
Improvement of a Yairipok Chujak Maize Landrace from North Eastern Himalayan Region for β-Carotene Content through Molecular Marker-Assisted Backcross Breeding


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
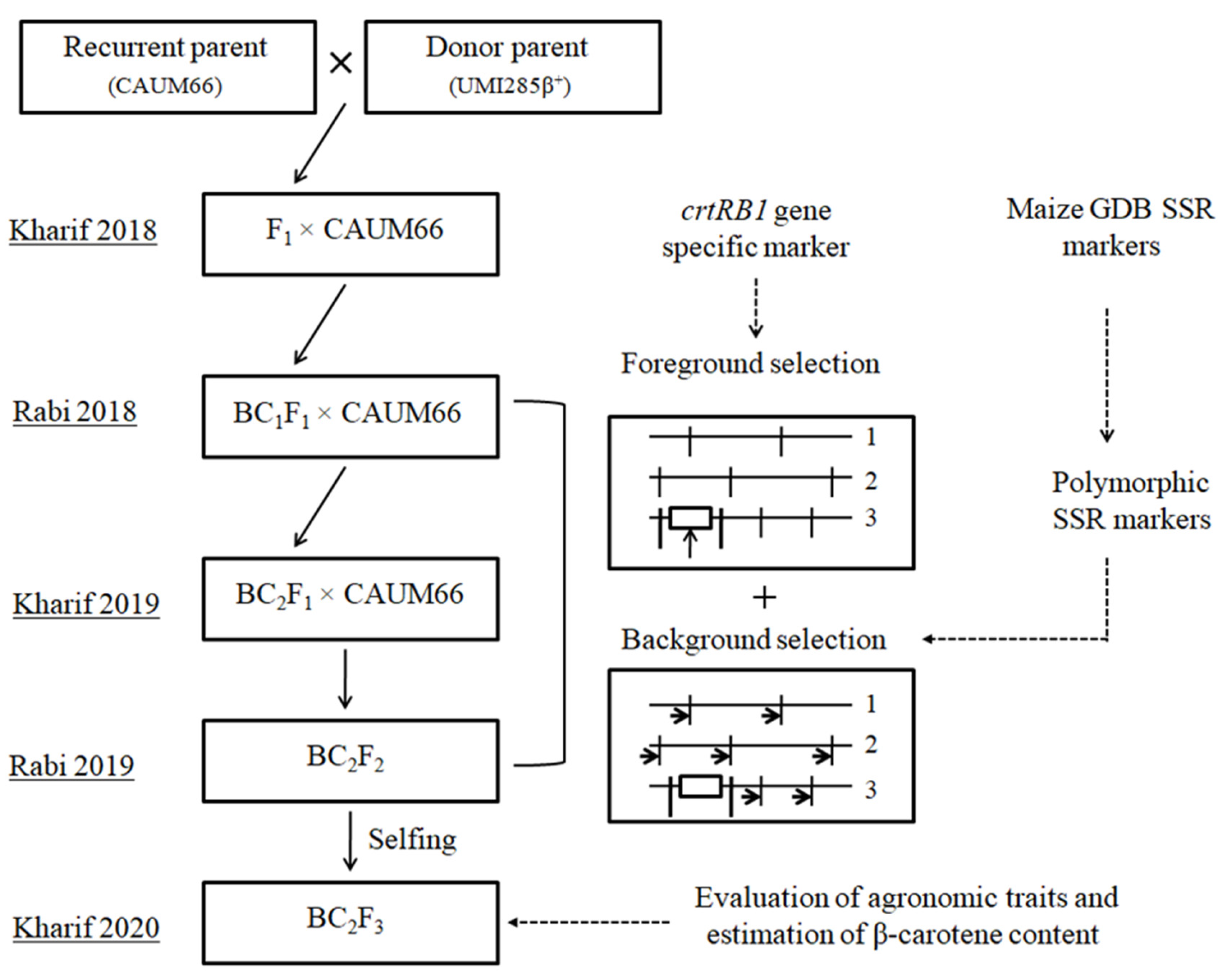
2.1. Plant Materials and Generation of Backcross Population
2.2. MABC Breeding Strategy
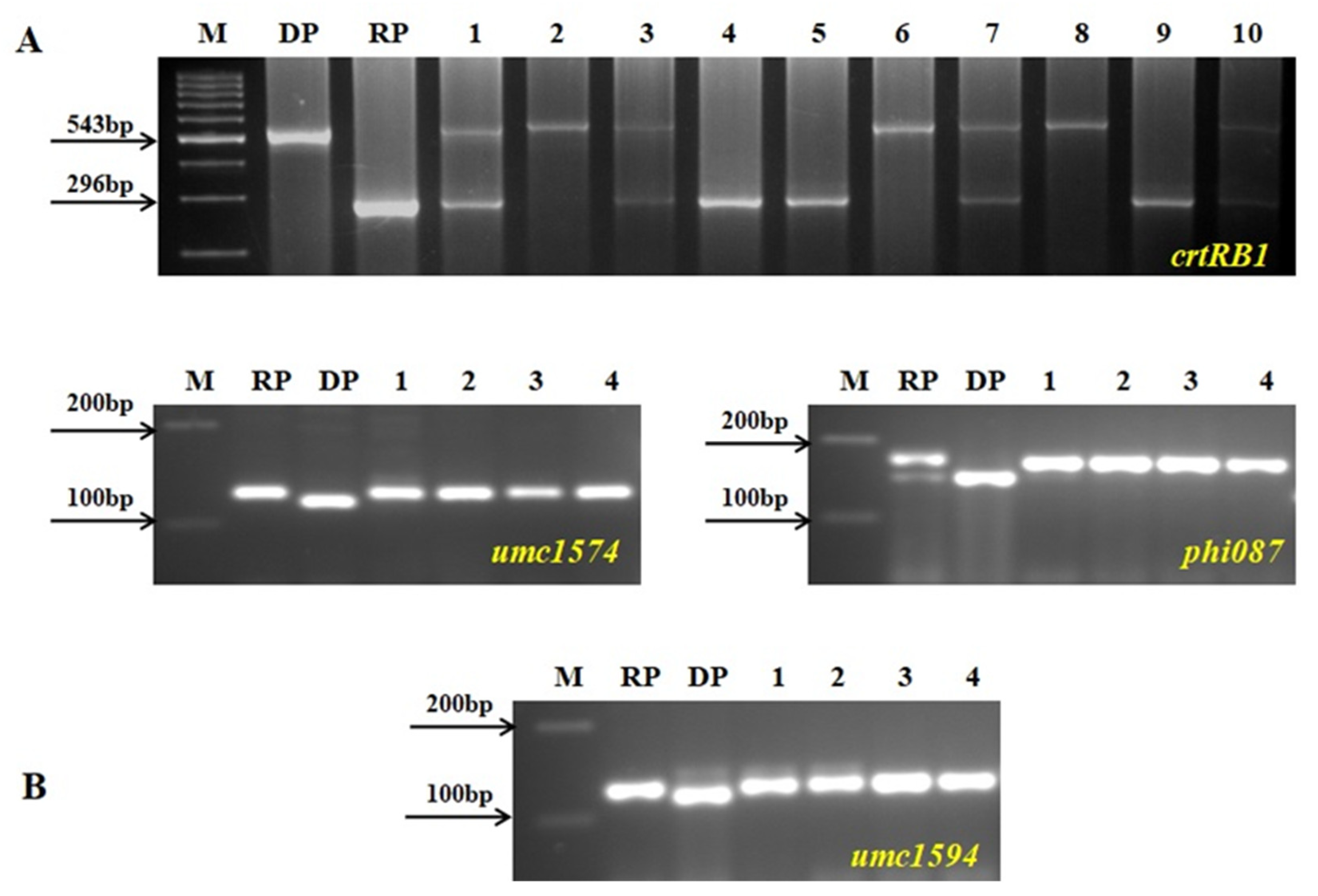
2.3. Molecular Marker Analysis
2.4. Evaluation of Major Agronomic Traits of the Selected Lines
2.5. Characterization of the Selected Lines for β-Carotene Content
3. Results
3.1. Incorporation of β-Carotene Gene crtRB1 into the Background of CAUM66
3.2. Agronomic Traits of the Selected Lines
3.3. β-Carotene Content of the Selected Lines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agricultural Organization of the United Nations. FAO STAT. Import and Export Value of Maize. Available online: http://www.fao.org/faostat/en/#rankings/countries_by_commodity (accessed on 2 February 2021).
- Natesan, S.; Singh, T.; Duraisamy, T.; Chandrasekharan, N.; Chandran, S.; Adhimoolam, K.; Muniyandi, S.; Sampathrajan, V.; Kalipatty Nalliappan, G.; Muthurajan, R. Characterization of crtRB1 Gene Polymorphism and and β carotene content in Maize Landrances Originated from North Eastern Himalayan Region (NEHR) of India. Front. Sustain. Food Syst. 2020. [Google Scholar] [CrossRef]
- Saha, S.; Lungmuana, D.; Ansari, M.; Jat, S.; Chowdhury, S.; Saithantluanga, H.; Rakshit, S.; Prakash, N. Souvenir of National Workshop on Scientific Maize Cultivation in North East India held on 5 March, 2019 at SAMETI Training Hall, Aizawl, Mizoram. Available online: https://krishi.icar.gov.in/jspui/handle/123456789/20143 (accessed on 8 March 2021).
