Distribution of the Warmblood Fragile Foal Syndrome Type 1 Mutation (PLOD1 c.2032G>A) in Different Horse Breeds from Europe and the United States

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Description Population Screening
2.2. DNA Isolation and Genotyping of WFFS
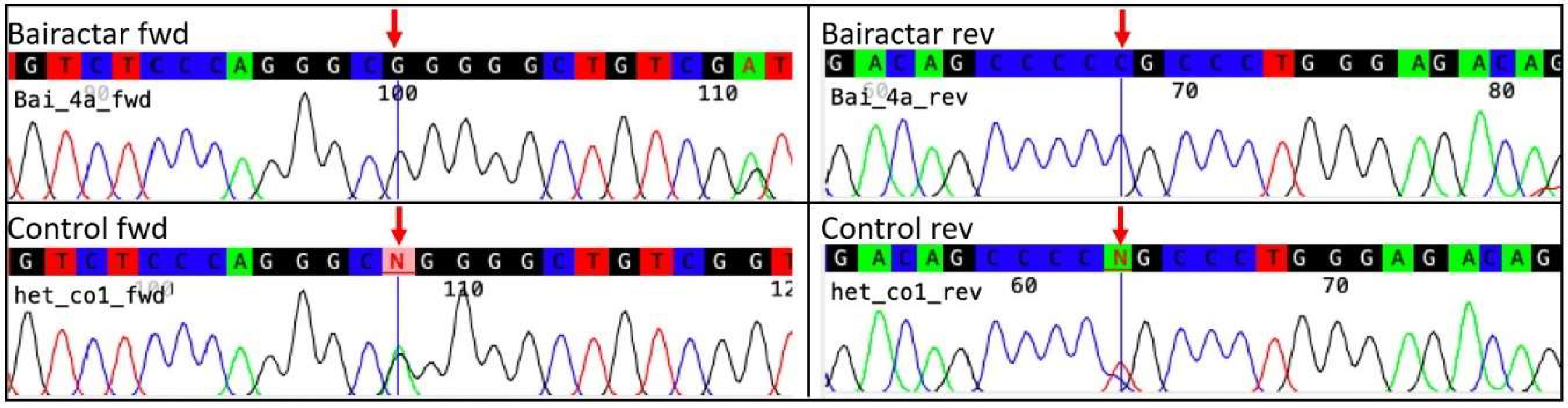
2.3. Historic DNA—Bairactar Or. Ar.
3. Results
3.1. Distribution of the WFFS Allele

{kind=link}
{kind=link}
| Breed | Total | Carriers | Carrier Frequency % | Allele Frequency % | 95% CI of WFFS Allele Frequency | Breed Information |
|---|---|---|---|---|---|---|
| Akhal Teke | 35 | 0 | 0 | 0 | ||
| American Sport Pony | 12 | 1 | 8.33 | 4.17 | Warmblood-derived, studbook not closed | |
| American Warmblood | 57 | 8 | 14.04 | 7.02 | 3.41 to 13.43 | Warmblood |
| Appaloosa | 43 | 0 | 0 | 0 | ||
| Arabian | 302 | 0 | 0 | 0 | ||
| Baden-Württemberger | 3 | 1 | 33.33 | 16.67 | Warmblood | |
| Belgian Sport Horse | 10 | 1 | 10.00 | 5.00 | Warmblood | |
| Belgian Warmblood | 44 | 5 | 11.36 | 5.68 | 2.14 to 12.93 | Warmblood |
| Canadian Warmblood | 29 | 3 | 10.34 | 5.17 | Warmblood | |
| Danish Warmblood | 127 | 22 | 17.32 | 8.66 | 5.74 to 12.82 | Warmblood |
| Dutch Warmblood | 249 | 19 | 7.63 | 3.82 | 2.42 to 5.92 | Warmblood |
| Friesian | 197 | 0 | 0 | 0 | ||
| Friesian Cross | 72 | 0 | 0 | 0 | ||
| Haflinger | 48 | 2 | 4.17 | 2.08 | 0.12 to 7.74 | Origin in Europe with influence from several breeds including Arabians, studbook not closed until 1946 |
| Hanoverian | 283 | 49 | 17.31 | 9.01 | 6.90 to 11.67 | Warmblood |
