
SLC6A3 (DAT1) as a Novel Candidate Biomarker Gene for Suicidal Behavior

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
- -
- Patients cohort included 80 females and 20 males, the table shows the values for the total sample, including females and males.
- -
- BDI—Beck’s Depression Inventory.
- -
- HAMD—Hamilton’s Depression Rating Scale.
- -
- SA—Situational anxiety.
- -
- PA—Personal anxiety.
2.2. DNA Isolation and Genotyping
2.3. Statistical Analysis
- -
- Codominant. This model assumes that each genotype can influence risk independently of the others.
- -
- Dominant. Common allele homozygotes were tested against rare allele homo- and heterozygotes.
- -
- Recessive. Rare allele homozygotes were tested against common allele homo- and heterozygotes.
- -
- Overdominant. Heterozygotes were tested against both homozygotes.
- -
- Log-additive. A trend test for the genotypes; according to this model, each allele changes the risk in an additive manner (i.e., the presence of two alleles doubles the risk compared to the presence of only one allele). The test was based on a logistic regression model and genotypes were coded as 0, 1, or 2, depending on the amount of minor alleles.
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fitzgerald, P.; Kulkarni, J. Suicide rates and mental health disorder prevention. Aust. N. Z. J. Psychiatry 2014, 49, 91–92. [Google Scholar] [CrossRef]
- Voracek, M.; Loibl, L.M. Genetics of suicide: A systematic review of twin studies. Wien. Klin. Wochenschr. 2007, 119, 463–475. [Google Scholar] [CrossRef]
- Fu, Q.; Heath, A.C.; Bucholz, K.K.; Nelson, E.C.; Glowinski, A.L.; Goldberg, J.; Lyons, M.; Tsuang, M.T.; Jacob, T.; True, M.R.; et al. A twin study of genetic and environmental influences on suicidality in men. Psychol. Med. 2002, 32, 11–24. [Google Scholar] [CrossRef]
- Glowinski, A.L.; Bucholz, K.K.; Nelson, E.C.; Fu, Q.; Madden, P.A.; Reich, W.; Heath, A.C. Suicide Attempts in an Adolescent Female Twin Sample. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 1300–1307. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Segal, N.L. Suicidal behavior in twins: A replication. J. Affect. Disord. 2001, 66, 71–74. [Google Scholar] [CrossRef]
- Von Borczyskowski, A.; Lindblad, F.; Vinnerljung, B.; Reintjes, R.; Hjern, A. Familial factors and suicide: An adoption study in a Swedish National Cohort. Psychol. Med. 2010, 41, 749–758. [Google Scholar] [CrossRef]
- Brent, D.A.; Mann, J.J. Family genetic studies, suicide, and suicidal behavior. Am. J. Med. Genet. Part C Semin. Med. Genet. 2005, 133C, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Tidemalm, D.; Runeson, B.; Waern, M.; Frisell, T.; Carlström, E.; Lichtenstein, P.; Långström, N. Familial clustering of suicide risk: A total population study of 11.4 million individuals. Psychol. Med. 2011, 41, 2527–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, J.J.; Henteleff, R.A.; Lagattuta, T.F.; Perper, J.A.; Li, S.; Arango, V. Lower3H-paroxetine binding in cerebral cortex of suicide victims is partly due to fewer high affinity, non-transporter sites. J. Neural Transm. 1996, 103, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.J.; Malone, K.M. Cerebrospinal fluid amines and higher-lethality suicide attempts in depressed inpatients. Biol. Psychiatry 1997, 41, 162–171. [Google Scholar] [CrossRef]