- Prasanna, B.M.; Palacios-Rojas, N.; Hossain, F.; Muthusamy, V.; Menkir, A.; Dhliwayo, T.; Ndhlela, T.; San Vicente, F.; Nair, S.K.; Vivek, B.S. Molecular breeding for nutritionally enriched maize: Status and prospects. Front. Genet. 2020, 10, 1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harjes, C.E.; Rocheford, T.R.; Bai, L.; Brutnell, T.P.; Kandianis, C.B.; Sowinski, S.G.; Stapleton, A.E.; Vallabhaneni, R.; Williams, M.; Wurtzel, E.T. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification. Science 2008, 319, 330–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Kandianis, C.B.; Harjes, C.E.; Bai, L.; Kim, E.-H.; Yang, X.; Skinner, D.J.; Fu, Z.; Mitchell, S.; Li, Q. Rare genetic variation at Zea mays crtRB1 increases β-carotene in maize grain. Nat. Genet. 2010, 42, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, R.; Rojas, N.P.; Gao, S.; Yan, J.; Pixley, K. Validation of the effects of molecular marker polymorphisms in LcyE and CrtRB1 on provitamin A concentrations for 26 tropical maize populations. Theor. Appl. Genet. 2013, 126, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, V.; Hossain, F.; Thirunavukkarasu, N.; Choudhary, M.; Saha, S.; Bhat, J.S.; Prasanna, B.M.; Gupta, H.S. Development of β-carotene rich maize hybrids through marker-assisted introgression of β-carotene hydroxylase allele. PLoS ONE 2014, 9, e113583. [Google Scholar] [CrossRef] [Green Version]
- Chandran, S.; Pukalenthy, B.; Adhimoolam, K.; Manickam, D.; Sampathrajan, V.; Chocklingam, V.; Eswaran, K.; Arunachalam, K.; Rajasekaran, R.; Muthusamy, V. Marker-assisted selection to pyramid the opaque-2 (o2) and β-carotene (crtRB1) genes in maize. Front. Genet. 2019, 10, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagare, D.B.; Shetti, P.; Surender, M.; Reddy, S. Marker-assisted backcross breeding for enhancing β-carotene of QPM inbreds. Mol. Breed. 2019, 39, 1–12. [Google Scholar] [CrossRef]
- Natesan, S.; Duraisamy, T.; Pukalenthy, B.; Chandran, S.; Nallathambi, J.; Adhimoolam, K.; Manickam, D.; Sampathrajan, V.; Muniyandi, S.J.; Meitei, L.J. Enhancing β-carotene concentration in parental lines of CO6 maize hybrid through marker-assisted backcross breeding (MABB). Front. Nutr. 2020, 7. [Google Scholar] [CrossRef]
- Selvi, D.T. Marker Assisted Selection of Favourable crtRB1 Allele for Enhanced Beta Carotene in Maize and Its Effect on Seed Storability. Ph.D. Thesis, Tamil Nadu Agricultural University, Coimbatore, India, 31 October 2013. [Google Scholar]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, N.S.; Sarma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus Handbook for Carotenoid Analysis; Harvest Plus: Washington, DC, USA, 2004; Volume 2. [Google Scholar]
- Kurilich, A.C.; Juvik, J.A. Quantification of Carotenoid and Tocopherol Antioxidants in Zea m ays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef]
- Nass, L.L.; Pellicano, I.J.; Valois, A.C.C. Utilization of genetic resources for maize and soybean breeding in Brazil. Rev. Bras. Genet. 1993, 16, 983–988. [Google Scholar]
- Singode, A.; Sekhar, J.; Srinivasan, K.; Prasanna, B. Evaluation of yield performance of selected North East Himalayan (NEH) maize landrace accessions of India, outside their habitat. Indian J. Genet. Plant Breed. 2009, 69, 191–198. [Google Scholar]
- Pandey, A.; Bhandari, D.; Bhatt, K.; Pareek, S.; Tomer, A.; Dhillon, B. Wild Relatives of Cflip Plants in India: Collection and Conservation; National Bureau of Plant Genetic Resources: New Delhi, India, 2005. [Google Scholar]
