
Noninvasive Prenatal Paternity Testing with a Combination of Well-Established SNP and STR Markers Using Massively Parallel Sequencing

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Library Preparation and Massively Parallel Sequencing
2.3. Pedigree Confirmation by CE-Based STR Typing
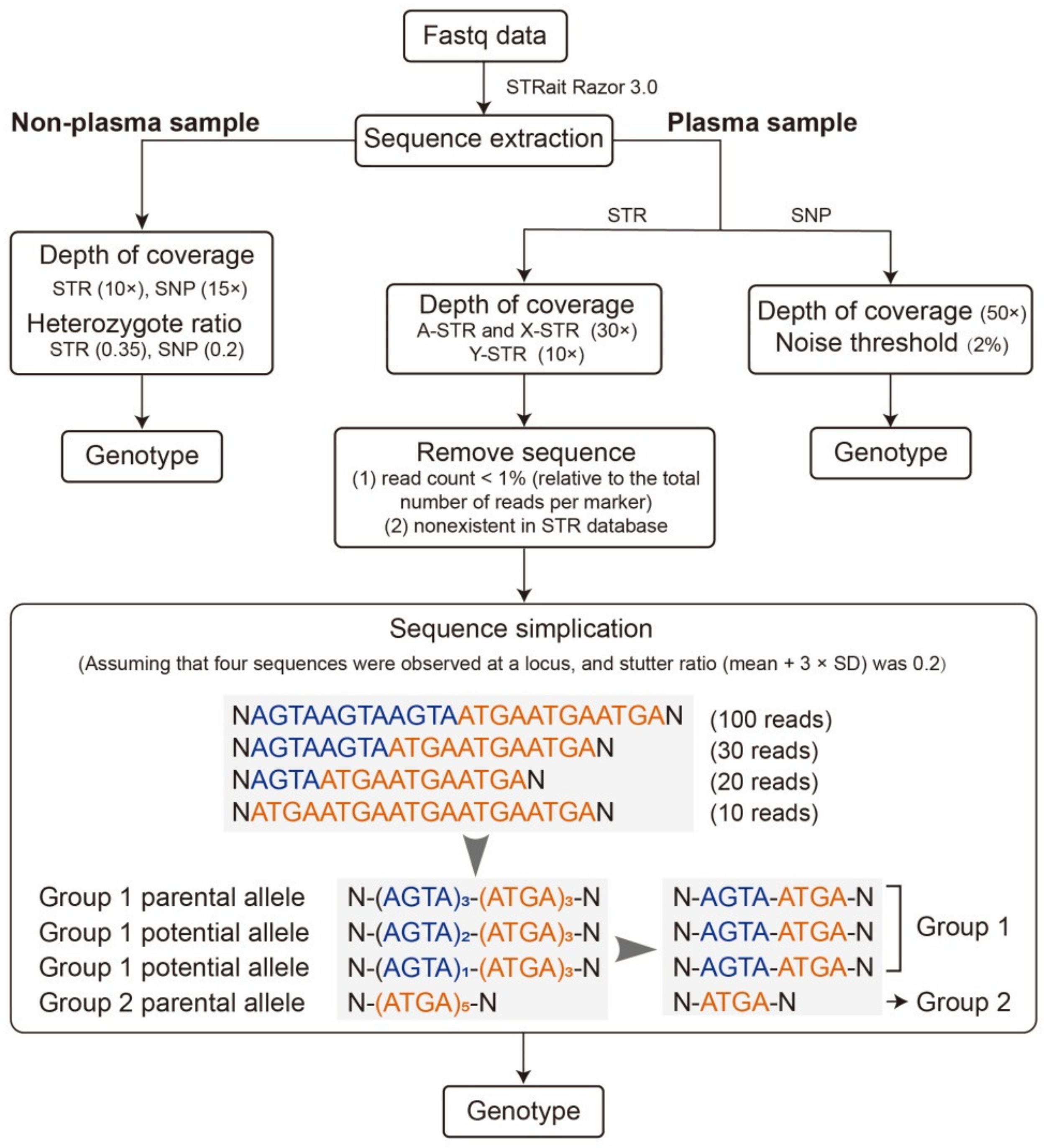
2.4. Genotype Calling
2.5. Data Analysis
2.6. PI and CPI Calculation for NIPPT
3. Results
3.1. Performance of the ForenSeq DNA Signature Prep Kit
3.2. PIA Identification from Plasma for NIPPT
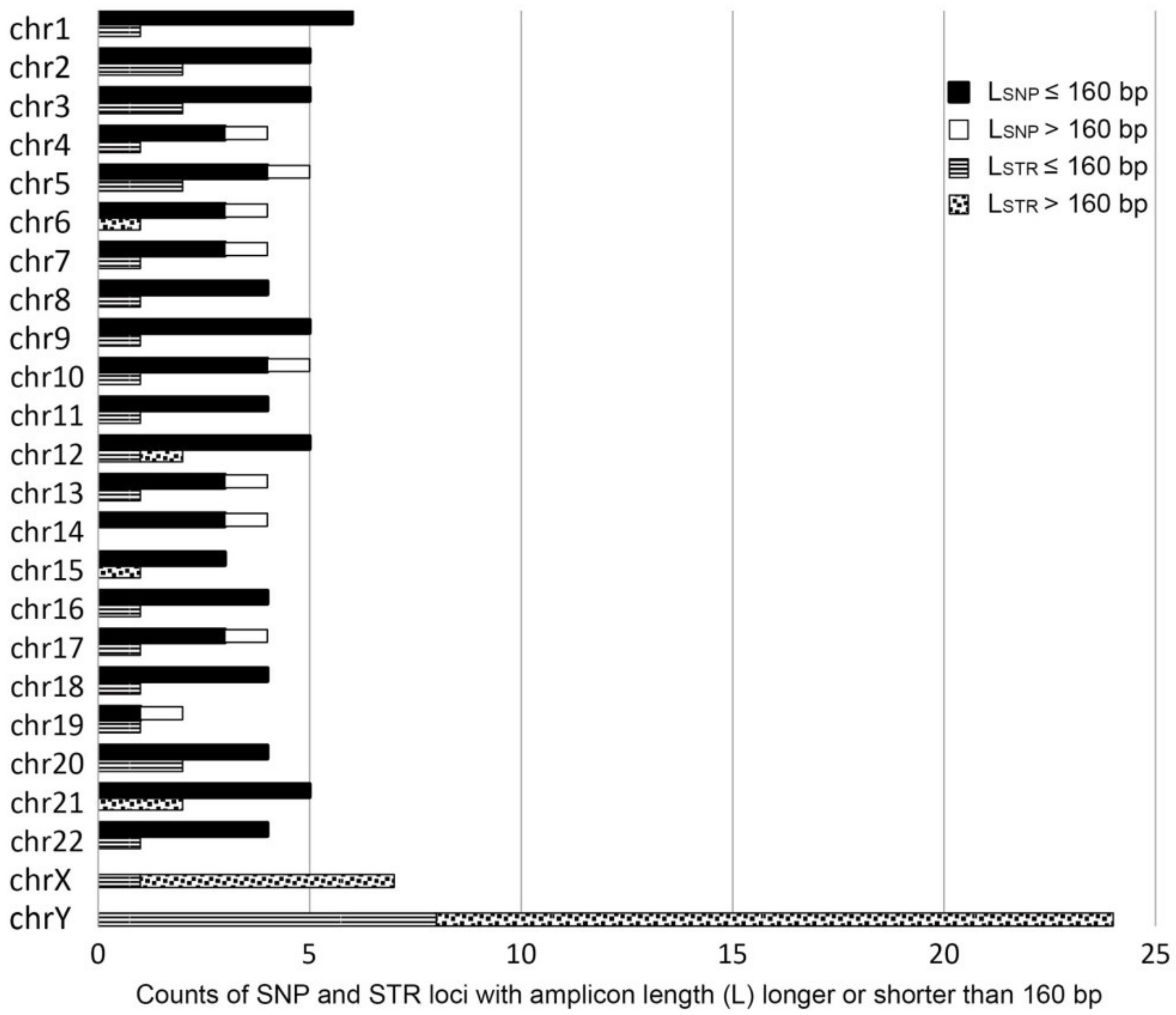
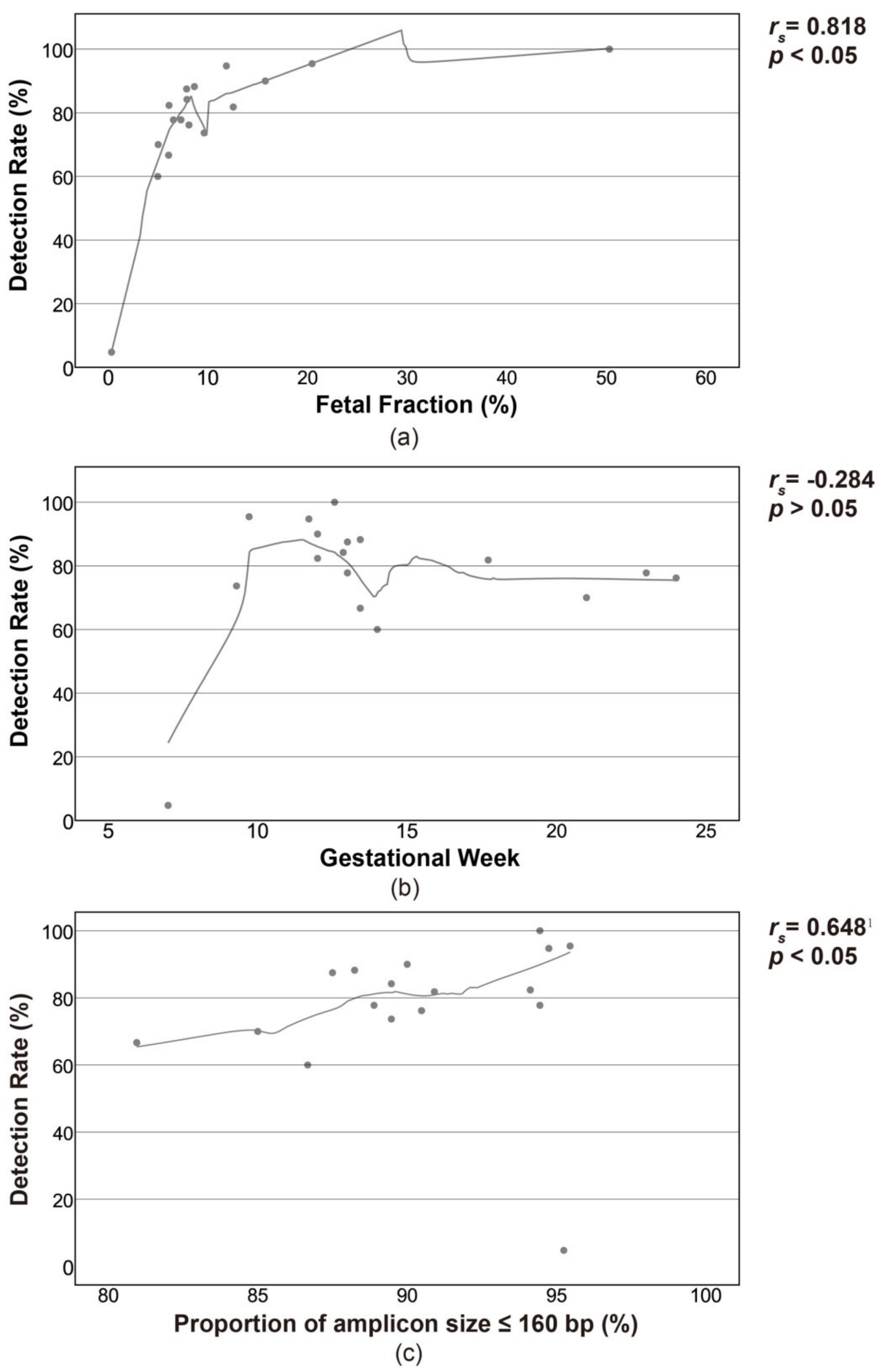
3.3. Influencing Factors on the Detection Rate
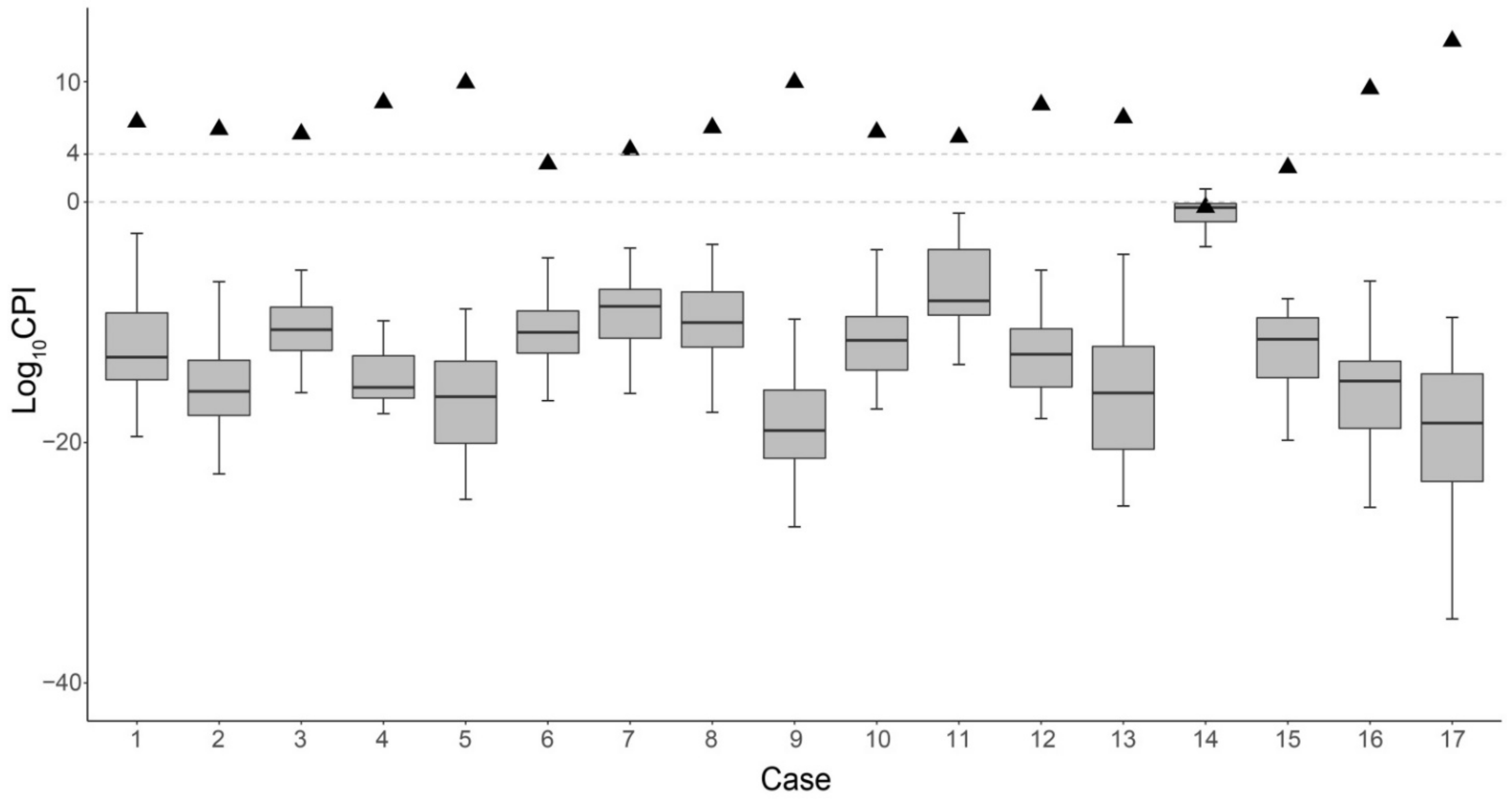
3.4. CPIs with SNP and STR Typing for NIPPT
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brambati, B.; Simoni, G.; Fabro, S. Chorionic villus Sampling: Fetal Diagnosis of Genetic Diseases in the First Trimester; M. Dekker: New York, NY, USA, 1986; p. 310. [Google Scholar]
- Brambati, B.; Simoni, G.; Travi, M.; Danesino, C.; Tului, L.; Privitera, O.; Stioui, S.; Tedeschi, S.; Russo, S.; Primignani, P. Genetic diagnosis by chorionic villus sampling before 8 gestational weeks: Efficiency, reliability, and risks on 317 completed pregnancies. Prenat. Diagn. 