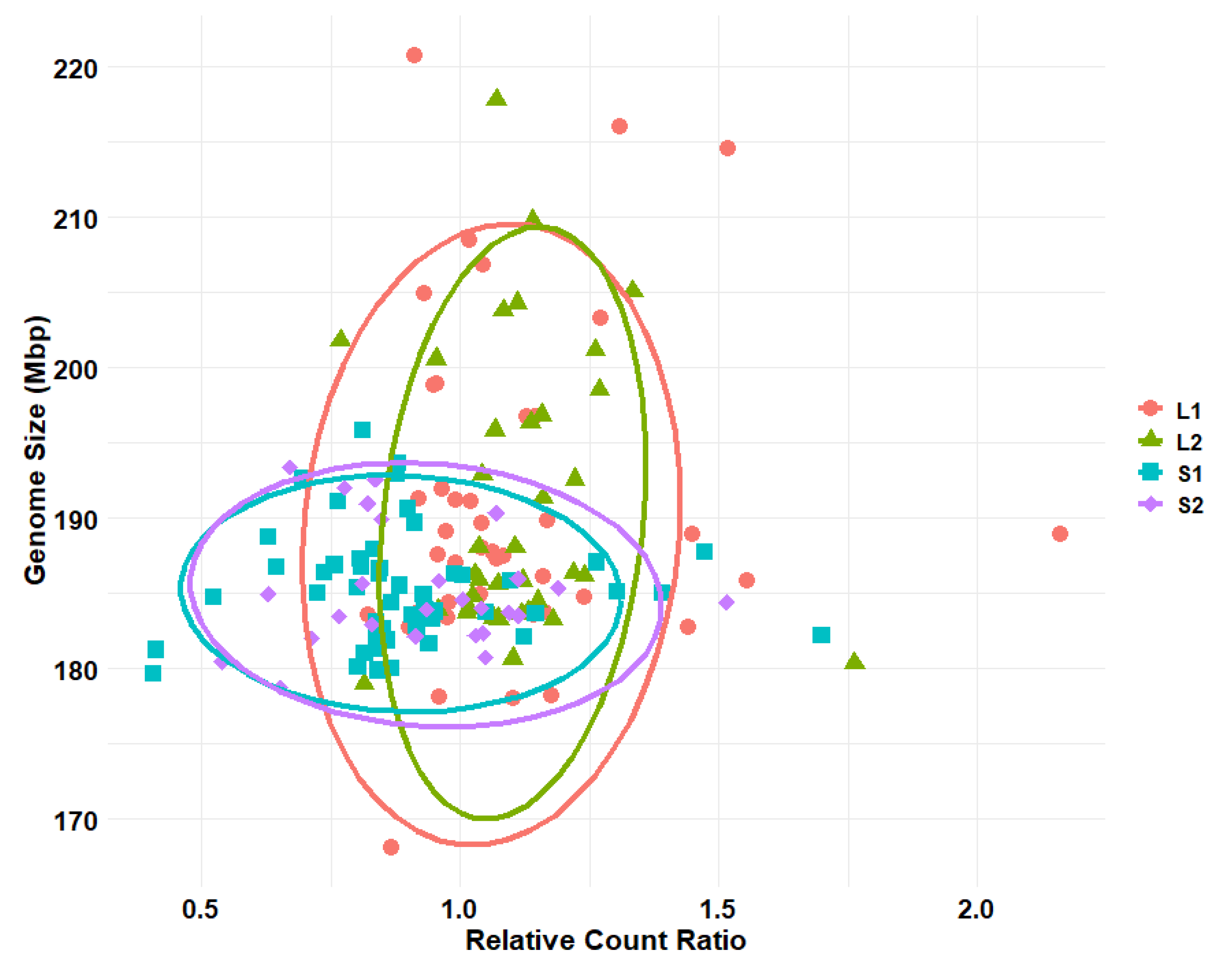
3.2. Drosophila Genome Size after Size Selection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oliver, M.J.; Petrov, D.; Ackerly, D.; Falkowski, P.; Schofield, O.M. The mode and tempo of genome size evolution in eukaryotes. Genome Res. 2007, 17, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Gregory, T.R. The Evolution of the Genome; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Gregory, T.R. The case for junk DNA. PLoS Genet 2014, 10, e1004351. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Massouras, A.; Inoue, Y.; Peiffer, J.; Ramia, M.; Tarone, A.M.; Turlapati, L.; Zichner, T.; Zhu, D.; Lyman, R.F.; et al. Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 2014, 24, 1193–1208. [Google Scholar] [CrossRef] [Green Version]
- Ellis, L.L.; Huang, W.; Quinn, A.M.; Ahuja, A.; Alfrejd, B.; Gomez, F.E.; Hjelmen, C.E.; Moore, K.L.; Mackay, T.F.C.; Johnston, J.S.; et al. Intrapopulation genome size in D. melanogaster reflects life history variation and plasticity. PLoS Genet. 2014, 10, e1004522. [Google Scholar] [CrossRef] [Green Version]
- Long, Q.; Rabanal, F.A.; Meng, D.; Huber, C.D.; Farlow, A.; Platzer, A.; Zhang, Q.; Vilhjálmsson, B.J.; Korte, A.; Nizhynska, V. Massive genomic variation and strong selection in Arabidopsis thaliana lines from Sweden. Nat. Genet. 2013, 45, 884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnqvist, G.; Sayadi, A.; Immonen, E.; Hotzy, C.; Rankin, D.; Tuda, M.; Hjelmen, C.E.; Johnston, J.S. Genome size correlates with reproductive fitness in seed beetles. Proc. R. Soc. B 2015, 282, 20151421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schielzeth, H.; Streitner, C.; Lampe, U.; Franzke, A.; Reinhold, K. Genome size variation affects song attractiveness in grasshoppers: Evidence for sexual selection against large genomes. Evolution 2014, 68, 3629–3635. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.I. Evolution of Genome Size. In eLS; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Gregory, T.R. Coincidence, coevolution, or causation? DNA content, cell size, and the C-value enigma. Biol. Rev. 2001, 76, 65–101. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Skeletal DNA and the evolution of genome size. Annu. Rev. Biophys. Bioeng. 1982, 11, 273–302. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. Economy, Speed and Size Matter: Evolutionary Forces Driving Nuclear Genome Miniaturization and Expansion. Ann. Bot. 2005, 95, 147–175. [Google Scholar] [CrossRef] [Green Version]
