
Metabolic Capacity of the Antarctic Cyanobacterium Phormidium pseudopriestleyi That Sustains Oxygenic Photosynthesis in the Presence of Hydrogen Sulfide

 , ,
, ,
Abstract
:1. Introduction

Site Description
2. Materials and Methods
2.1. Sulfide Concentrations
2.2. Field Work
2.3. DNA Extraction and Sequencing
2.4. Bioinformatics Analysis
3. Results
3.1. Sulfide Concentrations
3.2. Assembly and Binning Statistics
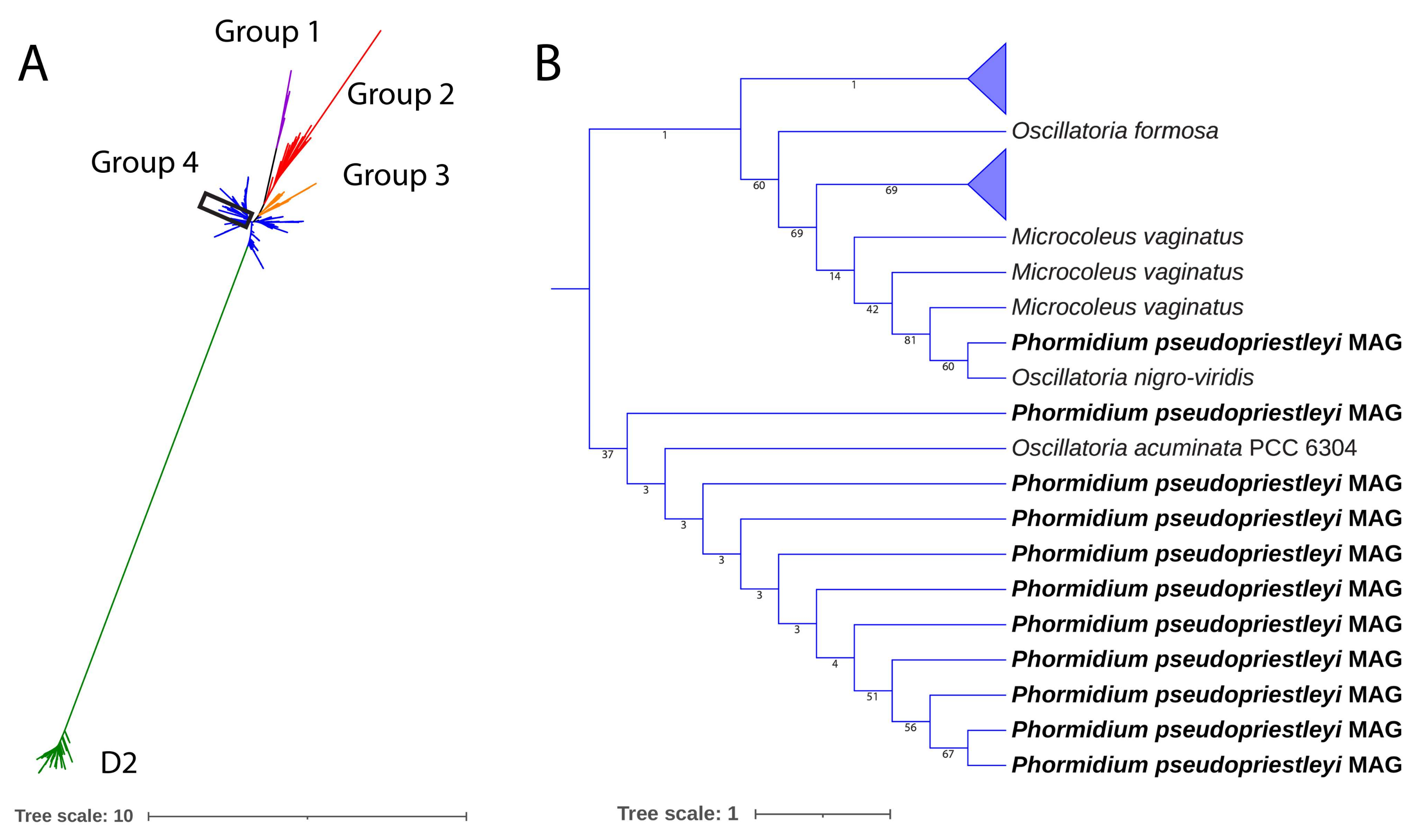
3.3. Taxonomic Assignment of MAG
3.4. Photosynthetic and Electron Transport Machinery
3.5. Metabolic Pathways
3.6. Genes Implicated in the Adaption to Environmental Stress
4. Discussion
4.1. Sulfide Resistance in P. pseudopriestleyi
4.2. Ecology of P. pseudopriestleyi in Lake Fryxell
4.3. P. pseudopriestleyi in Other Environments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasting, J.F. What Caused the Rise of Atmospheric O2? Chem. Geol. 2013, 362, 13–25. [Google Scholar] [CrossRef]
- Canfield, D.E. A New Model for Proterozoic Ocean Chemistry. Nature 1998, 396, 450–453. [Google Scholar] [CrossRef]
- Parnell, J.; Adrian, J.B. Microbial Sulphate Reduction during Neoproterozoic Glaciation, Port Askaig Formation, UK. J. Geol. Soc. 2017, 174, 850–854. [Google Scholar] [CrossRef]
- Dahl, T.W.; Canfield, D.E.; Rosing, M.T.; Frei, R.E.; Gordon, G.W.; Knoll, A.H.; Anbar, A.D. Molybdenum Evidence for Expansive Sulfidic Water Masses in ~750Ma Oceans. Earth Planet. Sci. Lett. 2011, 311, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Algeo, T.J.; Zhou, Q.; Yu, W.; Du, Y.; Qin, Y.; Xu, Y.; Yuan, L.; Pan, W. Large Accumulations of 34S-Enriched Pyrite in a Low-Sulfate Marine Basin: The Sturtian Nanhua Basin, South China. Precambrian Res. 2019, 335. [Google Scholar] [CrossRef]
- Voorhies, A.A.; Biddanda, B.A.; Kendall, S.T.; Jain, S.; Marcus, D.N.; Nold, S.C.; Sheldon, N.D.; Dick, G.J. Cyanobacterial Life at Low O2: Community Genomics and Function Reveal Metabolic Versatility and Extremely Low Diversity in a Great Lakes Sinkhole Mat. Geobiology 2012, 10, 250–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesada, A.; Vincent, W.F. Cyanobacteria in the Cryosphere: Snow, Ice and Extreme Cold. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 387–399. [Google Scholar] [CrossRef]
- Klatt, J.M.; Meyer, S.; Häusler, S.; Macalady, J.L.; de Beer, D.; Polerecky, L. Structure and Function of Natural Sulphide-Oxidizing Microbial Mats under Dynamic Input of Light and Chemical Energy. ISME J. 2016, 10, 921–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macalady, J.L.; Lyon, E.H.; Koffman, B.; Albertson, L.K.; Meyer, K.; Galdenzi, S.; Mariani, S. Dominant Microbial Populations in Limestone-Corroding Stream Biofilms, Frasassi Cave System, Italy. Appl. Environ. Microbiol. 2006, 72, 5596–5609. [Google Scholar] [CrossRef] [Green Version]
- De Beer, D.; Weber, M.; Chennu, A.; Hamilton, T.; Lott, C.; Macalady, J.; Klatt, J.M. Oxygenic and Anoxygenic Photosynthesis in a Microbial Mat from an Anoxic and Sulfidic Spring. Environ. Microbiol. 2017, 19, 1251–1265. [Google Scholar] [CrossRef]
