A New SNP in Rice Gene Encoding Pyruvate Phosphate Dikinase (PPDK) Associated with Floury Endosperm

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. Evaluation of Agronomic Traits and Grain Physicochemical Properties
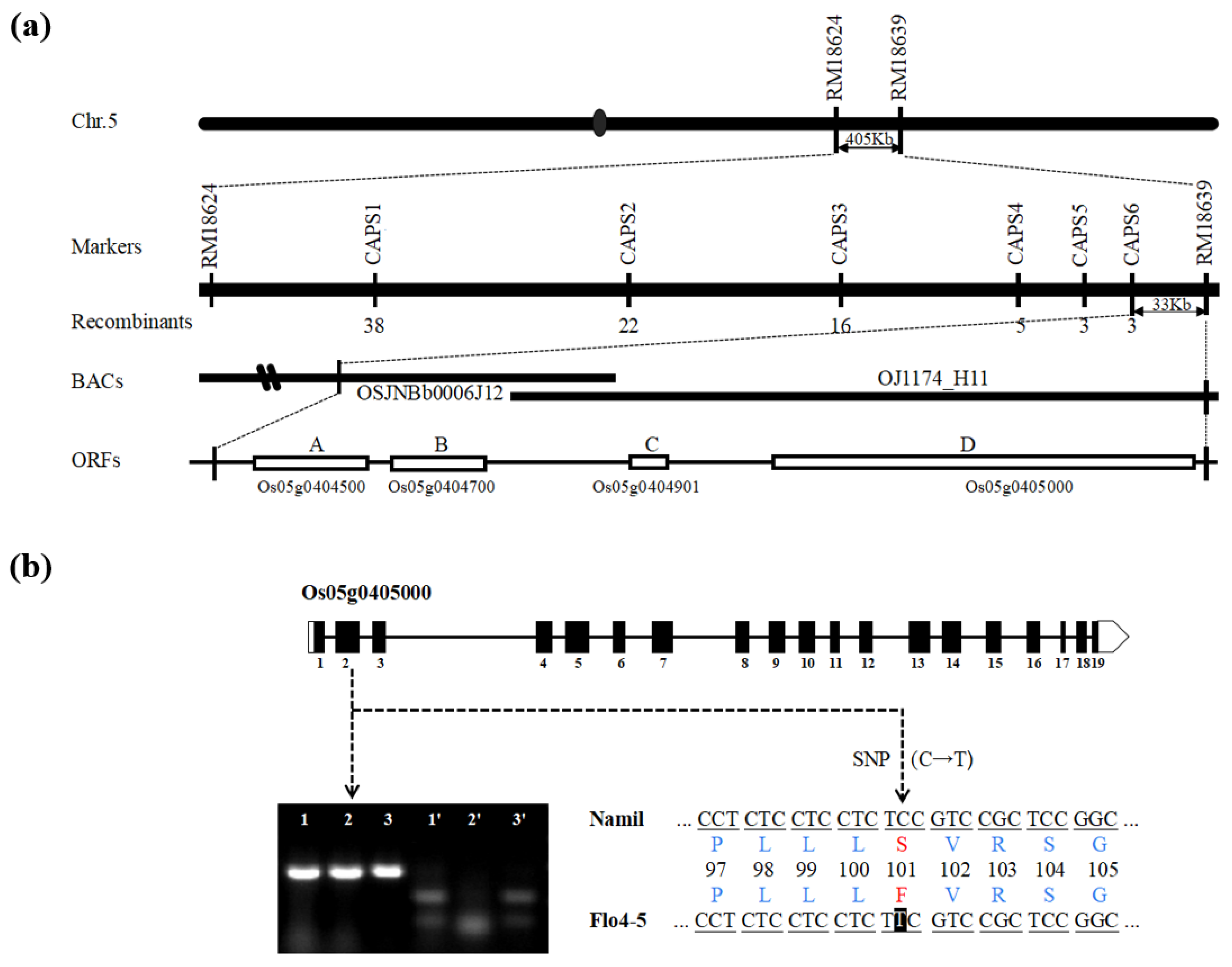
2.3. Fine Mapping of FLO4-5
2.4. Cloning of FLO4-5 and Identification of the Mutation Site
2.5. Predicting the Functional Effect of Amino Acid Substitutions and Real-Time qRT-PCR Analysis
3. Results
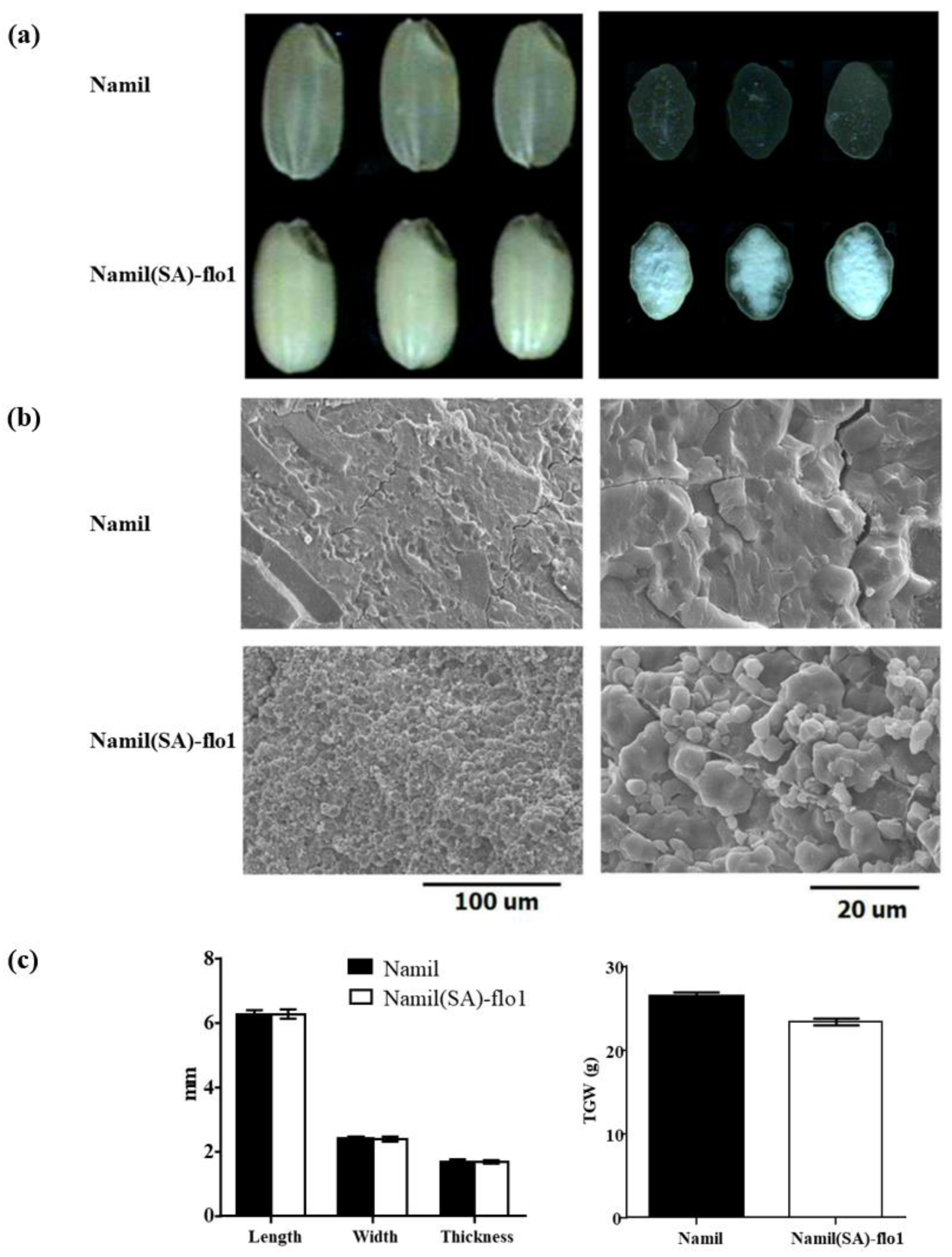
3.1. Agronomic Traits and Seed Characteristics of Namil(SA)-flo1
3.2. Dry Milling Properties of Namil(SA)-flo1
3.3. Fine Mapping of the Floury Endosperm Locus
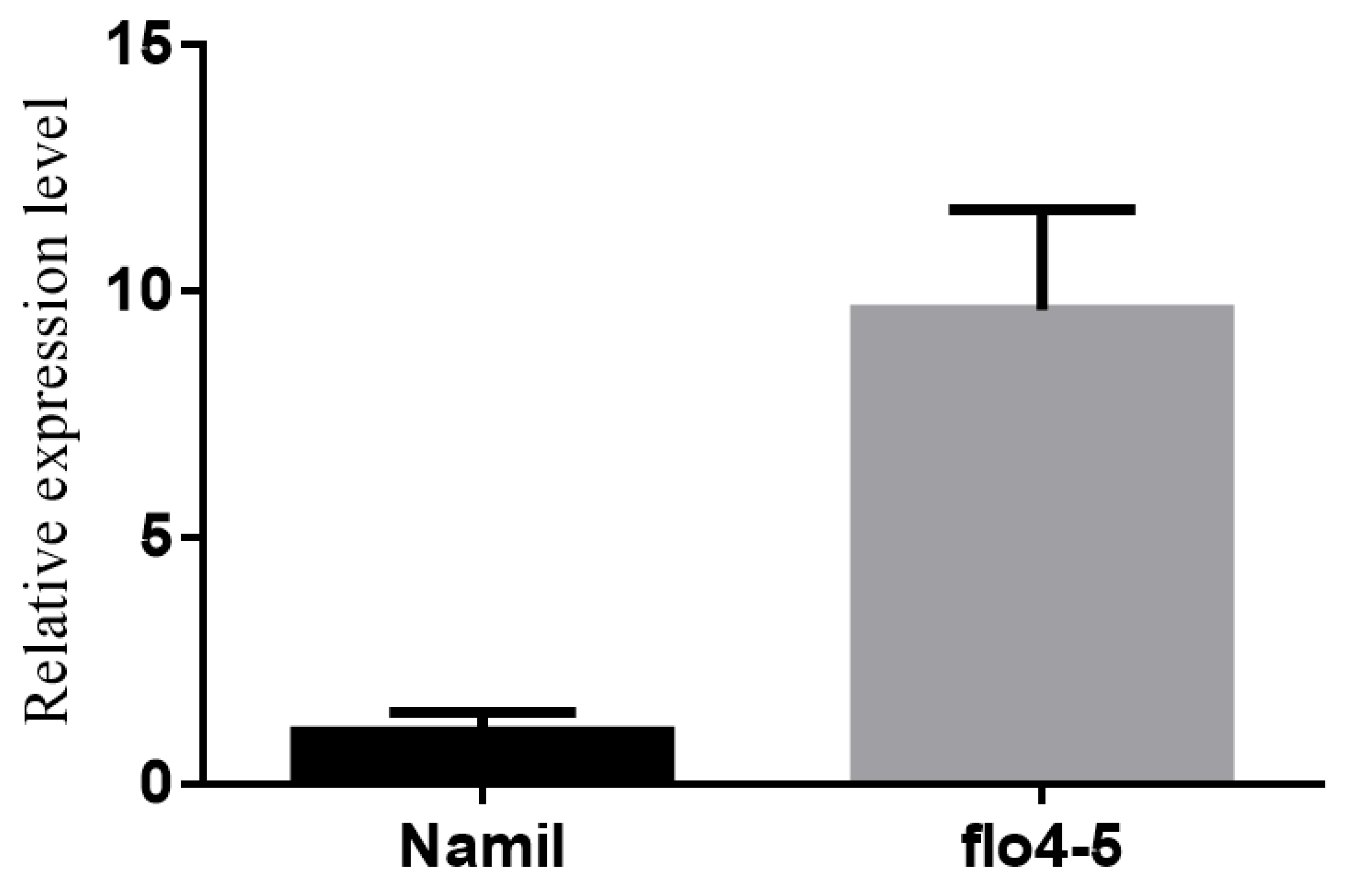
3.4. Co-Segregation and Expression Analyses
3.5. Mutation of FLO4-5 Changed the Expression Levels of Major Starch Synthesis Enzymes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tian, Z.; Liu, X.; Liu, G.; Wang, Y.; Li, J.; Qian, Q.; Yan, M.; Gao, Z.; Zeng, D.; Liu, Q.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, A.A.; Singh, P.; Shah, M.A.; Schweiggert-Weisz, U.; Gul, K.; Wani, I.A. Rice Starch Diversity: Effects on Structural, Morphological, Thermal, and Physicochemical Properties-A Review. Compr. Rev. Food Sci. Saf. 2012, 11, 417–436. [Google Scholar] [CrossRef]
- Wickramasinghe, H.A.M.; Noda, T. Physicochemical properties of starches from Sri Lankan rice varieties. Food Sci. Technol. Res. 2008, 14, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Whitt, S.R.; Buckler Iv, E.S.; Wilson, L.M.; Tenaillon, M.I.; Gaut, B.S. Genetic diversity and selection in the maize starch pathway. Proc. Natl. Acad. Sci. USA 2002, 99, 12959–12962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-K.; Hwang, S.