| Polish Heavy Draft | 209 | 0 | 0 | 0 | ||
| Hessen | 2 | 1 | 50.00 | 25.00 | Warmblood | |
| Holsteiner | 132 | 11 | 8.33 | 4.17 | 2.26 to 7.39 | Warmblood |
| Hucul | 146 | 0 | 0 | 0 | ||
| Knabstrupper | 46 | 3 | 6.52 | 3.26 | 0.72 to 9.55 | Baroque type; Warmblood crosses allowed, studbook not closed |
| Lesser Poland Warmblood | 157 | 3 | 1.91 | 0.96 | 0.19 to 2.91 | Warmblood |
| Lippizan | 42 | 0 | 0 | 0 | ||
| Norwegian Fjord | 42 | 0 | 0 | 0 | ||
| Oldenburg | 219 | 34 | 15.53 | 7.76 | 5.58 to 10.68 | Warmblood |
| Polish Konik | 96 | 0 | 0 | 0 | ||
| Quarter Horse | 112 | 0 | 0 | 0 | ||
| Rheinland | 12 | 2 | 16.67 | 8.33 | Warmblood | |
| Rocky Mountain Horse | 89 | 0 | 0 | 0 | ||
| Selle Français | 52 | 3 | 5.77 | 2.88 | 0.62 to 8.50 | Warmblood |
| Shetland Pony | 40 | 0 | 0 | 0 | ||
| Shire | 39 | 0 | 0 | 0 | ||
| Silesian Horse | 96 | 12 | 12.50 | 6.25 | 3.51 to 10.71 | Warmblood |
| Swedish Warmblood | 16 | 0 | 0 | 0 | ||
| Tennessee Walker | 39 | 0 | 0 | 0 | ||
| Thoroughbred * | 146 | 0 | 0 | 0 | ||
| Thoroughbred ** | 716 | 17 | 2.37 | 1.19 | 0.73 to 1.91 | Arabian, Barb, and Turkoman ancestry |
| Trakehner | 64 | 1 | 1.56 | 0.78 | 0.01 to 4.73 | Warmblood |
| Westfalen | 47 | 2 | 4.26 | 2.13 | 0.12 to 7.89 | Warmblood |
| Zangersheide | 11 | 0 | 0 | 0 | ||
| total | 4081 | 200 | 4.90 | 2.47 | 2.16 to 2.84 |
3.2. Historic DNA–Bairactar Or. Ar.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ehlers-Danlos Syndrome. Available online: https://ghr.nlm.nih.gov/condition/ehlers-danlos-syndrome (accessed on 17 September 2020).