1992, 12, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Elchalal, U.; Ben Shachar, I.; Peleg, D.; Schenker, J.G. Maternal Mortality following Diagnostic 2nd-Trimester Amniocentesis. Fetal Diagn. Ther. 2004, 19, 195–198. [Google Scholar] [CrossRef]
- Firth, H.; Boyd, P.; Chamberlain, P.; MacKenzie, I.Z.; Morriss-Kay, G.; Huson, S. Analysis of limb reduction defects in babies exposed to chorionic villus sampling. Lancet 1994, 343, 1069–1071. [Google Scholar] [CrossRef]
- Cederholm, M.; Haglund, B.; Axelsson, O. Infant morbidity following amniocentesis and chorionic villus sampling for prenatal karyotyping. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 394–402. [Google Scholar] [CrossRef]
- Akolekar, R.; Beta, J.; Picciarelli, G.; Ogilvie, C.; D’Antonio, F. Procedure-related risk of miscarriage following amniocentesis and chorionic villus sampling: A systematic review and meta-analysis. Ultrasound Obstet. Gynecol. 2015, 45, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.M.D.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]
- Tong, Y.-K.; Lo, Y.D. Diagnostic developments involving cell-free (circulating) nucleic acids. Clin. Chim. Acta 2006, 363, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.C.; Lee, S.W.; Jiang, P.; Leung, T.Y.; Chan, K.A.; Chiu, R.W.; Lo, Y.D. High-Resolution Profiling of Fetal DNA Clearance from Maternal Plasma by Massively Parallel Sequencing. Clin. Chem. 2013, 59, 1228–1237. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, S.L.; Jakobsen, B.; Børsting, C.; Udengaard, H.; Buchard, A.; Kampmann, M.-L.; Grøndahl, M.L.; Morling, N. Non-invasive prenatal paternity testing using a standard forensic genetic massively parallel sequencing assay for amplification of human identification SNPs. Int. J. Leg. Med. 2019, 133, 1361–1368. [Google Scholar] [CrossRef]
- Tang, D.-L.; Li, Y.; Zhou, X.; Li, X.; Zheng, F. Multiplex fluorescent PCR for noninvasive prenatal detection of fetal-derived paternally inherited diseases using circulatory fetal DNA in maternal plasma. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 144, 35–39. [Google Scholar] [CrossRef]
- Wagner, J.; Džijan, S.; Marjanovic, D.; Lauc, G. Non-invasive prenatal paternity testing from maternal blood. Int. J. Leg. Med. 2008, 123, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, Z.; Gao, Y.; Yuan, Y.; Guo, Y.; Zhou, L.; Liao, K.; Wang, J.; Du, B.; Hou, Y.; et