- Andrews, C.B.; Mackenzie, S.A.; Gregory, T.R. Genome size and wing parameters in passerine birds. Proc. R. Soc. B Biol. Sci. 2009, 276, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, J.M.; Leitch, I.J.; Patel, S.; Pendharkar, A.; Knight, C.A. Genome size is a strong predictor of cell size and stomatal density in angiosperms. New Phytol. 2008, 179, 975–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, T.R. The Bigger the C-Value, the Larger the Cell: Genome Size and Red Blood Cell Size in Vertebrates. Blood Cells Mol. Dis. 2001, 27, 830–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, T.R. A bird’s-eye view of the c-value enigma: Genome size, cell size, and metabolic rate in teh class Aves. Evolution 2002, 56, 121–130. [Google Scholar] [CrossRef]
- Knight, C.A.; Beaulieu, J.M. Genome Size Scaling through Phenotype Space. Ann. Bot. 2008, 101, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Calboli, F.C.; Gilchrist, G.W.; Partridge, L.; Fry, J. Different cell size and cell number contribution in two newly established and one ancient body size cline of Drosophila subobscura. Evolution 2003, 57, 566–573. [Google Scholar] [CrossRef]
- Gregory, T.R.; Hebert, P.D.; Kolasa, J. Evolutionary implications of the relationship between genome size and body size in flatworms and copepods. Heredity 2000, 84, 201. [Google Scholar] [CrossRef]
- Jeffery, N.W.; Yampolsky, L.; Gregory, T.R. Nuclear DNA content correlates with depth, body size, and diversification rate in amphipod crustaceans from ancient Lake Baikal, Russia. Genome 2016, 60, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Van’t Hof, J.; Sparrow, A.H. A relationship between DNA content, nuclear volume, and minimum mitotic cycle time. Proc. Natl. Acad. Sci. USA 1963, 49, 897. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.D. The Duration of Meiosis. Proc. R. Soc. Lond. B 1971, 178, 277–299. [Google Scholar] [CrossRef]
- Bennett, M.D. Nuclear DNA content and minimum generation time in herbaceous plants. Proc. R. Soc. Lond. B Biol. Sci. 1972, 181, 109–135. [Google Scholar] [CrossRef] [PubMed]
- Grosset, L.; Odartchenko, N. Duration of mitosis and separate mitotic phases compared to nuclear DNA content in erythroblasts of four vertebrates. Cell Prolif. 1975, 8, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ryan Gregory, T. Genome size and developmental complexity. Genetica 2002, 115, 131–146. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Schubert, V.; Meister, A.; Fuchs, J.; Schubert, I. Variation in genome size, cell and nucleus volume, chromosome number and rDNA loci among duckweeds. Sci. Rep. 2019, 9, 3234. [Google Scholar] [CrossRef]
- Wyngaard, G.A.; Rasch, E.M.; Manning, N.M.; Gasser, K.; Domangue, R. The relationship between genome size, development rate, and body size in copepods. Hydrobiologia 2005, 532, 123–137. [Google Scholar] [CrossRef]