- Sher, L.; Mann, J.J.; Traskman-Bendz, L.; Winchel, R.; Huang, Y.-Y.; Fertuck, E.; Stanley, B.H. Lower cerebrospinal fluid homovanillic acid levels in depressed suicide attempters. J. Affect. Disord. 2006, 90, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Samuelsson, M.; Jokinen, J.; Nordström, A.; Nordstrom, P. CSF 5-HIAA, suicide intent and hopelessness in the prediction of early suicide in male high-risk suicide attempters. Acta Psychiatr. Scand. 2006, 113, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.-P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of Anxiety-Related Traits with a Polymorphism in the Serotonin Transporter Gene Regulatory Region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Heils, A.; Teufel, A.; Petri, S.; Stöber, G.; Riederer, P.; Bengel, D.; Lesch, K.P. Allelic Variation of Human Serotonin Transporter Gene Expression. J. Neurochem. 2002, 66, 2621–2624. [Google Scholar] [CrossRef]
- Hu, X.-Z.; Lipsky, R.H.; Zhu, G.; Akhtar, L.A.; Taubman, J.; Greenberg, B.D.; Xu, K.; Arnold, P.D.; Richter, M.A.; Kennedy, J.L.; et al. Serotonin Transporter Promoter Gain-of-Function Genotypes Are Linked to Obsessive-Compulsive Disorder. Am. J. Hum. Genet. 2006, 78, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanelli, G.; Serretti, A. The influence of the serotonin transporter gene 5-HTTLPR polymorphism on suicidal behaviors: A meta-analysis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 88, 375–387. [Google Scholar] [CrossRef]
- Czesak, M.; Lemonde, S.; Peterson, E.A.; Rogaeva, A.; Albert, P.R. Cell-Specific Repressor or Enhancer Activities of Deaf-1 at a Serotonin 1A Receptor Gene Polymorphism. J. Neurosci. 2006, 26, 1864–1871. [Google Scholar] [CrossRef] [Green Version]
- Lemonde, S.; Turecki, G.; Bakish, D.; Du, L.; Hrdina, P.D.; Bown, C.D.; Sequeira, A.; Kushwaha, N.; Morris, S.J.; Basak, A.; et al. Impaired Repression at a 5-Hydroxytryptamine 1A Receptor Gene Polymorphism Associated with Major Depression and Suicide. J. Neurosci. 2003, 23, 8788–8799. [Google Scholar] [CrossRef] [Green Version]
- Castro, T.B.G.; Tovilla-Zárate, C.A.; Juárez-Rojop, I.; García, S.P.; Genis, A.; Nicolini, H.; Narváez, L.L. Association of 5HTR1A gene variants with suicidal behavior: Case-control study and updated meta-analysis. J. Psychiatr. Res. 2013, 47, 1665–1672. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Sanders, A.R.; Molen, J.E.V.; Martinolich, L.; Mowry, B.J.; Levinson, D.F.; Crowe, R.R.; Silverman, J.M.; Gejman, P.V. Polymorphisms in the 5′-untranslated region of the human serotonin receptor 1B (HTR1B) gene affect gene expression. Mol. Psychiatry 2003, 8, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Kia-Keating, B.M.; Glatt, S.J.; Tsuang, M.T. Meta-analyses suggest association between COMT, but not HTR1B, alleles, and suicidal behavior. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2007, 144B, 1048–1053. [Google Scholar] [CrossRef]
- Parsons, M.J.; D’Souza, U.M.; Arranz, M.-J.; Kerwin, R.W.; Makoff, A.J. The –1438A/G polymorphism in the 5-hydroxytryptamine type 2A receptor gene affects promoter activity. Biol. Psychiatry 2004, 56, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Duan, Y.; He, L. Association study of serotonin 2A receptor (5-HT2A) gene with schizophrenia and suicidal behavior using systematic meta-analysis. Biochem. Biophys. Res. Commun. 2006, 340, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Fuke, S.; Suo, S.; Takahashi, N.; Koike, H.; Sasagawa, N.; Ishiura, S. The VNTR polymorphism of the human dopamine transporter (DAT1) gene affects gene expression. Pharm. J. 2001, 1, 152–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue-Murayama, M.; Adachi, S.; Mishima, N.; Mitani, H.; Takenaka, O.; Terao, K.; Hayasaka, I.; Ito, S.; Murayama, Y. Variation of variable number of tandem repeat sequences in the 3′-untranslated region of primate dopamine transporter genes that affects reporter gene expression. Neurosci. Lett. 2002, 334, 206–210. [Google Scholar] [CrossRef]