al. Effects of Maternal and Fetal Characteristics on Cell-Free Fetal DNA Fraction in Maternal Plasma. Reprod. Sci. 2015, 22, 1429–1435. [Google Scholar] [CrossRef]
- Fan, H.C.; Blumenfeld, Y.J.; Chitkara, U.; Hudgins, L.; Quake, S.R. Analysis of the Size Distributions of Fetal and Maternal Cell-Free DNA by Paired-End Sequencing. Clin. Chem. 2010, 56, 1279–1286. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Chen, P.; Lu, J.; Liu, Z.; Jia, E.; Ge, Q. The fragmentation patterns of maternal plasma cell-free DNA and its applications in non-invasive prenatal testing. Prenat. Diagn. 2020, 40, 911–917. [Google Scholar] [CrossRef]
- Jiang, H.; Xie, Y.; Li, X.; Ge, H.; Deng, Y.; Mu, H.; Feng, X.; Yin, L.; Du, Z.; Chen, F.; et al. Noninvasive Prenatal Paternity Testing (NIPAT) through Maternal Plasma DNA Sequencing: A Pilot Study. PLoS ONE 2016, 11, e0159385. [Google Scholar] [CrossRef]
- Ou, X.; Wang, H.; Qu, D.; Chen, Y.; Gao, J.; Sun, H. Epigenome-wide DNA methylation assay reveals placental epigenetic markers for noninvasive fetal single-nucleotide polymorphism genotyping in maternal plasma. Transfusion 2014, 54, 2523–2533. [Google Scholar] [CrossRef]