- Hamilton, T.L.; Klatt, J.M.; de Beer, D.; Macalady, J.L. Cyanobacterial Photosynthesis under Sulfidic Conditions: Insights from the Isolate Leptolyngbya Sp. Strain Hensonii. ISME J. 2018, 12, 568–584. [Google Scholar] [CrossRef] [Green Version]
- Sumner, D.Y.; Hawes, I.; Mackey, T.J.; Jungblut, A.D.; Doran, P.T. Antarctic Microbial Mats: A Modern Analog for Archean Lacustrine Oxygen Oases. Geology 2015, 43, 887–890. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Hawes, I.; Mackey, T.J.; Krusor, M.; Doran, P.T.; Sumner, D.Y.; Eisen, J.A.; Hillman, C.; Goroncy, A.K. Microbial Mat Communities along an Oxygen Gradient in a Perennially Ice-Covered Antarctic Lake. Appl. Environ. Microbiol. 2016, 82, 620–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A.; Padan, E.; Malkin, S. Sulfide Inhibition of Photosystem II in Cyanobacteria (Blue-Green Algae) and Tobacco Chloroplasts. Biochim. Biophys. Acta 1979, 546, 270–279. [Google Scholar] [CrossRef]
- Miller, S.R.; Bebout, B.M. Variation in Sulfide Tolerance of Photosystem II in Phylogenetically Diverse Cyanobacteria from Sulfidic Habitats. Appl. Environ. Microbiol. 2004, 70, 736–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Y.; Jørgensen, B.B.; Revsbech, N.P.; Poplawski, R. Adaptation to Hydrogen Sulfide of Oxygenic and Anoxygenic Photosynthesis among Cyanobacteria. Appl. Environ. Microbiol. 1986, 51, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Dick, G.J.; Grim, S.L.; Klatt, J.M. Controls on O2 Production in Cyanobacterial Mats and Implications for Earth’s Oxygenation. Annu. Rev. Earth Planet. Sci. 2018, 46. [Google Scholar] [CrossRef]
- Klatt, J.M.; Haas, S.; Yilmaz, P.; de Beer, D.; Polerecky, L. Hydrogen Sulfide Can Inhibit and Enhance Oxygenic Photosynthesis in a Cyanobacterium from Sulfidic Springs. Environ. Microbiol. 2015, 17, 3301–3313. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Toward Understanding the Origin and Evolution of Cellular Organisms. Protein Sci. Publ. Protein Soc. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Bishop, N.I. The Influence of the Herbicide, DCMU, on the Oxygen-Evolving System of Photosynthesis. Biochim. Biophys. Acta 1958, 27, 205–206. [Google Scholar] [CrossRef]
- Mulo, P.; Sicora, C.; Aro, E. Cyanobacterial PsbA Gene Family: Optimization of Oxygenic Photosynthesis. Cell. Mol. Life Sci. 2009, 66, 3697. [Google Scholar] [CrossRef] [Green Version]
- Mulo, P.; Sakurai, I.; Aro, E. Strategies for PsbA Gene Expression in Cyanobacteria, Green Algae and Higher Plants: From Transcription to PSII Repair. Biochim. Biophys. Acta 2012, 1817, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Summerfield, T.C.; Toepel, J.; Sherman, L.A. Low-Oxygen Induction of Normally Cryptic PsbA Genes in Cyanobacteria. Biochemistry 2008, 47, 12939–12941. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T.; Murray, J.W.; Rutherford, A.W. Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria. Mol. Biol. Evol. 2015, 32, 1310–1328. [Google Scholar] [CrossRef]