-K.; Han, M.; Eom, J.-S.; Kang, H.-G.; Han, Y.; Choi, S.-B.; Cho, M.-H.; Bhoo, S.H.; An, G.; et al. Identification of the ADP-glucose pyrophosphorylase isoforms essential for starch synthesis in the leaf and seed endosperm of rice (Oryza sativa L.). Plant Mol. Biol. Rep. 2007, 65, 531–546. [Google Scholar] [CrossRef]
- Tang, X.-J.; Peng, C.; Zhang, J.; Cai, Y.; You, X.-M.; Kong, F.; Yan, H.-G.; Wang, G.-X.; Wang, L.; Jin, J.; et al. ADP-glucose pyrophosphorylase large subunit 2 is essential for storage substance accumulation and subunit interactions in rice endosperm. Plant Sci. 2016, 249, 70–83. [Google Scholar] [CrossRef]
- Li, N.; Zhang, S.; Zhao, Y.; Li, B.; Zhang, J. Over-expression of AGPase genes enhances seed weight and starch content in transgenic maize. Planta 2011, 233, 241–250. [Google Scholar] [CrossRef]
- Hanashiro, I.; Kuratomi, Y.; Matsugasako, J.I.; Takeda, Y.; Itoh, K.; Yamazaki, M.; Igarashi, T. Granule-bound starch synthase I is responsible for biosynthesis of extra-long unit chains of amylopectin in rice. Plant Cell Physiol. 2008, 49, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Sano, Y.; Maekawa, M.; Kikuchl, H. Temperature effects on the Wx protein level and amylose content in the endosperm of rice. J. Hered. 1985, 76, 221–222. [Google Scholar] [CrossRef]
- Wang, J.-C.; Xu, H.; Zhu, Y.; Liu, Q.-Q.; Cai, X.-L. OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J. Exp. Bot. 2013, 64, 3453–3466. [Google Scholar] [CrossRef]