- Moriot, A.; Hall, D. Analysis of fetal DNA in maternal plasma with markers designed for forensic DNA mixture resolution. Genet. Med. 2019, 21, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Z.; Zhao, H.; Lin, S.; Huang, L.; He, Z.; Wang, H.; Ou, X. Evaluation of a Microhaplotype-Based Noninvasive Prenatal Test in Twin Gestations: Determination of Paternity, Zygosity, and Fetal Fraction. Genes 2020, 12, 26. [Google Scholar] [CrossRef]
- Chang, L.; Yu, H.; Miao, X.; Zhang, J.; Li, S. Development and comprehensive evaluation of a noninvasive prenatal paternity testing method through a scaled trial. Forensic Sci. Int. Genet. 2019, 43, 102158. [Google Scholar] [CrossRef] [PubMed]
- Qu, N.; Xie, Y.; Li, H.; Liang, H.; Lin, S.; Huang, E.; Gao, J.; Chen, F.; Shi, Y.; Ou, X. Noninvasive prenatal paternity testing using targeted massively parallel sequencing. Transfusion 2018, 58, 1792–1799. [Google Scholar] [CrossRef]
- Guo, F.; Yu, J.; Zhang, L.; Li, J. Massively parallel sequencing of forensic STRs and SNPs using the Illumina® ForenSeq™ DNA Signature Prep Kit on the MiSeq FGx™ Forensic Genomics System. Forensic Sci. Int. Genet. 