- Bronstein, M.; Schütz, M.; Hauska, G.; Padan, E.; Shahak, Y. Cyanobacterial Sulfide-Quinone Reductase: Cloning and Heterologous Expression. J. Bacteriol. 2000, 182, 3336–3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlick, S.; Oren, A.; Padan, E. Occurrence of Facultative Anoxygenic Photosynthesis among Filamentous and Unicellular Cyanobacteria. J. Bacteriol. 1997, 129, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Y.; Padan, E.; Shilo, M. Facultative Anoxygenic Photosynthesis in the Cyanobacterium Oscillatoria Limnetica. J. Bacteriol. 1975, 123, 855–861. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Y.; Jørgensen, B.B.; Padan, E.; Shilo, M. Sulphide-Dependent Anoxygenic Photosynthesis in the Cyanobacterium Oscillatoria Limnetica. Nature 1975, 257, 489. [Google Scholar] [CrossRef]
- Green, W.J.; Lyons, W.B. The Saline Lakes of the McMurdo Dry Valleys, Antarctica. Aquat. Geochem. 2009, 15, 321–348. [Google Scholar] [CrossRef]
- McKnight, D.M.; Niyogi, D.K.; Alger, A.S.; Bomblies, A.; Conovitz, P.A.; Tate, C.M. Dry Valley Streams in Antarctica: Ecosystems Waiting for Water. BioScience 1999, 49, 985–995. [Google Scholar] [CrossRef]
- Lawrence, M.J.F.; Hendy, C.H. Water Column and Sediment Characteristics of Lake Fryxell, Taylor Valley, Antarctica. N. Z. J. Geol. Geophys. 1985, 28, 543–552. [Google Scholar] [CrossRef]
- Vincent, W.F. Production Strategies in Antarctic Inland Waters: Phytoplankton Eco-Physiology in a Permanently Ice-Covered Lake. Ecology 1981, 62, 1215–1224. [Google Scholar] [CrossRef]
- Lyons, W.B.; Frape, S.K.; Welch, K.A. History of McMurdo Dry Valley Lakes, Antarctica, from Stable Chlorine Isotope Data. Geology 1999, 27, 527–530. [Google Scholar] [CrossRef]
- Roberts, E.C.; Laybourn-Parry, J.; McKnight, D.M.; Novarino, G. Stratification and Dynamics of Microbial Loop Communities in Lake Fryxell, Antarctica. Freshw. Biol. 2000, 44, 649–661. [Google Scholar] [CrossRef]
- Laybourn-Parry, J.; James, M.R.; McKnight, D.M.; Priscu, J.; Spaulding, S.A.; Shiel, R. The Microbial Plankton of Lake Fryxell, Southern Victoria Land, Antarctica during the Summers of 1992 and 1994. Polar Biol. 1997, 17, 54–61. [Google Scholar] [CrossRef]
- Spigel, R.H.; Priscu, J.C. Physical Limnology of the McMurdo Dry Valleys Lakes. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica; American Geophysical Union (AGU): Washington, DC, USA, 2013; pp. 153–187. [Google Scholar] [CrossRef]
- Howard-Williams, C.; Schwarz, A.; Hawes, I.; Priscu, J.C. Optical Properties of the McMurdo Dry Valley Lakes, Antarctica. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica; American Geophysical Union (AGU): Washington, DC, USA, 2013; pp. 189–203. [Google Scholar] [CrossRef]
- Lizotte, M.P.; Priscu, J.C. Photosynthesis-Irradiance Relationships in Phytoplankton from the Physically Stable Water Column of a Perennially Ice-Covered Lake (Lake Bonney, Antarctica). J. Phycol. 1992, 28, 179–185. [Google Scholar] [CrossRef]
- Miner, G. Standard Methods for the Examination of Water and Wastewater, 21st Edition. Am. Water Works Assoc. J. 2006, 98, 130. [Google Scholar] [CrossRef]
- Hillman, C. Structure of Benthic Microbial Mat Assemblages in Lake Fryxell, Antarctica. 2013. Available online: https://ir.canterbury.ac.nz/handle/10092/8737 (accessed on 28 November 2018).