- Malfait, F.; De Paepe, A. The Ehlers-Danlos syndrome. Adv. Exp. Med. Biol. 2014, 802, 129–143. [Google Scholar] [CrossRef]
- Halper, J. Connective tissue disorders in domestic animals. Adv. Exp. Med. Biol. 2014, 802, 231–240. [Google Scholar] [CrossRef]
- Eßer, M.; Niederacher, V.; Pfeffer, K.; Scheuer, H. Über die selten auftretende Dermatosparaxie (Ehlers-Danlos-Syndrom) bei einem Fohlen—Ein Fallbericht. Pferdeheilkunde 1999, 15, 434–436. [Google Scholar] [CrossRef]
- Witzig, P.; Suter, M.; Wild, P.; Rao, V.H.; Steinmann, B.; von Rotz, A. Dermatosparaxis in a foal and a cow—A rare disease? Schweiz. Arch. Tierheilkd. 1984, 126, 589–596. [Google Scholar]
- Marshall, V.L.; Secombe, C.; Nicholls, P.K. Cutaneous asthenia in a Warmblood foal. Aust. Vet. J. 2011, 89, 77–81. [Google Scholar] [CrossRef]
- Rüfenacht, S.; Straub, R.; Steinmann, B.; Winand, N.; Bidaut, A.; Stoffel, M.H.; Gerber, V.; Wyder, M.; Müller, E.; Roosje, P.J. Swiss warmblood horse with symptoms of hereditary equine regional dermal asthenia without mutation in the cyclophylin B gene (PPIB). Schweiz. Arch. Tierheilkd. 2010, 152, 188–192. [Google Scholar] [CrossRef]
- Winter, T.; Börgel, C.; Aupperle, H.; Schoon, H. Eine dem Ehlers-Danlos Syndrom des Menschen ähnliche Bindegewebserkrankung der Haut bei einem Fohlen. Pferdeheilkunde 2004, 20, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Rueff, A.G. Eigenthümliche Krankheit neugeborener Fohlen (a peculiar disease of newborn foals). In Jahrbuch für Pferdezucht; Verlag Voigt: Weimar, Germany, 1855; pp. 267–277. [Google Scholar]
- Gunson, D.E.; Halliwell, R.E.; Minor, R.R. Dermal collagen degradation and phagocytosis. Occurrence in a horse with hyperextensible fragile skin. Arch. Dermatol. 1984, 120, 599–604. [Google Scholar] [CrossRef]
- Rashmir-Raven, A. Heritable equine regional dermal asthenia. Vet. Clin. N. Am. Equine Pract. 2013, 29, 689–702. [Google Scholar] [CrossRef]
- White, S.D.; Affolter, V.K.; Bannasch, D.L.; Schultheiss, P.C.; Hamar, D.W.; Chapman, P.L.; Naydan, D.; Spier, S.J.; Rosychuk, R.A.W.; Rees, C.; et al. Hereditary equine regional dermal asthenia (“hyperelastosis cutis”) in 50 horses: Clinical, histological, immunohistological and ultrastructural findings. Vet. Dermatol. 2004, 15, 207–217. [Google Scholar] [CrossRef]
- Steelman, S.M.; Jackson, N.D.; Conant, E.; Juras, R.; Cothran, E.G.; Edwards, J.F.; Chowdhary, B.P. Ehlers-danlos syndrome in a quarter horse gelding: A case report of PPIB-independent hereditary equine regional dermal asthenia. J. Equine Vet. Sci. 2014, 34, 565–568. [Google Scholar] [CrossRef]
- Borges, A.S.; Conceição, L.G.; Alves, A.L.G.; Fabris, V.E.; Pessoa, M.A. Hereditary equine regional dermal asthenia in three related Quarter horses in Brazil. Vet. Dermatol. 2005, 16, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Lerner, D.; McCracken, M. Hyperelastosis cutis in 2 Horses. J. Equine Med. Surg. 1978, 2, 350–352. [Google Scholar]
- White, S.D.; Affolter, V.K.; Schultheiss, P.C.; Ball, B.A.; Wessel, M.T.; Kass, P.; Molinaro, A.M.; Bannasch, D.L.; Ihrke, P.J. Clinical and pathological findings in a HERDA-affected foal for 1.5 years of life. Vet. Dermatol. 2007, 18, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H.; Fisher, K.R.; Vrablic, O.E.; Yager, J.A.; Nimmo-Wilkie, J.S.; Parker, W.; Keeley, F.W. An inherited connective tissue disease in the horse. Lab. Investig. 1988, 59, 253–262. [Google Scholar] [PubMed]