- Dinges, J.R.; Colleoni, C.; Myers, A.M.; James, M.G. Molecular structure of three mutations at the maizesugary1 locus and their allele-specific phenotypic effects. Plant Physiol. 2001, 125, 1406–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Li, S.; Jiao, G.; Sheng, Z.; Wu, Y.; Shao, G.; Xie, L.; Peng, C.; Xu, J.; Tang, S. Os PK 2 encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, compound granule formation and grain filling. Plant Biotechnol. J. 2018, 16, 1878–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Omura, T. New Endosperm Mutations Induced by Chemical Mutagens in Rice Oryza sativa L. Jpn. J. Breed 1981, 31, 316–326. [Google Scholar] [CrossRef]
- Kang, H.G.; Park, S.; Matsuoka, M.; An, G. White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C4-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J. 2005, 42, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, R.; Khush, G. Genetic analysis of endosperm mutants in rice Oryza sativa L. Theor. Appl. Genet. 1991, 83, 146–152. [Google Scholar] [CrossRef]
- Nishio, T.; Iida, S. Mutants having a low content of 16-kDa allergenic protein in rice (Oryza sativa L.). Theor. Appl. Genet. 1993, 86, 317–321. [Google Scholar] [CrossRef]
- Ryoo, N.; Yu, C.; Park, C.-S.; Baik, M.-Y.; Park, I.M.; Cho, M.-H.; Bhoo, S.H.; An, G.; Hahn, T.-R.; Jeon, J.-S. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1083–1095. [Google Scholar] [CrossRef]
- She, K.-C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakata, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M. A novel factor FLOURY ENDOSPERM2 is involved in regulation of rice grain size and starch quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Wang, Y.; Liu, F.; Ren, Y.; Zhou, K.; Lv, J.; Zheng, M.; Zhao, S.; Zhang, L.; Wang, C. FLOURY ENDOSPERM 6 encodes a CBM 48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm. Plant J. 2014, 77, 917–930. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, Y.; Lu, B.; Yang, C.; Feng, Z.; Liu, Z.; Chen, J.; Ma, W.; Wang, Y.; Yu, X. FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice. J. Exp. Bot. 2016, 67, 633–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Wu, J.; Zhang, Y.; Shi, C. Phenotypic and candidate gene analysis of a new floury endosperm mutant (osagpl2-3) in rice. Mol Genet Genomics. 2012, 30, 1303–1312. [Google Scholar] [CrossRef]
- Crofts, N.; Abe, N.; Oitome, N.F.; Matsushima, R.; Hayashi, M.; Tetlow, I.J.; Emes, M.J.; Nakamura, Y.; Fujita, N. Amylopectin biosynthetic enzymes from developing rice seed form enzymatically active protein complexes. J. Exp. Bot. 2015, 66, 4469–4482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennen-Bierwagen, T.A.; Lin, Q.; Grimaud, F.; Planchot, V.; Keeling, P.L.; James, M.G.; Myers, A.M. Proteins from multiple metabolic pathways associate with starch biosynthetic enzymes in high molecular weight complexes: A model for regulation of carbon allocation in maize amyloplasts. Plant Physiol. 2009, 149, 1541–1559. [Google Scholar] [CrossRef] [Green Version]
- Ordonio, R.L.; Matsuoka, M. Increasing resistant starch content in rice for better consumer health. Proc. Natl. Acad. Sci. USA 2016, 113, 12616–12618. [Google Scholar] [CrossRef] [Green Version]