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A Fast and Accurate Illumina Paired-End ReAd MergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crusoe, M.R.; Alameldin, H.F.; Awad, S.; Boucher, E.; Caldwell, A.; Cartwright, R.; Charbonneau, A.; Constantinides, B.; Edvenson, G.; Fay, S.; et al. The Khmer Software Package: Enabling Efficient Nucleotide Sequence Analysis. F1000Research 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, C.; Luo, R.; Sadakane, K.; Lam, T. MEGAHIT: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly via Succinct de Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. Available online: http://arxiv.org/abs/1303.3997 (accessed on 20 May 2019).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eren, A.M.; Esen, Ö.C.; Quince, C.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L.; Delmont, T.O. Anvi’o: An Advanced Analysis and Visualization Platform for ‘omics Data. PeerJ 2015, 3. [Google Scholar] [CrossRef]
- Delmont, T.O.; Eren, A.M. Linking Pangenomes and Metagenomes: The Prochlorococcus Metapangenome. PeerJ 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Chaumeil, P.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A Toolkit to Classify Genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic Gene Recognition and Translation Initiation Site Identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7. [Google Scholar] [CrossRef] [Green Version]
- Matsen, F.A.; Kodner, R.B.; Armbrust, E.V. Pplacer: Linear Time Maximum-Likelihood and Bayesian Phylogenetic Placement of Sequences onto a Fixed Reference Tree. BMC Bioinform. 2010, 11, 538. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast Genome and Metagenome Distance Estimation Using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, J.; Holder, M.T. DendroPy: A Python Library for Phylogenetic Computing. Bioinformatics 2010, 26, 1569–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array Programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. Comput. Resour. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Arkin, A.P.; Cottingham, R.W.; Henry, C.W.; Harris, N.L.; Stevens, R.L.; Maslov, S.; Dehal, P.; Ware, D.; Perez, F.; Canon, S.; et al. KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.T.; Moritz, D.; O’Brien, M.P.; Reidl, F.; Reiter, T.; Sullivan, B.D. Exploring Neighborhoods in Large Metagenome Assembly Graphs Using Spacegraphcats Reveals Hidden Sequence Diversity. Genome Biol. 2020, 21, 164. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Shahak, Y.; Hauska, H. Advances in Photosynthesis and Respiration. In Sulfur Metabolism in Phototrophic Organisms, Sulfide Oxidation from Cyanobacteria to Humans: Sulfide–Quinone Oxidoreductase (SQR); Springer: Dordrecht, The Netherlands, 2008; pp. 319–335. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldónm, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Chrismas, N.A.M.; Williamson, C.J.; Yallop, M.L.; Anesio, A.M.; Sánchez-Baracaldo, P. Photoecology of the Antarctic Cyanobacterium Leptolyngbya sp. BC1307 Brought to Light through Community Analysis, Comparative Genomics and in Vitro Photosphysiology. Mol. Ecol. 2018, 27, 5279–5293. [Google Scholar] [CrossRef]
- Grettenberger, C.L.; Sumner, D.Y.; Wall, K.C.; Brown, C.T.; Eisen, J.A.; Mackey, T.J.; Hawes, I.; Jospin, G.; Jungblut, A.D. A Phylogenetically Novel Cyanobacterium Most Closely Related to Gloeobacter. ISME J. 2020, 14, 2142–2152. [Google Scholar] [CrossRef]
- Tang, J.; Du, L.-M.; Lian, Y.-M.; Daroch, M. Complete Genome Sequence and Comparative Analysis of Synechococcus sp. CS-601 (SynAce01), a Cold-Adapted Cyanobacterium from an Oligotrophic Antarctic Habitat. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Lara, Y.; Durieu, B.; Cornet, L.; Verlaine, O.; Rippka, R.; Pessi, I.S.; Misztak, A.; Joris, B.; Javaux, E.J.; Baurain, D.; et al. Draft Genome Sequence of the Axenic Strain Phormidesmis priestleyi ULC007, a Cyanobacterium Isolated from Lake Bruehwiler (Larsemann Hills). Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Chrismas, N.A.M.; Barker, G.; Anesio, A.M.; Sánchez-Baracaldo, P. Genomic Mechanisms for Cold Tolerance and Production of Exopolysaccharides in the Arctic Cyanobacterium Phormidesmis Priestleyi BC1401. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; de Marsac, N.T.; Rippka, R.; et al. Improving the Coverage of the Cyanobacterial Phylum Using Diversity-Driven Genome Sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The Enveomics Collection: A Toolbox for Specialized Analyses of Microbial Genomes and Metagenomes. PeerJ Inc. 2016. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within Cyanobacterial Mat Communities in Variable Salinity Meltwater Ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Taton, A.; Grubisic, S.; Balthasart, P.; Hodgson, D.A.; Laybourn-Parry, J.; Wilmotte, A. Biogeographical Distribution and Ecological Ranges of Benthic Cyanobacteria in East Antarctic Lakes. FEMS Microbiol. Ecol. 2006, 57, 272–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barco, R.A.; Garrity, G.M.; Scott, J.J.; Amend, J.P.; Nealson, K.H.; Emerson, D. A Genus Definition for Bacteria and Archaea Based on a Standard Genome Relatedness Index. MBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Bogorad, L. Phycobiliproteins and Complementary Chromatic Adaptation. Annu. Rev. Plant Physiol. 1975, 26, 369–401. [Google Scholar] [CrossRef]
- Durán, R.V.; Hervás, M.; de la Rosa, M.; Navarro, J.A. The Efficient Functioning of Photosynthesis and Respiration in Synechocystis sp. PCC 6803 Strictly Requires the Presence of Either Cytochrome c6 or Plastocyanin. J. Biol. Chem. 2004, 279, 7229–7233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, C.I.; Vass, I.; Rákhely, G.; Vass, I.Z.; Tóth, A.; Duzs, A.; Peca, L.; Kruk, J.; Kós, P.B. Coregulated Genes Link Sulfide:Quinone Oxidoreductase and Arsenic Metabolism in Synechocystis Sp. Strain PCC6803. J. Bacteriol. 2014, 196, 3430–3440. [Google Scholar] [CrossRef] [Green Version]
- Lea-Smith, D.J.; Bombelli, P.; Vasudevan, R.; Howe, C.J. Photosynthetic, Respiratory and Extracellular Electron Transport Pathways in Cyanobacteria. Biochim. Biophys. Acta 2016, 1857, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, C.W. Electron Transport and Light-Harvesting Switches in Cyanobacteria. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.W. Sequence Variation at the Oxygen-Evolving Centre of Photosystem II: A New Class of ‘Rogue’ Cyanobacterial D1 Proteins. Photosynth. Res. 2012, 110, 177–184. [Google Scholar] [CrossRef]
- Vermaas, W.; Charité, J.; Shen, G.Z. Glu-69 of the D2 Protein in Photosystem II Is a Potential Ligand to Mn Involved in Photosynthetic Oxygen Evolution. Biochemistry 1990, 29, 5325–5332. [Google Scholar] [CrossRef]
- Varin, T.; Lovejoy, C.; Jungblut, A.D.; Vincent, W.F.; Corbeil, J. Metagenomic Profiling of Arctic Microbial Mat Communities as Nutrient Scavenging and Recycling Systems. Limnol. Oceanogr. 2010, 55, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Dillon, M.L.; Hawes, I.; Jungblut, A.D.; Mackey, T.J.; Eisen, J.A.; Doran, P.T.; Sumner, D.Y. Energetic and Environmental Constraints on the Community Structure of Benthic Microbial Mats in Lake Fryxell, Antarctica. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, A.R.; Grein, F.; Oliveira, G.P.; Venceslau, S.S.; Keller, K.L.; Wall, J.D.; Pereira, I.A.C. The FlxABCD-HdrABC Proteins Correspond to a Novel NADH Dehydrogenase/Heterodisulfide Reductase Widespread in Anaerobic Bacteria and Involved in Ethanol Metabolism in Desulfovibrio Vulgaris Hildenborough. Environ. Microbiol. 2015, 17, 2288–2305. [Google Scholar] [CrossRef] [PubMed]
- Hincha, D.K.; Hagemann, M. Stabilization of Model Membranes during Drying by Compatible Solutes Involved in the Stress Tolerance of Plants and Microorganisms. Biochem. J. 2004, 383, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Hershkovitz, N.; Oren, A.; Cohen, Y. Accumulation of Trehalose and Sucrose in Cyanobacteria Exposed to Matric Water Stress. Appl. Environ. Microbiol. 1991, 57, 645–648. [Google Scholar] [CrossRef] [Green Version]
- Page-Sharp, M.; Behm, C.A.; Smith, G.D. Involvement of the Compatible Solutes Trehalose and Sucrose in the Response to Salt Stress of a Cyanobacterial Scytonema Species Isolated from Desert Soils. Biochim. Biophys. Acta 1999, 1472, 519–528. [Google Scholar] [CrossRef]
- Potts, M.; Slaughter, S.M.; Hunneke, F.U.; Garst, J.F.; Helm, R.F. Desiccation Tolerance of Prokaryotes: Application of Principles to Human Cells. Integr. Comp. Biol. 2005, 45, 800–809. [Google Scholar] [CrossRef] [Green Version]