- Brounts, S.H.; Rashmir-Raven, A.M.; Black, S.S. Zonal dermal separation: A distinctive histopathological lesion associated with hyperelastosis cutis in a Quarter Horse. Vet. Dermatol. 2001, 12, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Bowser, J.E.; Elder, S.H.; Pasquali, M.; Grady, J.G.; Rashmir-Raven, A.M.; Wills, R.; Swiderski, C.E. Tensile properties in collagen-rich tissues of Quarter Horses with hereditary equine regional dermal asthenia (HERDA). Equine Vet. J. 2014, 46, 216–222. [Google Scholar] [CrossRef]
- Grady, J.G.; Elder, S.H.; Ryan, P.L.; Swiderski, C.E.; Rashmir-Raven, A.M. Biomechanical and molecular characteristics of hereditary equine regional dermal asthenia in Quarter Horses. Vet. Dermatol. 2009, 20, 591–599. [Google Scholar] [CrossRef]
- Mochal, C.A.; Miller, W.W.; Cooley, A.J.; Linford, R.L.; Ryan, P.L.; Rashmir-Raven, A.M. Ocular findings in Quarter Horses with hereditary equine regional dermal asthenia. J. Am. Vet. Med. Assoc. 2010, 237, 304–310. [Google Scholar] [CrossRef]
- Tryon, R.C.; Penedo, M.C.T.; McCue, M.E.; Valberg, S.J.; Mickelson, J.R.; Famula, T.R.; Wagner, M.L.; Jackson, M.; Hamilton, M.J.; Nooteboom, S.; et al. Evaluation of allele frequencies of inherited disease genes in subgroups of American Quarter Horses. J. Am. Vet. Med. Assoc. 2009, 234, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Finno, C.J.; Spier, S.J.; Valberg, S.J. Equine diseases caused by known genetic mutations. Vet. J. 2009, 179, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Solomons, B. Equine cutis hyperelastica. Equine Vet. J. 1984, 16, 541–542. [Google Scholar] [CrossRef] [PubMed]
- Tryon, R.C.; White, S.D.; Bannasch, D.L. Homozygosity mapping approach identifies a missense mutation in equine cyclophilin B (PPIB) associated with HERDA in the American Quarter Horse. Genomics 2007, 90, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monthoux, C.; de Brot, S.; Jackson, M.; Bleul, U.; Walter, J. Skin malformations in a neonatal foal tested homozygous positive for Warmblood Fragile Foal Syndrome. BMC Vet. Res. 2015, 11, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winand, N.J. Identification of the Causative Mutation for Inherited Connective Tissue Disorders in Equines, United States Department of Commerce Application Number: 61/486,464; (Filing Date: May 16th, 2011). Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2012158711&recNum=1&maxRec=1&office=&prevFilter=&sortOption=Pub+Date+Desc&queryString=FP%3A%28WO2012158711%29&tab=PCT+Biblio (accessed on 20 September 2020).
- Aurich, C.; Müller-Herbst, S.; Reineking, W.; Müller, E.; Wohlsein, P.; Gunreben, B.; Aurich, J. Characterization of abortion, stillbirth and non-viable foals homozygous for the Warmblood Fragile Foal Syndrome. Anim. Reprod. Sci. 2019, 211, 106202. [Google Scholar] [CrossRef] [PubMed]
- Metzger, J.; Kreft, O.; Sieme, H.; Martinsson, G.; Reineking, W.; Hewicker-Trautwein, M.; Distl, O. Hanoverian F/W-line contributes to segregation of Warmblood fragile foal syndrome type 1 variant PLOD1:c.2032G>A in Warmblood horses. Equine Vet. J. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Bellone, R.R.; Ocampo, N.R.; Hughes, S.S.; Le, V.; Arthur, R.; Finno, C.J.; Penedo, M.C.T. Warmblood fragile foal syndrome type 1 mutation (PLOD1 c.2032G>A) is not associated with catastrophic breakdown and has a low allele frequency in the Thoroughbred breed. Equine Vet. J. 2020, 52, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Wege zum Angemessenen Umgang mit WFFS und Anderen Genetischen Eigenschaften. Available online: https://www.vit.de/aktuelles/20191/wege-zum-angemessenen-umgang-mit-wffs-und-anderen-genetischen-eigenschaften/ (accessed on 17 December 2020).