- Imaizumi, N.; Ku, M.S.; Ishihara, K.; Samejima, M.; Kaneko, S.; Matsuoka, M. Characterization of the gene for pyruvate, orthophosphate dikinase from rice, a C3 plant, and a comparison of structure and expression between C3 and C4 genes for this protein. Plant Mol. Biol. 1997, 34, 701–716. [Google Scholar] [CrossRef]
- Chastain, C.J.; Heck, J.W.; Colquhoun, T.A.; Voge, D.G.; Gu, X.-Y. Posttranslational regulation of pyruvate, orthophosphate dikinase in developing rice (Oryza sativa) seeds. Planta 2006, 224, 924–934. [Google Scholar] [CrossRef]
- Lappe, R.R.; Baier, J.W.; Boehlein, S.K.; Huffman, R.; Lin, Q.; Wattebled, F.; Settles, A.M.; Hannah, L.C.; Borisjuk, L.; Rolletschek, H. Functions of maize genes encoding pyruvate phosphate dikinase in developing endosperm. Proc. Natl. Acad. Sci. USA 2018, 115, E24–E33. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Huang, B.; Zhang, M.; Zhang, X.; Rivenbark, J.; Lappe, R.L.; James, M.G.; Myers, A.M.; Hennen-Bierwagen, T.A. Functional interactions between starch synthase III and isoamylase-type starch-debranching enzyme in maize endosperm. Plant physiol. 2012, 158, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Méchin, V.; Thévenot, C.; Le Guilloux, M.; Prioul, J.-L.; Damerval, C. Developmental analysis of maize endosperm proteome suggests a pivotal role for pyruvate orthophosphate dikinase. Plant physiol. 2007, 143, 1203–1219. [Google Scholar] [CrossRef] [Green Version]
- Moons, A.; Valcke, R.; Van, M. Low-oxygen stress and water deficit induce cytosolic pyruvate orthophosphate dikinase (PPDK) expression in roots of rice, a C3 plant. Plant J. 1998, 15, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.S.; Park, C.S.; Seo, Y.W.; Jeung, J.U. Characteristics of endosperm starch of the rice mutant lines induced by sodium azide. Korean J. Bredding Sci. 2009, 41, 84–91. [Google Scholar]