- Hagemann, M. Molecular Biology of Cyanobacterial Salt Acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, F.; Klähn, S.; Hagemann, M. Salt-Regulated Accumulation of the Compatible Solutes Sucrose and Glucosylglycerol in Cyanobacteria and Its Biotechnological Potential. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Scherer, S. UV Protection in Cyanobacteria. Eur. J. Phycol. 1999, 34, 329–338. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Multiple Roles of Photosynthetic and Sunscreen Pigments in Cyanobacteria Focusing on the Oxidative Stress. Metabolites 2013, 3, 463–483. [Google Scholar] [CrossRef]
- Sorrels, C.M.; Proteau, P.J.; Gerwick, W.H. Organization, Evolution, and Expression Analysis of the Biosynthetic Gene Cluster for Scytonemin, a Cyanobacterial UV-Absorbing Pigment. Appl. Environ. Microbiol. 2009, 75, 4861–4869. [Google Scholar] [CrossRef] [Green Version]
- Mills, L.A.; McCormick, A.J.; Lea-Smith, D.J. Current Knowledge and Recent Advances in Understanding Metabolism of the Model Cyanobacterium Synechocystis sp. PCC 6803. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [Green Version]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar]
- Zhu, Y.; Graham, J.E.; Ludwig, M.; Xiong, W.; Alvey, R.M.; Shen, G.; Bryant, D.A. Roles of Xanthophyll Carotenoids in Protection against Photoinhibition and Oxidative Stress in the Cyanobacterium Synechococcus Sp. Strain PCC 7002. Arch. Biochem. Biophys. 2010, 504, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, L.; Brejnrod, A.; Schliep, M.; Sørensen, S.J.; Larkum, A.W.D.; Kühl, M. Chlorophyll f -Driven Photosynthesis in a Cavernous Cyanobacterium. ISME J. 2015, 9, 2108–2111. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, D.F.; Tiedje, J.M. Coping with Our Cold Planet. Appl. Environ. Microbiol. 2008, 74, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watzer, B.; Forchhammer, K. Cyanophycin Synthesis Optimizes Nitrogen Utilization in the Unicellular Cyanobacterium Synechocystis Sp. Strain PCC 6803. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Klatt, J.M.; Al-Najjar, M.A.A.; Yilmaz, P.; Lavik, G.; de Beer, D.; Polerecky, L. Anoxygenic Photosynthesis Controls Oxygenic Photosynthesis in a Cyanobacterium from a Sulfidic Spring. Appl. Environ. Microbiol. 2015, 81, 2025–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard-Williams, C.; Pridmore, R.; Downes, M.T.; Vincent, W.F. Microbial Biomass, Photosynthesis and Chlorophyll a Related Pigments in the Ponds of the McMurdo Ice Shelf, Antarctica. Antarct. Sci. 1989, 1, 125–131. [Google Scholar] [CrossRef]
- Priscu, J.C. Phytoplankton Nutrient Deficiency in Lakes of the McMurdo Dry Valleys, Antarctica. Freshw. Biol. 1995, 34, 215–227. [Google Scholar] [CrossRef]
- Roos, J.C.; Vincent, W.F. Temperature Dependence of UV Radiation Effects on Antarctic Cyanobacteria. J. Phycol. 1998, 34, 118–125. [Google Scholar] [CrossRef]
- Keys, J.R.; Williams, K. Origin of Crystalline, Cold Desert Salts in the McMurdo Region, Antarctica. Geochim. Cosmochim. Acta 1981, 2299–2309. [Google Scholar] [CrossRef]
- De Mora, S.J.; Whitehead, R.F.; Gregory, M. The Chemical Composition of Glacial Melt Water Ponds and Streams on the McMurdo Ice Shelf, Antarctica. Antarct. Sci. 1994, 6, 17–27. [Google Scholar] [CrossRef]
- Hodgson, D.A.; Verleyen, E.; Sabbe, K.; Squier, A.H.; Keely, B.J.; Leng, M.J.; Saunders, K.M.; Vyverman, W. Late Quaternary Climate-Driven Environmental Change in the Larsemann Hills, East Antarctica, Multi-Proxy Evidence from a Lake Sediment Core. Quat. Res. 2005, 64, 83–99. [Google Scholar] [CrossRef]
- Hawes, I.; Jungblut, A.D.; Matys, E.D.; Summons, R.E. The ‘Dirty Ice’ of the McMurdo Ice Shelf: Analogues for Biological Oases during the Cryogenian. Geobiology 2018, 16, 369–377. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Depth (m) | Total Sulfide (mg L−1) |
|---|---|
| 8 | <0.01 |
| 9 | <0.01 |
| 9.5 | <0.01 |
| 9.8 | 0.091 |
| 10.1 | 0.571 |
| 10.3 | 0.885 |
| 10.7 | 2.238 |
| Metric | Coassembly |
|---|---|
| Total number of contigs | 137,226 |
| Longest contig length | 453,833 |
| Total length (bp) | 175,382,959 |
| GC content (%) | 61.61 |
| N50 | 1441 |
| Reads mapped from culture (%) | 79.80 |
| Reads mapped from mat (%) | 54.27 |
| Total reads mapped (%) | 77.85 |
| Number of contigs ≥ 0 bp | 336,522 |
| Number of contigs ≥ 1000 bp | 41,881 |
| Number of contigs ≥ 5000 bp | 2359 |
| Number of contigs ≥ 10.000 bp | 814 |
| Number of contigs ≥ 25.000 bp | 303 |
| Number of contigs ≥ 50.000 bp | 152 |
| Metric | MAG |
|---|---|