- Baseggio, M.; Murray, M.; Magallanes-Lundback, M.; Kaczmar, N.; Chamness, J.; Buckler, E.S.; Smith, M.E.; DellaPenna, D.; Tracy, W.F.; Gore, M.A. Natural variation for carotenoids in fresh kernels is controlled by uncommon variants in sweet corn. Plant Genome 2020, 13, e20008. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A.; Ashkani, S.; Malek, M.A.; Latif, M.A. Marker-assisted backcrossing: A useful method for rice improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Ribaut, J.M.; Jiang, C.; Hoisington, D. Simulation experiments on efficiencies of gene introgression by backcrossing. Crop Sci. 2002, 42, 557–565. [Google Scholar] [CrossRef]
- Yang, D.; Tang, J.; Yang, D.; Chen, Y.; Ali, J.; Mou, T. Improving rice blast resistance of Feng39S through molecular marker-assisted backcrossing. Rice 2019, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pukalenthy, B.; Manickam, D.; Chandran, S.; Adhimoolam, K.; Sampathrajan, V.; Rajasekaran, R.; Arunachalam, K.; Ganapathyswamy, H.; Chocklingam, V.; Muthusamy, V. Incorporation of opaque-2 into ‘UMI 1200’, an elite maize inbred line, through marker-assisted backcross breeding. Biotechnol. Biotechnol. Equip. 2019, 33, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Pukalenthy, B.; Manickam, D.; Adhimoolam, K.; Mahesh, S.G.; Ramanathan, N.; Chandran, S.; Sampathrajan, V.; Rajasekaran, R.; Arunachalam, K.; Senthil, K. Marker aided introgression of opaque 2 (o2) allele improving lysine and tryptophan in maize (Zea mays L.). Physiol. Mol. Biol. Plants 2020, 26, 1925–1930. [Google Scholar] [CrossRef]
- Mehta, B.K.; Muthusamy, V.; Zunjare, R.U.; Baveja, A.; Chauhan, H.S.; Chhabra, R.; Singh, A.K.; Hossain, F. Biofortification of sweet corn hybrids for provitamin-A, lysine and tryptophan using molecular breeding. J. Cereal Sci. 2020, 96, 103093. [Google Scholar] [CrossRef]
- Sarika, K.; Hossain, F.; Muthusamy, V.; Zunjare, R.U.; Baveja, A.; Goswami, R.; Thirunavukkarasu, N.; Jha, S.K.; Gupta, H.S. Opaque16, a high lysine and tryptophan mutant, does not influence the key physico-biochemical characteristics in maize kernel. PLoS ONE 2018, 13, e0190945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zunjare, R.U.; Hossain, F.; Muthusamy, V.; Baveja, A.; Chauhan, H.S.; Bhat, J.S.; Thirunavukkarasu, N.; Saha, S.; Gupta, H.S. Development of biofortified maize hybrids through marker-assisted stacking of β-carotene hydroxylase, lycopene-ε-cyclase and opaque2 genes. Front. Plant Sci. 2018, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.; Zunjare, R.U.; Khan, S.; Baveja, A.; Muthusamy, V.; Hossain, F. Marker-assisted introgression of rare allele of β-carotene hydroxylase (crtRB1) gene into elite quality protein maize inbred for combining high lysine, tryptophan and provitamin A in maize. Plant Breed. 2019, 138, 174–183. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| S. No. | Generation | No. of Plants Screened | Favorable Allele (1) | Heterozygotes (1 and 3) | Unfavorable Allele (3) | Chi-Square Value | p-Value |
|---|---|---|---|---|---|---|---|
| 1. | BC1F1 | 193 | 0 | 124 | 69 | 15.673 | 0.000075 ** |
| 2. | BC2F1 | 198 | 0 | 114 | 84 | 4.545 | 0.033006 ns |
| 3. | BC2F2 | 129 | 36 | 64 | 29 | 0.7674 | 0.681321 ns |
| S. No. | Generations | Progenies Selected | RPG (%) | No. of Markers Restored | Range of RPG (%) among the Selected Progenies |
|---|---|---|---|---|---|
| 1. | BC1F1 | CAUM66-2-27 | 63.75 | 59–63 | 61.82–64.34 |