- McLaren, I.; Sevigny, J.-M.; Corkett, C. Body sizes, development rates, and genome sizes among Calanus species. In Biology of Copepods; Springer: Berlin, Germany, 1988; pp. 275–284. [Google Scholar] [CrossRef]
- Finston, T.L.; Hebert, P.D.N.; Foottit, R.B. Genome size variation in aphids. Insect Biochem. Mol. Biol. 1995, 25, 189–196. [Google Scholar] [CrossRef]
- Powell, J.R. Progress and Prospects in Evolutionary Biology: The Drosophila Model (Oxford Series in Ecology & Evolution); Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Gregory, T.R. Genome size evolution in animals. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier: San Diego, CA, USA, 2005; pp. 3–87. [Google Scholar] [CrossRef]
- Gregory, T.R.; Johnston, J.S. Genome size diversity in the family Drosophilidae. Heredity 2008, 101, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Conery, J.S. The origins of genome complexity. Science 2003, 302, 1401–1404. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.L.; Stewart, A.D.; Fields, A.T.; Rice, W.R.; Tarone, A.M. Population-based resequencing of experimentally evolved populations reveals the genetic basis of body size variation in Drosophila melanogaster. PLoS Genet. 2011, 7, e1001336. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.D.; Rice, W.R. Arrest of sex-specific adaptation during the evolution of sexual dimorphism in Drosophila. Nat. Ecol. Evol. 2018, 2, 1507–1513. [Google Scholar] [CrossRef]
- Pischedda, A.; Friberg, U.; Stewart, A.D.; Miller, P.M.; Rice, W.R. Sexual selection has minimal impact on effective population sizes in species with high rates of random offspring mortality: An empirical demonstration using fitness distributions. Evolution 2015, 69, 2638–2647. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.S.; Bernardini, A.; Hjelmen, C.E. Genome Size Estimation and Quantitative Cytogenetics in Insects. In Insect Genomics; Springer: Berlin, Germany, 2019; pp. 15–26. [Google Scholar] [CrossRef]
- Picard, C.; Johnston, J.; Tarone, A. Genome sizes of forensically relevant Diptera. J. Med Entomol. 2012, 49, 192–197. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org (accessed on 1 September 2017).
- Schuetzenmeister, A.; Dufey, F. VCA-Package: Variance Component Analysis. R Package Version 1.4.2. 2019. Available online: https://CRAN.R-project.org/package=VCA (accessed on 10 January 2020).
- Xiao, Y. Peacock.test: Two and Three Dimensional Kolmogorov-Smirnov Two-Sample Tests. R Package Version. 1.0. 2016. Available online: https://CRAN.R-project.org/package=Peacock.test (accessed on 6 February 2020).