- Miller, G.M.; Madras, B.K. Polymorphisms in the 3′-untranslated region of human and monkey dopamine transporter genes affect reporter gene expression. Mol. Psychiatry 2002, 7, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanness, S.H.; Owens, M.J.; Kilts, C.D. The variable number of tandem repeats element in DAT1 regulates in vitro dopamine transporter density. BMC Genet. 2005, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Bieliński, M.; Jaracz, M.; Lesiewska, N.; Tomaszewska, M.; Sikora, M.; Junik, R.; Kamińska, A.; Tretyn, A.; Borkowska, A. Association between COMT Val158Met and DAT1 polymorphisms and depressive symptoms in the obese population. Neuropsychiatr. Dis. Treat. 2017, ume 13, 2221–2229. [Google Scholar] [CrossRef] [Green Version]
- Joyce, P.R.; McHugh, P.C.; Light, K.J.; Rowe, S.; Miller, A.L.; Kennedy, M.A. Relationships Between Angry-Impulsive Personality Traits and Genetic Polymorphisms of the Dopamine Transporter. Biol. Psychiatry 2009, 66, 717–721. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, U.M.; Russ, C.; Tahir, E.; Mill, J.; McGuffin, P.; Asherson, P.; Craig, I.W. Functional effects of a tandem duplication polymorphism in the 5′flanking region of the DRD4 gene. Biol. Psychiatry 2004, 56, 691–697. [Google Scholar] [CrossRef]
- Doorley, J.; Williams, C.; Mallard, T.; Esposito-Smythers, C.; McGeary, J. Sexual Trauma, the Dopamine D4 Receptor, and Suicidal Ideation Among Hospitalized Adolescents: A Preliminary Investigation. Arch. Suicide Res. 2016, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zalsman, G.; Frisch, A.; Lewis, R.; Michaelovsky, E.; Hermesh, H.; Sher, L.; Nahshoni, E.; Wolovik, L.; Tyano, S.; Apter, A.; et al. DRD4 receptor gene exon III polymorphism in inpatient suicidal adolescents. J. Neural Transm. 2004, 111, 1593–1603. [Google Scholar] [CrossRef]
- Munafò, M.R.; Yalcin, B.; Willis-Owen, S.; Flint, J. Association of the Dopamine D4 Receptor (DRD4) Gene and Approach-Related Personality Traits: Meta-Analysis and New Data. Biol. Psychiatry 2008, 63, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Thomas, N.; Singleton, A.; Piggott, M.; Lloyd, S.; Perry, E.K.; Morris, C.M.; Perry, R.H.; Ferrier, N.R.; Court, J.A. D2 dopamine receptor gene (DRD2) Taq1 A polymorphism: Reduced dopamine D2 receptor binding in the human striatum associated with the A1 allele. Pharmacogenetics 1997, 7, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Suda, A.; Kawanishi, C.; Kishida, I.; Sato, R.; Yamada, T.; Nakagawa, M.; Hasegawa, H.; Kato, D.; Furuno, T.; Hirayasu, Y. Dopamine D2 Receptor Gene Polymorphisms Are Associated with Suicide Attempt in the Japanese Population. Neuropsychobiology 2009, 59, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Jasiewicz, A.; Samochowiec, A.; Samochowiec, J.; Małecka, I.; Suchanecka, A.; Grzywacz, A. Suicidal Behavior and Haplotypes of the Dopamine Receptor Gene (DRD2) and ANKK1 Gene Polymorphisms in Patients with Alcohol Dependence—Preliminary Report. PLoS ONE 2014, 9, e111798. [Google Scholar] [CrossRef]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.