| Total number of contigs | 678 |
| Longest contig length | 44,245 |
| Total length (bp) | 5,965,908 |
| GC content (%) | 47.43 |
| N50 | 10,908 |
| Completion (%) | 91.73 |
| Contamination (%) | 1.35 |
| Number of protein coding genes | 4738 |
| Number of contigs ≥ 0 bp | 678 |
| Number of contigs ≥ 1000 bp | 678 |
| Number of contigs ≥ 5000 bp | 458 |
| Number of contigs ≥ 10.000 bp | 203 |
| Number of contigs ≥ 25.000 bp | 20 |
| Number of contigs ≥ 50.000 bp | 0 |
| Complex | Gene | Presence | Function |
|---|---|---|---|
| Allophycocyanin | apcA | Yes | Allophycocyanin α subunit |
| apcB | Yes | Allophycocyanin β subunit | |
| apcC | Yes | Phycobilisome core linker protein | |
| apcD | Yes | Allophycocyanin-B | |
| apcE | Yes | Phycobilisome core-membrne linker protein | |
| apcF | Yes | Phycobilisome core component | |
Phycocyanin/ Phycoerythrocyanin | cpcA | Yes | Phycocyanin α chain |
| cpcB | Yes | Phycocyanin β chain | |
| cpcC | Yes | Phycocyanin-associated rod linker protein | |
| cpcD | No | Phycocyanin-associated, rod | |
| cpcE | Yes | Phycocyanobilin lyase α subunit | |
| cpcF | Yes | Phycocyanobilin lyase β subunit | |
| cpcG | Yes | Phycobilisome rod-core linker protein | |
| Phycoerythrin | cpeA | No | Phycoerythrin α chain |
| cpeB | No | Phycoerythrin β chain | |
| cpeC | No | Phycoerythrin-associated linker protein | |
| cpeD | No | Phycoerythrin-associated linker protein | |
| cpeE | No | Phycoerythrin-associated linker protein | |
| cpeR | No | Phycoerythrin-associated linker protein | |
| cpeS | No | Phycoerythrin-associated linker protein | |
| cpeT | No | CpeT protein | |
| cpeU | No | Billin biosynthesis protein | |
| cpeY | No | Billin biosynthesis protein | |
| cpeZ | No | Billin biosynthesis protein | |
| Photosystem II | psbA | Yes | Photosystem II P680 reaction center D1 protein |
| psbD | Yes | Photosystem II P680 reaction center D2 protein | |
| psbC | Yes | Photosystem II CP43 chlorophyll apoprotein | |
| psbB | Yes | Photosystem II CP47 chlorophyll apoprotein | |
| psbE | Yes | Photosystem II cytochrome b559 subunit α | |
| psbF | Yes | Photosystem II cytochrome b559 subunit β | |
| psbL | Yes | Photosystem II PsbL protein | |
| psbJ | Yes | Photosystem II PsbJ protein | |
| psbK | Yes | Photosystem II PsbK protein | |
| pskM | Yes | Photosystem II PsbM protein | |
| psbH | Yes | Photosystem II PsbH protein | |
| psbI | Yes | Photosystem II PsbI protein | |
| psbO | Yes | Photosystem II oxygen-evolving enhancer protein 1 | |
| psbP | Yes | Photosystem II oxygen-evolving enhancer protein 2 | |
| psbQ | No | Photosystem II oxygen-evolving enhancer protein 3 | |
| psbR | No | Photosystem II 10 kDa protein | |
| psbS | No | Photosystem II 22kDa protein | |
| psbT | Yes | Photosystem II PsbT protein | |
| psbU | Yes | Photosystem II PsbU protein | |
| psbV | Yes | Photosystem II cytochrome c550 | |
| psbW | No | Photosystem II PsbW protein | |
| psbX | Yes | Photosystem II PsbX protein | |
| psbY | Yes | Photosystem II PsbY protein | |
| psbZ | Yes | Photosystem II PsbZ protein | |
| Psb27 | Yes | Photosystem II Psb27 protein | |
| psb28 | Yes | Photosystem II 13kDa protein | |
| psb28-2 | No | Photosystem II Psb28-2 protein | |
| Photosystem I | psaA | Yes | Photosystem I P700 chlorophyll a apoprotein A1 |
| psaB | Yes | Photosystem I P700 chlorophyll a apoprotein A2 | |
| psaC | Yes | Photosystem I subunit VII | |
| psaD | Yes | Photosystem I subunit II | |
| psaE | Yes | Photosystem I subunit IV | |
| psaF | Yes | Photosystem I subunit III | |
| psaG | No | Photosystem I subunit V | |
| psaH | No | Photosystem I subunit VI | |
| psaI | Yes | Photosystem I subunit VIII | |
| psaJ | No | Photosystem I subunit IX | |
| psaK | Yes | Photosystem I subunit X | |
| psaL | Yes | Photosystem I subunit XI | |
| psaM | Yes | Photosystem I subunit XII | |
| psaN | No | Photosystem I subunit PsaN | |
| psaO | No | Photosystem I subunit PsaO | |
| psaX | No | Photosystem I 4.8kDa protein | |
Cytochrome b6f Complex | petB | Yes | cytochrome b6 |
| petD | Yes | cytochrome b6f complex subunit 4 | |
| petA | Yes | apocytochrome f | |
| petC | Yes | cytochrome b6f complex iron-sulfur subunit | |
| petL | No | cytochrome b6f complex subunit 6 | |
| petM | No | cytochrome b6f subunit 7 | |
| petN | No | cytochrome b6f complex subunit 8 | |
| petG | No | cytochrome b6f complex subunit 5 | |