2017, 31, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wu, R.; Li, H.; Zhang, Y.; Peng, D.; Wang, N.; Shen, X.; Wang, Z.; Sun, H. Characterizing stutter variants in forensic STRs with massively parallel sequencing. Forensic Sci. Int. Genet. 2020, 45, 102225. [Google Scholar] [CrossRef] [PubMed]
- Chim, S.S.C.; Tong, Y.K.; Chiu, R.W.K.; Lau, T.K.; Leung, T.N.; Chan, L.Y.S.; Oudejans, C.B.M.; Ding, C.; Lo, Y.M.D. Detection of the placental epigenetic signature of the maspin gene in maternal plasma. Proc. Natl. Acad. Sci. USA 2005, 102, 14753–14758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.K.; Ding, C.; Chiu, R.W.; Gerovassili, A.; Chim, S.S.; Leung, T.Y.; Leung, T.N.; Lau, T.K.; Nicolaides, K.H.; Lo, Y.D. Noninvasive Prenatal Detection of Fetal Trisomy 18 by Epigenetic Allelic Ratio Analysis in Maternal Plasma: Theoretical and Empirical Considerations. Clin. Chem. 2006, 52, 2194–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- State Administration for Market Regulation, China National Standardization Administration. Specification of Parentage Testing, GB/T 37223-2018, Issued on 2018-12-28. Available online: https://www.chinesestandard.net/PDF/English.aspx/GBT37223-2018 (accessed on 28 December 2018).