- Mo, Y.-J.; Jeung, J.-U.; Kang, K.-H.; Lee, J.-S.; Kim, B.-K. Genetic Analysis on Floury Endosperm Characteristics of’Namil (SA)-flo1’, a Japonica Rice Mutant Line. Korean J. Crop Sci. 2013, 58, 283–291. [Google Scholar] [CrossRef]
- Wang, H.; Mo, Y.-J.; Im, D.-E.; Jang, S.-G.; Ham, T.-H.; Lee, J.; Jeung, J.-U.; Kwon, S.-W. A new SNP in cyOsPPDK gene is associated with floury endosperm in Suweon 542. Mol. Genet. Genom. 2018, 293, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.-J.; Jeung, J.-U.; Shin, Y.-S.; Park, C.S.; Kang, K.-H.; Kim, B.-K. Agronomic and genetic analysis of Suweon 542, a rice floury mutant line suitable for dry milling. Rice 2013, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliano, B. A simplified assay for milled rice amylose. Cereal Sci. Today 1971, 16, 334–360. [Google Scholar]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Jeung, J.U.; Kim, B.R.; Cho, Y.C.; Han, S.S.; Moon, H.P.; Lee, Y.T.; Jena, K.K. A novel gene, Pi40(t), linked to the DNA markers derived from NBS-LRR motifs confers broad spectrum of blast resistance in rice. Theor. Appl. Genet 2007, 115, 1163–1177. [Google Scholar] [CrossRef]
- Rice Genome Annotation Project. Available online: http://rice.plantbiology.msu.edu/index.shtml (accessed on 23 April 2020).
- WatCut. Available online: http://watcut.uwaterloo.ca/template.php?act=snp_new (accessed on 23 April 2020).
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [Green Version]
- Rap-db. Available online: https://rapdb.dna.affrc.go.jp/index.html (accessed on 23 April 2020).