-M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Gonzalez-Castro, T.B.; Hernandez-Diaz, Y.; Juarez-Rojop, I.E.; López-Narváez, M.L.; Tovilla-Zárate, C.A.; Ramirez-Bello, J.; Pérez-Hernández, N.; Genis-Mendoza, A.D.; Fresan, A.; Guzmán-Priego, C.G. The role of COMT gene Val108/158Met polymorphism in suicidal behavior: Systematic review and updated meta-analysis. Neuropsychiatr. Dis Treat. 2018, 14, 2485–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhagen, M.; Van Der Meij, A.; Van Deurzen, P.; Janzing, J.; Arias-Vasquez, A.; Buitelaar, J.; Franke, B. Meta-analysis of the BDNF Val66Met polymorphism in major depressive disorder: Effects of gender and ethnicity. Mol. Psychiatry 2008, 15, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Zai, C.C.; Manchia, M.; De Luca, V.; Tiwari, A.K.; Chowdhury, N.I.; Zai, G.C.; Tong, R.P.; Yilmaz, Z.; Shaikh, S.A.; Strauss, J.; et al. The brain-derived neurotrophic factor gene in suicidal behaviour: A meta-analysis. Int. J. Neuropsychopharmacol. 2012, 15, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Hranilovic, D.; Stefulj, J.; Furac, I.; Kubat, M.; Balija, M.; Jernej, B. Serotonin transporter gene promoter (5-HTTLPR) and intron 2 (VNTR) polymorphisms in Croatian suicide victims. Biol. Psychiatry 2003, 54, 884–889. [Google Scholar] [CrossRef]
- Pungercic, G.; Videtic, A.; Pestotnik, A.; Pajnic, I.Z.; Zupanc, T.; Balazic, J.; Tomori, M.; Komel, R. Serotonin transporter gene promoter (5-HTTLPR) and intron 2 (VNTR) polymorphisms: A study on Slovenian population of suicide victims. Psychiatr. Genet. 2006, 16, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Videtic, A.; Pungercic, G.; Pajnic, I.Z.; Zupanc, T.; Balazic, J.; Tomori, M.; Komel, R. Association study of seven polymorphisms in four serotonin receptor genes on suicide victims. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141B, 669–672. [Google Scholar] [CrossRef]
- Videtic, A.; Zupanc, T.; Pregelj, P.; Balažic, J.; Tomori, M.; Komel, R. Suicide, stress and serotonin receptor 1A promoter polymorphism -1019C>G in Slovenian suicide victims. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Pregelj, P.; Nedic, G.; Paska, A.V.; Zupanc, T.; Nikolac, M.; Balažic, J.; Tomori, M.; Komel, R.; Seler, D.M.; Pivac, N. The association between brain-derived neurotrophic factor polymorphism (BDNF Val66Met) and suicide. J. Affect. Disord. 2011, 128, 287–290. [Google Scholar] [CrossRef]
- Pivac, N.; Pregelj, P.; Nikolac, M.; Zupanc, T.; Nedic, G.; Muck-Seler, D.; Paska, V.A. The association between catechol-O-methyl-transferase Val108/158Met polymorphism and suicide. Genes Brain Behav. 2011, 10, 565–569. [Google Scholar] [CrossRef]
- Nedic, G.; Nikolac, M.; Sviglin, K.N.; Muck-Seler, D.; Borovecki, F.; Pivac, N. Association study of a functional catechol- O-methyltransferase (COMT) Val108/158Met polymorphism and suicide attempts in patients with alcohol dependence. Int. J. Neuropsychopharmacol. 2011, 14, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Zhernakova, D.V.; Brukhin, V.; Malov, S.; Oleksyk, T.K.; Koepfli, K.P.; Zhuk, A.; Dobrynin, P.; Kliver, S.; Cherkasov, N.; Tamazian, G.; et al. Genome-wide sequence analyses of ethnic populations across Russia. Genomics 2020, 112, 442–458. [Google Scholar] [CrossRef]