- Woerner, A.E.; King, J.L.; Budowle, B. Fast STR allele identification with STRait Razor 3.0. Forensic Sci. Int. Genet. 2017, 30, 18–23. [Google Scholar] [CrossRef]
- Wu, R.; Peng, D.; Ren, H.; Li, R.; Li, H.; Wang, N.; Shen, X.; Huang, E.; Zhang, Y.; Sun, H. Characterization of genetic polymorphisms in Nigerians residing in Guangzhou using massively parallel sequencing. Forensic Sci. Int. Genet. 2020, 48, 102323. [Google Scholar] [CrossRef]
- Novroski, N.M.; King, J.L.; Churchill, J.D.; Seah, L.H.; Budowle, B. Characterization of genetic sequence variation of 58 STR loci in four major population groups. Forensic Sci. Int. Genet. 2016, 25, 214–226. [Google Scholar] [CrossRef]
- Hernandis, E.; Dórum, G.; Egeland, T. relMix: An open source software for DNA mixtures with related contributors. Forensic Sci. Int. Genet. Suppl. Ser. 2019, 7, 221–223. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.; Budowle, B. How many familial relationship testing results could be wrong? PLoS Genet. 2020, 16, e1008929. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, Y.; Ren, H.; Li, H.; Li, R.; Shen, X.; Wang, N.; Huang, E.; Wu, R.; Sun, H. Identification of sequence polymorphisms at 58 STRs and 94 iiSNPs in a Tibetan population using massively parallel sequencing. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Yang, D.; Liang, H.; Gao, Y.; Lin, S.; He, Z.; Gao, J.; Sun, H.; Li, Q.; Ma, X.; Ou, X. Noninvasive fetal genotyping of paternally inherited alleles using targeted massively parallel sequencing in parentage testing cases. Transfusion 2017, 57, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Parson, W. Age Estimation with DNA: From Forensic DNA Fingerprinting to Forensic (Epi)Genomics: A Mini-Review. Gerontology 2018, 64, 326–332. [Google Scholar] [CrossRef]
- Schlötterer, C.; Tautz, D. Slippage synthesis of simple sequence DNA. Nucleic Acids Res. 1992, 20, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Woerner, A.E.; Mandape, S.; King, J.L.; Muenzler, M.; Crysup, B.; Budowle, B. Reducing noise and stutter in short tandem repeat loci with unique molecular identifiers. Forensic Sci. Int. Genet. 2020, 51, 102459. [Google Scholar] [CrossRef] [PubMed]
- Moray, N.; Pink, K.E.; Borry, P.; Larmuseau, M.H. Paternity testing under the cloak of recreational genetics. Eur. J. Hum. Genet. 2017, 25, 768–770. [Google Scholar] [CrossRef]
- Caenazzo, L.; Comacchio, A.; Tozzo, P.; Rodriguez, D.; Benciolini, P. Paternity testing requested by private parties in Italy: Some ethical considerations. J. Med. Ethics 2008, 34, 735–737. [Google Scholar] [CrossRef]
- Evans, M.I.; Andriole, S.; Pergament, E.; Speer, J.; Curtis, J.; Britt, D.W. Paternity Balancing. Fetal Diagn. Ther. 2013, 34, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Toya, W. Ethical, Legal and Social Issues in Japan on the Determination of Blood Relationship via DNA Testing. Asian Bioeth. Rev. 2017, 9, 19–32. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | SNPs | A-STRs | X-STRs | Y-STRs | ||||