- Chastain, C.J.; Chollet, R. Regulation of pyruvate, orthophosphate dikinase by ADP-/Pi-dependent reversible phosphorylation in C3 and C4 plants. Plant Physiol. Biochem. 2003, 41, 523–532. [Google Scholar] [CrossRef]
- Huang, S.; Colmer, T.D.; Millar, A.H. Does anoxia tolerance involve altering the energy currency towards PPi? Trends Plant Sci. 2008, 13, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Shen, Z.; Sartor, R.; Wu, K.J.; Osborn, J.; Smith, L.G.; Briggs, S.P. Reconstruction of protein networks from an atlas of maize seed proteotypes. Proc. Natl. Acad. Sci. USA 2013, 110, E4808–E4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, N.; Yoshida, M.; Asakura, N.; Ohdan, T.; Miyao, A.; Hirochika, H.; Nakamura, Y. Function and characterization of starch synthase I using mutants in rice. Plant Physiol. 2006, 140, 1070–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, N.; Yoshida, M.; Kondo, T.; Saito, K.; Utsumi, Y.; Tokunaga, T.; Nishi, A.; Satoh, H.; Park, J.-H.; Jane, J.-L. Characterization of SSIIIa-deficient mutants of rice: The function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm. Plant Physiol. 2007, 144, 2009–2023. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Francisco, P.B.; Hosaka, Y.; Sato, A.; Sawada, T.; Kubo, A.; Fujita, N. Essential amino acids of starch synthase IIa differentiate amylopectin structure and starch quality between japonica and indica rice varieties. Plant Mol. Biol. Rep. 2005, 58, 213–227. [Google Scholar] [CrossRef]
- Umemoto, T.; Yano, M.; Satoh, H.; Shomura, A.; Nakamura, Y. Mapping of a gene responsible for the difference in amylopectin structure between japonica-type and indica-type rice varieties. Theor. Appl. Genet. 2002, 104, 1–8. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects ofamylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Yang, R.; Sun, C.; Bai, J.; Luo, Z.; Shi, B.; Zhang, J.; Yan, W.; Piao, Z. A putative gene sbe3-rs for resistant starch mutated from SBE3 for starch branching enzyme in rice (Oryza sativa L.). PLoS ONE 2012, 7, e43026. [Google Scholar] [CrossRef]
- Kubo, A.; Fujita, N.; Harada, K.; Matsuda, T.; Satoh, H.; Nakamura, Y. The starch-debranching enzymes isoamylase and pullulanase are both involved in amylopectin biosynthesis in rice endosperm. Plant Physiol. 1999, 121, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Kubo, A.; Rahman, S.; Utsumi, Y.; Li, Z.; Mukai, Y.; Yamamoto, M.; Ugaki, M.; Harada, K.; Satoh, H.; Konik-Rose, C. Complementation of sugary-1 phenotype in rice endosperm with the wheat isoamylase1 gene supports a direct role for isoamylase1 in amylopectin biosynthesis. Plant Physiol. 2005, 137, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Choi, M.-S.; Lee, G.; Jang, S.; Yoon, M.-R.; Kim, B.; Piao, R.; Woo, M.-O.; Chin, J.H.; Koh, H.-J. Sugary endosperm is modulated by starch branching enzyme IIa in rice (Oryza sativa L.). Rice 2017, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Cakir, B.; Shiraishi, S.; Tuncel, A.; Matsusaka, H.; Satoh, R.; Singh, S.; Crofts, N.; Hosaka, Y.; Fujita, N.; Hwang, S.-K. Analysis of the rice ADP-glucose transporter (OsBT1) indicates the presence of regulatory processes in the amyloplast stroma that control ADP-glucose flux into starch. Plant Physiol. 2016, 170, 1271–1283. [Google Scholar] [PubMed] [Green Version]