- Zhang, X.; Hirschfeld, M.; Schafberg, R.; Swalve, H.; Brenig, B. Skin exhibits of Dark Ronald XX are homozygous wild type at the Warmblood fragile foal syndrome causative missense variant position in lysyl hydroxylase gene PLOD1. Anim. Genet. 2020, 1–2. [Google Scholar] [CrossRef]
- Locke, M.M.; Penedo, M.C.T.; Bricker, S.J.; Millon, L.V.; Murray, J.D. Linkage of the grey coat colour locus to microsatellites on horse chromosome 25. Anim. Genet. 2002, 33, 329–337. [Google Scholar] [CrossRef]
- Agrest, A.; Coull, B.S. Approximate is Better than “Exact” for Interval Estimation of Binomial Proportions. Am. Stat. 1998, 52, 119–126. [Google Scholar] [CrossRef]
- Dias, N.M.; de Andrade, D.G.A.; Teixeira-Neto, A.R.; Trinque, C.M.; de Oliveira-Filho, J.P.; Winand, N.J.; Araújo, J.P.; Borges, A.S. Warmblood Fragile Foal Syndrome causative single nucleotide polymorphism frequency in Warmblood horses in Brazil. Vet. J. 2019, 248, 101–102. [Google Scholar] [CrossRef]
- Sandoval-Castellanos, E.; Wutke, S.; Gonzalez-Salazar, C.; Ludwig, A. Coat colour adaptation of post-glacial horses to increasing forest vegetation. Nat. Ecol. Evol. 2017, 1, 1816–1819. [Google Scholar] [CrossRef]
- Georgescu, S.; Manea, M.; Dudu, A.; Costache, M.; Mol, G. Phylogenetic relationships of the Hucul horse from Romania inferred from mitochondrial D-loop variation. Genet. Mol. Res. 2011, 10, 4104–4113. [Google Scholar] [CrossRef]
- Polak, G. Genetic variability of cold-blooded horses participating in genetic resources conservation programs, using pedigree analysis. Ann. Anim. Sci. 2019, 19, 49–60. [Google Scholar] [CrossRef]
- Fung, T.; Kennan, K. General case of sampling form a finite diploid population of any size. PLoS ONE 2014, 9, e85925. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reiter, S.; Wallner, B.; Brem, G.; Haring, E.; Hoelzle, L.; Stefaniuk-Szmukier, M.; Długosz, B.; Piórkowska, K.; Ropka-Molik, K.; Malvick, J.; et al. Distribution of the Warmblood Fragile Foal Syndrome Type 1 Mutation (PLOD1 c.2032G>A) in Different Horse Breeds from Europe and the United States. Genes 2020, 11, 1518. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121518
Reiter S, Wallner B, Brem G, Haring E, Hoelzle L, Stefaniuk-Szmukier M, Długosz B, Piórkowska K, Ropka-Molik K, Malvick J, et al. Distribution of the Warmblood Fragile Foal Syndrome Type 1 Mutation (PLOD1 c.2032G>A) in Different Horse Breeds from Europe and the United States. Genes. 2020; 11(12):1518. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121518
Chicago/Turabian StyleReiter, Simone, Barbara Wallner, Gottfried Brem, Elisabeth Haring, Ludwig Hoelzle, Monika Stefaniuk-Szmukier, Bogusława Długosz, Katarzyna Piórkowska, Katarzyna Ropka-Molik, Julia Malvick, and et al. 2020. "Distribution of the Warmblood Fragile Foal Syndrome Type 1 Mutation (PLOD1 c.2032G>A) in Different Horse Breeds from Europe and the United States" Genes 11, no. 12: 1518. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121518