| Photosynthetic Electron Transport Chain | petE | No | plastocyanin |
| petF | Yes | ferredoxin | |
| petH | Yes | ferredoxin-NADP+ reductase | |
| petJ | Yes | cytochrome c6 | |
| F-type ATPase | atpD | Yes | H+/Na+ transporting ATPase subunit β |
| atpA | Yes | F-type H+/Na+ transporting ATPase subunit α | |
| atpG | Yes | H+ transporting ATPase subunit γ | |
| atpH | Yes | F-type H+ transporting ATPase subunit δ | |
| atpC | Yes | F-type H+ transporting ATPase subunit ε | |
| atpE | Yes | F-type H+ transporting ATPase subunit c | |
| atpB | Yes | F-type H+ transporting ATPase subunit a | |
| atpF | Yes | F-type H+ transporting ATPase subunit b | |
NADH Dehydrogenase | ndhC | Yes | NADH-quinone oxidoreductase subunit 3 |
| ndhK | Yes | NADH-quinone oxidoreductase subunit K | |
| ndhJ | Yes | NADH-quinone oxidoreductase subunit J | |
| ndhH | Yes | NADH-quinone oxidoreductase subunit H | |
| ndhA | Yes | NADH-quinone oxidoreductase subunit 1 | |
| ndhI | Yes | NADH-quinone oxidoreductase subunit I | |
| ndhG | Yes | NADH-quinone oxidoreductase subunit 6 | |
| ndhE | Yes | NADH-quinone oxidoreductase subunit 4L | |
| ndhF | Yes | NADH-quinone oxidoreductase subunit 5 | |
| ndhD | Yes | NADH-quinone oxidoreductase subunit 4 | |
| ndhB | Yes | NADH-quinone oxidoreductase subunit 2 | |
| ndhL | Yes | NADH-quinone oxidoreductase subunit L | |
| ndhM | Yes | NADH-quinone oxidoreductase subunit M | |
| ndhN | No | NADH-quinone oxidoreductase subunit N | |
| hoxE | Yes | bidirectional [NiFe] hydroganse diaphorase subunit | |
| hoxF | Yes | bidirectional [NiFe] hydroganse diaphorase subunit | |
| hoxU | Yes | bidirectional [NiFe] hydroganse diaphorase subunit | |
| Succinate Dehydrogenase | sdhC | Yes | H+/Na+ transporting ATPase subunit β |
| sdhD | No | F-type H+/Na+ transporting ATPase subunit α | |
| sdhA | Yes | H+ transporting ATPase subunit γ | |
| sdhB | Yes | F-type H+ transporting ATPase subunit δ | |
| Cytochrome c oxidase | ctaC | Yes | cytochrome c oxidase subunit 2 |
| ctaD | Yes | cytochrome c oxidase subunit 1 | |
| ctaE | Yes | cytochrome c oxidase subunit 3 | |
| ctaF | No | cytochrome c oxidase subunit 4 | |
| Cytochrome bd complex | cydA | Yes | cytochrome bd ubiquinol oxidase subunit I |
| cydB | Yes | cytochrome bd ubiquinol oxidase subunit II | |
| cydX | No | cytochrome bd ubiquinol oxidase subunit X |
| Category | Complex or System | Number of Genes in MAG | Total Number of Genes in KEGG Category |
|---|---|---|---|
| Phycobilisome Antenna Proteins | Allophycocyanin | 6 | 6 |
| Phycocyanin/Phycoerythrin | 6 | 7 | |
| Phycoerythrin | 0 | 11 | |
| Photosynthesis Machinery | Photosystem II | 22 | 27 |
| Photosystem I | 10 | 16 | |
| Cytochrome b6f complex | 4 | 8 | |
| Photosynthetic electron transport | 3 | 4 | |
| F-type ATPase | 8 | 8 | |
Nitrogen Metabolism | Dissimilatory Nitrate Reduction | 0 | 4 |
| Assimilatory Nitrate Reduction | 2 | 5 | |
| All Nitrogen Metabolism | 9 | 35 | |
| Sulfur Metabolism | Assimilatory Sulfate Reduction | 4 | 7 |
| Dissimilatory Sulfate Reduction and Oxidation | 1 | 3 | |
| All Sulfur Metabolism | 10 | 54 | |
| Carbon Fixation | Carbon Fixation in Photosynthetic Organisms | 9 | 23 |
| Methane Metabolism | Methane Metabolism | 19 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lumian, J.E.; Jungblut, A.D.; Dillion, M.L.; Hawes, I.; Doran, P.T.; Mackey, T.J.; Dick, G.J.; Grettenberger, C.L.; Sumner, D.Y. Metabolic Capacity of the Antarctic Cyanobacterium Phormidium pseudopriestleyi That Sustains Oxygenic Photosynthesis in the Presence of Hydrogen Sulfide. Genes 2021, 12, 426. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030426
Lumian JE, Jungblut AD, Dillion ML, Hawes I, Doran PT, Mackey TJ, Dick GJ, Grettenberger CL, Sumner DY. Metabolic Capacity of the Antarctic Cyanobacterium Phormidium pseudopriestleyi That Sustains Oxygenic Photosynthesis in the Presence of Hydrogen Sulfide. Genes. 2021; 12(3):426. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030426
Chicago/Turabian StyleLumian, Jessica E., Anne D. Jungblut, Megan L. Dillion, Ian Hawes, Peter T. Doran, Tyler J. Mackey, Gregory J. Dick, Christen L. Grettenberger, and Dawn Y. Sumner. 2021. "Metabolic Capacity of the Antarctic Cyanobacterium Phormidium pseudopriestleyi That Sustains Oxygenic Photosynthesis in the Presence of Hydrogen Sulfide" Genes 12, no. 3: 426. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030426