|---|---|---|---|---|---|---|---|---|
| ePIAs | oPIAs | ePIAs | oPIAs | ePIAs | oPIAs | ePIAs | oPIAs | |
| 1 | 18 | 14 | 21 | 7 + 1 1 | 2 | 1 1 | 0 | 0 |
| 2 | 21 | 16 | 21 | 9 + 2 1 | 0 | 0 | 27 | 11 |
| 3 | 21 | 14 | 20 | 7 + 2 1 | 3 | 0 | 0 | 0 |
| 4 | 17 | 15 | 19 | 10 | 4 | 1 + 1 1 | 0 | 0 |
| 5 | 19 | 16 | 18 | 9 | 1 | 1 | 0 | 0 |
| 6 | 18 | 14 | 19 | 3 + 3 1 | 4 | 0 | 0 | 0 |
| 7 | 10 | 9 | 16 | 7 + 3 1 | 3 | 11 | 0 | 0 |
| 8 | 17 | 14 | 14 | 4 | 2 | 11 | 0 | 0 |
| 9 | 19 | 18 + 1 1 | 16 | 8 + 2 1 | 4 | 1 | 0 | 0 |
| 10 | 16 | 14 | 14 | 5 + 2 1 | 4 | 0 | 0 | 0 |
| 11 | 15 | 9 | 16 | 5 + 1 1 | 3 | 2 1 | 0 | 0 |
| 12 | 20 | 14 | 11 | 7 | 3 | 1 | 0 | 0 |
| 13 | 22 | 18 | 17 | 8 + 2 1 | 3 | 1 + 2 1 | 0 | 0 |
| 14 | 21 | 1 | 15 | 1 + 1 1 | 2 | 0 | 0 | 0 |
| 15 | 19 | 14 | 17 | 4 + 3 1 | 3 | 1 | 0 | 0 |
| 16 | 22 | 21 | 13 | 10 + 1 1 | 0 | 0 | 27 | 15 |
| 17 | 18 | 18 | 19 | 17 | 2 | 2 | 0 | 0 |
| Case | Sum of oPIAs for SNPs and A-STRs | Log10CPI Based on SNP Typing | Log10CPI Based on A-STR Typing | Log10CPI Based on SNP and A-STR Typing |
|---|---|---|---|---|
| 1 | 22 | 2.91 | 3.82 | 6.73 |
| 2 | 27 | 2.65 | 3.47 | 6.12 |
| 3 | 23 | 1.78 | 3.96 | 5.74 |
| 4 | 25 | 3.86 | 4.48 | 8.34 |
| 5 | 25 | 3.74 | 6.25 | 9.99 |
| 6 | 20 | 3.08 | 0.17 | 3.25 |
| 7 | 19 | 1.47 | 2.95 | 4.42 |
| 8 | 18 | 3.92 | 2.34 | 6.26 |
| 9 | 29 | 6.27 | 3.76 | 10.03 |
| 10 | 21 | 3.50 | 2.38 | 5.88 |
| 11 | 15 | 1.79 | 3.68 | 5.47 |
| 12 | 21 | 3.39 | 4.78 | 8.17 |
| 13 | 28 | 2.63 | 4.47 | 7.10 |
| 14 | 3 | 0.28 | −0.64 | −0.36 |
| 15 | 21 | 2.24 | 0.70 | 2.94 |
| 16 | 32 | 4.42 | 5.08 | 9.50 |
| 17 | 35 | 4.60 | 8.86 | 13.46 |
| LR Threshold | Real Parentage (n = 17) | Unrelated (n = 272) |
|---|---|---|
| >10 | 94.12% | 0.36% |
| >100 | 94.12% | 0 |
| >1000 | 88.24% | 0 |
| >10,000 | 82.35% | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Li, R.; Li, H.; Gao, Y.; Chen, H.; Qu, N.; Peng, D.; Wu, R.; Sun, H. Noninvasive Prenatal Paternity Testing with a Combination of Well-Established SNP and STR Markers Using Massively Parallel Sequencing. Genes 2021, 12, 454. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030454
Shen X, Li R, Li H, Gao Y, Chen H, Qu N, Peng D, Wu R, Sun H. Noninvasive Prenatal Paternity Testing with a Combination of Well-Established SNP and STR Markers Using Massively Parallel Sequencing. Genes. 2021; 12(3):454. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030454
Chicago/Turabian StyleShen, Xuefeng, Ran Li, Haixia Li, Yu Gao, Hui Chen, Ning Qu, Dan Peng, Riga Wu, and Hongyu Sun. 2021. "Noninvasive Prenatal Paternity Testing with a Combination of Well-Established SNP and STR Markers Using Massively Parallel Sequencing" Genes 12, no. 3: 454. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030454