| CAUM66-28-35 | 62.96 | ||||
| CAUM66-15-12 | 61.82 | ||||
| CAUM66-54-9 | 64.34 | ||||
| 2. | BC2F1 | CAUM66-54-9-7 | 82.12 | 78–84 | 79.31–83.22 |
| CAUM66-54-9-11 | 81.01 | ||||
| CAUM66-54-9-12 | 83.22 | ||||
| CAUM66-54-9-18 | 79.31 | ||||
| 3. | BC2F2 | CAUM66-54-9-12-2 | 86.74 | 83–94 | 86.74–90.16 |
| CAUM66-54-9-12-11 | 87.71 | ||||
| CAUM66-54-9-12-13 | 88.86 | ||||
| CAUM66-54-9-12-24 | 90.16 |
| S. No. | Traits | Recurrent Parent | Donor Parent | CAUM66-54-9-12-2 | CAUM66-54-9-12-11 | CAUM66-54-9-12-13 | CAUM66-54-9-12-24 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Mean | Mean | RGP% | Mean | RGP % | Mean | RGP % | Mean | RGP % | ||
| 1 | Days to tasselling | 59.08 | 57.64 | 57.24 | 96.88 | 56.47 | 95.58 | 55.73 | 94.37 | 57.34 | 97.05 |
| 2 | Days to silking | 61.04 | 61.01 | 60.21 | 98.64 | 59.84 | 98.03 | 58.35 | 95.59 | 59.57 | 97.59 |
| 3 | Plant height | 168.82 | 120.60 | 157.48 | 93.28 | 143.25 | 84.85 | 167.89 | 99.44 | 161.45 | 95.63 |
| 4 | Ear placement height | 69.78 | 58.60 | 65.47 | 93.82 | 63.47 | 90.95 | 64.25 | 92.07 | 65.48 | 93.83 |
| 5 | Tassel length | 36.80 | 19.02 | 33.29 | 90.46 | 34.25 | 93.07 | 31.27 | 84.97 | 35.87 | 97.47 |
| 6 | Number of tassel branches | 08.04 | 20.32 | 07.58 | 94.27 | 07.54 | 93.78 | 07.58 | 94.27 | 07.53 | 93.65 |
| 7 | Leaf length | 60.09 | 56.32 | 54.25 | 90.28 | 54.27 | 90.31 | 54.28 | 90.33 | 54.78 | 91.16 |
| 8 | Leaf breadth | 07.41 | 06.31 | 06.28 | 84.75 | 06.59 | 88.93 | 06.57 | 88.66 | 06.87 | 92.71 |
| 9 | Cob length | 16.29 | 13.16 | 14.65 | 89.93 | 13.59 | 83.42 | 15.68 | 96.25 | 15.41 | 94.59 |
| 10 | Number of kernel rows per cob | 11.12 | 10.58 | 10.74 | 96.58 | 10.14 | 91.18 | 9.25 | 83.18 | 9.57 | 86.06 |
| 11 | Number of kernels per row | 19.84 | 26.62 | 17.69 | 89.16 | 16.25 | 81.90 | 15.64 | 78.83 | 18.21 | 91.78 |
| 12 | 100 kernel weight | 25.78 | 23.85 | 21.45 | 83.20 | 22.87 | 88.71 | 21.87 | 84.83 | 24.36 | 94.49 |
| 13 | Cob weight | 95.51 | 74.93 | 80.64 | 84.43 | 80.54 | 84.32 | 85.49 | 89.50 | 88.74 | 92.91 |
| 14 | Single plant yield | 66.57 | 62.31 | 64.33 | 96.63 | 62.34 | 93.64 | 65.34 | 98.15 | 63.58 | 95.50 |
| 15 | Beta carotene content (µg/g) | 0.86 | 9.214 | 7.541 | 8.258 | 8.647 | 8.711 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qutub, M.; Chandran, S.; Rathinavel, K.; Sampathrajan, V.; Rajasekaran, R.; Manickam, S.; Adhimoolam, K.; Muniyandi, S.J.; Natesan, S. Improvement of a Yairipok Chujak Maize Landrace from North Eastern Himalayan Region for β-Carotene Content through Molecular Marker-Assisted Backcross Breeding. Genes 2021, 12, 762. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050762
Qutub M, Chandran S, Rathinavel K, Sampathrajan V, Rajasekaran R, Manickam S, Adhimoolam K, Muniyandi SJ, Natesan S. Improvement of a Yairipok Chujak Maize Landrace from North Eastern Himalayan Region for β-Carotene Content through Molecular Marker-Assisted Backcross Breeding. Genes. 2021; 12(5):762. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050762
Chicago/Turabian StyleQutub, Maqbool, Sarankumar Chandran, Krishnakumar Rathinavel, Vellaikumar Sampathrajan, Ravikesavan Rajasekaran, Sudha Manickam, Karthikeyan Adhimoolam, Samuel Jeberson Muniyandi, and Senthil Natesan. 2021. "Improvement of a Yairipok Chujak Maize Landrace from North Eastern Himalayan Region for β-Carotene Content through Molecular Marker-Assisted Backcross Breeding" Genes 12, no. 5: 762. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050762