- Charlesworth, B.; Sniegowski, P.; Stephan, L. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 1994, 371, 215–220. [Google Scholar] [CrossRef]
- Athrey, N.; Leberg, P.L.; Klerks, P.L. Laboratory culturing and selection for increased resistance to cadmium reduce genetic variation in the least killifish, Heterandria formosa. Environ. Toxicol. Chem. 2009, 26, 1916–1921. [Google Scholar] [CrossRef]
- Cheng, G.W.; Breen, P.J. Cell-count and size in relation to fruit size among strawberry cultivars. J. Am. Soc. Hortic. Sci. 1992, 117, 946–950. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, B.; Barton, N. Genome size: Does bigger mean worse? Curr. Biol. 2004, 14, R233–R235. [Google Scholar] [CrossRef] [Green Version]
- Gregory, T.R.; Witt, J.D.S. Population size and genome size in fishes: A closer look. Genome 2008, 51, 309–313. [Google Scholar] [CrossRef]
- Black, W.C.T.; Rai, K.S. Genome evolution in mosquitoes: Intraspecific and interspecific variation in repetitive DNA amounts and organization. Genet. Res. 1988, 51, 185–196. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Kellogg, E.A. Do plants have a one-way ticket to genomic obesity? Plant Cell 1997, 9, 1509. [Google Scholar] [CrossRef]
- Ågren, J.A.; Wright, S.I. Co-evolution between transposable elements and their hosts: A major factor in genome size evolution? Chromosome Res. 2011, 19, 777–786. [Google Scholar] [CrossRef]
- Śliwińska, E.B.; Martyka, R.; Tryjanowski, P. Evolutionary interaction between W/Y chromosome and transposable elements. Genetica 2016, 144, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Lower, S.S.; Spencer Johnston, J.; Stanger-Hall, K.; Hjelmen, C.E.; Hanrahan, S.J.; Korunes, K.; Hall, D. Genome size in North American fireflies: Substantial variation likely driven by neutral processes. Genome Biol. Evol. 2017, 9, 1499–1512. [Google Scholar] [CrossRef]
- Bosco, G.; Campbell, P.; Leiva-Neto, J.T.; Markow, T.A. Analysis of Drosophila species genome size and satellite DNA content reveals significant differences among strains as well as between species. Genetics 2007, 177, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Trivers, R.; Burt, A.; Palestis, B.G. B chromosomes and genome size in flowering plants. Genome 2004, 47, 1–8. [Google Scholar] [CrossRef]
- Otto, S.P. The Evolutionary Consequences of Polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Force, A.G. The Origin of Interspecific Genomic Incompatibility via Gene Duplication. Am. Nat. 2000, 156, 590–605. [Google Scholar] [CrossRef]
- Tantawy, A.; Vetukhiv, M. Effects of size on fecundity, longevity and viability in populations of Drosophila pseudoobscura. Am. Nat. 1960, 94, 395–403. [Google Scholar] [CrossRef]
- Lefranc, A.; Bundgaard, J. The influence of male and female body size on copulation duration and fecundity in Drosophila melanogaster. Hereditas 2000, 132, 243–247. [Google Scholar] [CrossRef]
- Long, T.A.; Pischedda, A.; Stewart, A.D.; Rice, W.R. A cost of sexual attractiveness to high-fitness females. PLoS Biol. 2009, 7, e1000254. [Google Scholar] [CrossRef] [Green Version]
- Nijhout, H.F. The control of body size in insects. Dev. Biol. 2003, 261, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Petrov, D.A. DNA loss and evolution of genome size in Drosophila. Genetica 2002, 115, 81–91. [Google Scholar] [CrossRef]
- Petrov, D.A. Evolution of genome size: New approaches to an old problem. Trends Genet. 2001, 17, 23–28. [Google Scholar] [CrossRef]
- Petrov, D.A.; Hartl, D.L. High rate of DNA loss in the Drosophila melanogaster and Drosophila virilis species groups. Mol. Biol. Evol. 1998, 15, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Petrov, D.A.; Sangster, T.A.; Johnston, J.S.; Hartl, D.L.; Shaw, K.L. Evidence for DNA loss as a determinant of genome size. Science 2000, 287, 1060–1062. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, A.; Suh, A.; Feschotte, C. Dynamics of genome size evolution in birds and mammals. Proc. Natl. Acad. Sci. USA 2017, 114, E1460–E1469. [Google Scholar] [CrossRef] [Green Version]