- Hannah, L.C.; Futch, B.; Bing, J.; Shaw, J.R.; Boehlein, S.; Stewart, J.D.; Beiriger, R.; Georgelis, N.; Greene, T. A shrunken-2 transgene increases maize yield by acting in maternal tissues to increase the frequency of seed development. Plant Cell. 2012, 24, 2352–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannah, L.C.; Shaw, J.R.; Clancy, M.A.; Georgelis, N.; Boehlein, S.K. A brittle-2 transgene increases maize yield by acting in maternal tissues to increase seed number. Plant Direct. 2017, 1, e00029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, D.M.; Timmerman, K.P.; Barry, G.F.; Preiss, J.; Kishore, G.M. Regulation of the amount of starch in plant tissues by ADP glucose pyrophosphorylase. Science 1992, 258, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Smidansky, E.D.; Clancy, M.; Meyer, F.D.; Lanning, S.P.; Blake, N.K.; Talbert, L.E.; Giroux, M.J. Enhanced ADP-glucose pyrophosphorylase activity in wheat endosperm increases seed yield. Proc. Natl. Acad. Sci. USA 2002, 99, 1724–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smidansky, E.D.; Martin, J.M.; Hannah, C.L.; Fischer, A.M.; Giroux, M.J. Seed yield and plant biomass increases in rice are conferred by deregulation of endosperm ADP-glucose pyrophosphorylase. Planta 2003, 216, 656–664. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | HD (Days) | CL (cm) | PL (cm) | TN (No.) | SN (No.) | RGP (%) |
|---|---|---|---|---|---|---|
| Namil | 101 b | 78 b | 25 a | 11 a | 117 a | 87 a |
| Namil(SA)-flo1 | 104 a | 82 a | 25 a | 10 a | 120 a | 74 b |
| Line | Hardness Index | Grain Hardness (Kg) | Mean Particle Size (µm) | Damaged Starch (%) | Lightness (CIE Value) | Ash (%) | Protein (%) | Amylose (%) | Lipid (%) |
|---|---|---|---|---|---|---|---|---|---|
| Hwaseong | 1.04 a | 7825 a | 112.2 ± 0.40 a | 10.3 ± 0.19 a | 88.6 ± 0.01 b | 0.84 ± 0.02 a | 7.5 ± 0.16 c | 18.5 ± 0.24 | - |
| Seolgaeng | 0.79 b | 5962 b | 97.6 ± 1.63 c | 7.1 ± 0.10 c | 90.3 ± 0.06 a | 0.72 ± 0.01 c | 6.6 ± 0.11 d | 17.5 ± 0.60 | - |
| Namil | 1.00 a | 7526 a | 109.1 ± 0.62 b | 9.2 ± 0.17 b | 88.7 ± 0.12 b | 0.82 ± 0.01 a | 9.2 ± 0.25 a | 17.7 ± 1.34 | 1.5 ± 0.56 b |
| Namil(SA)-flo1 | 0.45 c | 3417 c | 86.1 ± 0.81 d | 5.1 ± 0.06 d | 90.4 ± 0.09 a | 0.77 ± 0.02 b | 7.8 ± 0.04 b | 17.8 ± 0.27 | 3.1 ± 0.54 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Ham, T.-H.; Im, D.-E.; Lar, S.M.; Jang, S.-G.; Lee, J.; Mo, Y.; Jeung, J.-U.; Kim, S.T.; Kwon, S.-W. A New SNP in Rice Gene Encoding Pyruvate Phosphate Dikinase (PPDK) Associated with Floury Endosperm. Genes 2020, 11, 465. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040465
Wang H, Ham T-H, Im D-E, Lar SM, Jang S-G, Lee J, Mo Y, Jeung J-U, Kim ST, Kwon S-W. A New SNP in Rice Gene Encoding Pyruvate Phosphate Dikinase (PPDK) Associated with Floury Endosperm. Genes. 2020; 11(4):465. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040465
Chicago/Turabian StyleWang, Heng, Tae-Ho Ham, Da-Eun Im, San Mar Lar, Seong-Gyu Jang, Joohyun Lee, Youngjun Mo, Ji-Ung Jeung, Sun Tae Kim, and Soon-Wook Kwon. 2020. "A New SNP in Rice Gene Encoding Pyruvate Phosphate Dikinase (PPDK) Associated with Floury Endosperm" Genes 11, no. 4: 465. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040465