- Kidwell, M.G. Transposable elements and the evolution of genome size in eukaryotes. Genetica 2002, 115, 49–63. [Google Scholar] [CrossRef]
- Sessegolo, C.; Burlet, N.; Haudry, A. Strong phylogenetic inertia on genome size and transposable element content among 26 species of flies. Biol. Lett. 2016, 12, 20160407. [Google Scholar] [CrossRef] [Green Version]
- Vieira, C.; Biemont, C. Transposable element dynamics in two sibling species: Drosophila melanogaster and Drosophila simulans. Genetica 2004, 120, 115–123. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Gen. | City | Selection | City*Selection | Error |
|---|---|---|---|---|---|
| Genome Size | 1 | 0 | 0 | 272.1 | 467.6 |
| 10 | 0 | 9.3 | 31.7 | 291.9 | |
| 32 | 8.6 | 5.7 | 17.4 | 83.5 | |
| Dev. Time | 1 | 211.7 | 0 | 0 | 258.7 |
| 10 | 0.4 | 684.8 | 114.7 | 169.7 | |
| 32 | 0 | 6929.8 | 152.8 | 482.3 |
| College Station | |||||||
| Genome Size | Development Time | ||||||
| Selection | Gen. | N | Mbp | St. Dev | N | Hours | St. Dev |
| Control | 1 | 11 | 531.26 | 18.33 | 1209 | 258.23 | 18.42 |
| 10 | 8 | 524.29 | 18.9 | 1038 | 256.62 | 13.59 | |
| 32 | 10 | 539.72 | 7.03 | 1161 | 269.53 | 15.85 | |
| Fast | 1 | 11 | 523.71 | 14.26 | 1209 | - | - |
| 10 | 3 | 551.35 | 14.42 | 1100 | 241.88 | 8.26 | |
| 32 | 10 | 529.37 | 5.44 | 988 | 203.29 | 27.52 | |
| Slow | 1 | 9 | 559.11 | 24.84 | 1209 | - | - |
| 10 | 10 | 525.76 | 18.26 | 1125 | 310.33 | 10.79 | |
| 32 | 10 | 544.08 | 9.38 | 996 | 367.75 | 20.97 | |
| Longview | |||||||
| Genome Size | Development Time | ||||||
| Selection | Gen. | N | Mbp | St. Dev | N | Hours | St. Dev |
| Control | 1 | 10 | 550.1 | 20.5 | 1003 | 284.61 | 14.61 |
| 10 | 9 | 523.9 | 12.45 | 986 | 280.73 | 17.55 | |
| 32 | 9 | 531.91 | 3.03 | 1068 | 304.4 | 14.62 | |
| Fast | 1 | 5 | 566.77 | 20.52 | 1003 | - | - |
| 10 | 5 | 534.32 | 11.6 | 1015 | 264.85 | 15.28 | |
| 32 | 10 | 528.53 | 9.63 | 1044 | 193.08 | 32.67 | |
| Slow | 1 | 6 | 538.92 | 27.41 | 1003 | - | - |
| 10 | 9 | 541.21 | 19.31 | 974 | 299.86 | 13.06 | |
| 32 | 10 | 527.99 | 11.27 | 1151 | 366.33 | 13.57 | |
| San Marcos | |||||||
| Genome Size | Development Time | ||||||
| Selection | Gen. | N | Mbp | St. Dev | N | Hours | St. Dev |
| Control | 1 | 9 | 543.15 | 18.37 | 1176 | 281.26 | 14.68 |
| 10 | 3 | 538.83 | 7.49 | 990 | 281.04 | 18.13 | |
| 32 | 10 | 529.05 | 7.02 | 1032 | 280.39 | 24.54 | |
| Fast | 1 | 10 | 550.58 | 23.28 | 1176 | - | - |
| 10 | 7 | 540.5 | 16.64 | 1026 | 252.86 | 6.32 | |
| 32 | 10 | 531.49 | 8.03 | 847 | 194.67 | 20.86 | |
| Slow | 1 | 12 | 528.45 | 26.06 | 1176 | - | - |
| 10 | 10 | 533.21 | 20 | 1013 | 306.03 | 10.03 | |
| 32 | 10 | 539.87 | 15.29 | 958 | 365.95 | 21.35 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hjelmen, C.E.; Parrott, J.J.; Srivastav, S.P.; McGuane, A.S.; Ellis, L.L.; Stewart, A.D.; Johnston, J.S.; Tarone, A.M. Effect of Phenotype Selection on Genome Size Variation in Two Species of Diptera. Genes 2020, 11, 218. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020218
Hjelmen CE, Parrott JJ, Srivastav SP, McGuane AS, Ellis LL, Stewart AD, Johnston JS, Tarone AM. Effect of Phenotype Selection on Genome Size Variation in Two Species of Diptera. Genes. 2020; 11(2):218. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020218
Chicago/Turabian StyleHjelmen, Carl E., Jonathan J. Parrott, Satyam P. Srivastav, Alexander S. McGuane, Lisa L. Ellis, Andrew D. Stewart, J. Spencer Johnston, and Aaron M. Tarone. 2020. "Effect of Phenotype Selection on Genome Size Variation in Two Species of Diptera" Genes 11, no. 2: 218. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020218