- Bondarenko, E.; Shadrina, M.; Grishkina, M.; Druzhkova, T.; Akzhigitov, R.; Gulyaeva, N.; Guekht, A.; Slominsky, P. Genetic Analysis of BDNF, GNB3, MTHFR, ACE and APOE Variants in Major and Recurrent Depressive Disorders in Russia. Int. J. Med. Sci. 2016, 13, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Butovskaya, M.L.; Butovskaya, P.R.; Vasilyev, V.A.; Sukhodolskaya, J.M.; Fekhredtinova, D.I.; Karelin, D.; Fedenok, J.; Mabulla, A.Z.P.; Ryskov, A.P.; Lazebny, O.E. Serotonergic gene polymorphisms (5-HTTLPR, 5HTR1A, 5HTR2A), and population differences in aggression: Traditional (Hadza and Datoga) and industrial (Russians) populations compared. J. Physiol. Anthr. 2018, 37, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekhretdinova, D.I.; Sukhodolskaya, E.M.; Shibalev, D.V.; Lazebnyy, O.E.; Butovskaya, M.L.; Ryskov, A.P.; Vasil’Yev, V.A. Polymorphism of the Two Genes Encoding Catecholamine Degradation Enzymes (COMT and MAOA) in the Hadza and Datoga African Ethnic Populations. Mol. Genet. Microbiol. Virol. 2018, 33, 195–200. [Google Scholar] [CrossRef]
- Sukhodolskaya, E.M.; Fehretdinova, D.I.; Shibalev, D.V.; Lazebny, O.E.; Mabulla, A.Z.P.; Butovskaya, M.L.; Ryskov, A.P.; Vasilyev, V.V. Polymorphisms of dopamine receptor genesDRD2andDRD4in African populations of Hadza and Datoga differing in the level of culturally permitted aggression. Ann. Hum. Genet. 2018, 82, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Sukhodolskaya, E.M.; Vasilyev, V.A.; Shibalev, D.V.; Shcherbakova, O.I.; Kulikov, A.M.; Lazebny, O.E.; Karelin, D.; Butovskaya, M.L.; Ryskov, A.P. Comparative analysis of polymorphisms of the serotonin receptor genes HTR1A, HTR2A, and HTR1B in Hadza and Datoga males. Russ. J. Genet. 2015, 51, 1129–1134. [Google Scholar] [CrossRef]
- Sukhodol’Skaia, E.M.; Vasil’Ev, V.; Shibalev, D.V.; Shcherbakova, O.; Kulikov, A.M.; Lazebnyĭ, O.; A Dronova, D.; Butovskaia, M.L.; Ryskov, A.P. The 3′-UTR polymorphism of dopamine transporter gene in hadza and datoga males. Moлeкyляpнaя биoлoгия 2015, 48, 295–299. [Google Scholar] [CrossRef]


{kind=link}
| Age, Years | BDI | HAMD | SA | PA | |
|---|---|---|---|---|---|
| min | 18 | 8 | 7 | 25 | 25 |
| max | 77 | 53 | 35 | 80 | 80 |
| mean, SD | 31.54 ± 11.13 | 28.85 ± 9.58 | 21.19 ± 5.22 | 55.52 ± 12.93 | 56.44 ± 10.05 |
| Genotype | Control, n (%) | Suicide, n (%) | Logistic Regression, Genetic Model; LRT, P | ORs, P, 95% CI |
|---|---|---|---|---|
| SLC6A3 (DAT1) 40 bp VNTR | ||||
| LL | 113 (69.3) | 52 (52) | codominant: 10.16, 0.006 * dominant: 10.54, 0.001 ** overdominant: 8.14, 0.004 ** log-additive: 8.05, 0.004 ** | 0.48, 0.005 *, 0.29–0.82 |
| LS | 46 (28.2) | 44 (44) | 1.998, 0.009 *, 1.19–3.37 | |
| SS | 4 (2.5) | 4 (4) | 1.66, 0.483, 0.4–6.78 | |
| COMT rs4680 | ||||
| AA | 48 (29.4) | 17 (17) | codominant: 6.42, 0.04 * dominant: 6.11, 0.013 * log-additive: 5.39, 0.02 * | 0.49, 0.025 *, 0.26–0.91 |
| GA | 78 (47.9) | 53 (53) | 1.23, 0.418, 0.75–2.02 | |
| GG | 37 (22.7) | 30 (30) | 1.46, 0.188, 0.83–2.56 | |
| SLC6A4 5-HTTLPR + rs25531 | ||||
| high/high | 49 (31.2) | 35 (35) | recessive: 5.06, 0.024 * | 1.19, 0.528, 0.7–2.02 |
| high/low | 67 (42.7) | 50 (50) | 1.34, 0.251, 0.81–2.22 | |
| low/low | 41 (26.1) | 15 (15) | 0.5, 0.038, 0.26–0.96 | |
| BDNF rs6264 | ||||
| CC | 183 (70.7) | 61 (61) | recessive: 2.82, 0.093+ log-additive: 2.86, 0.09+ | 0.65, 0.08 +, 0.4–1.05 |
| CT | 69 (26.6) | 32 (32) | 1.3, 0.312, 0.78–2.14 | |
| TT | 7 (2.7) | 7 (7) | 2.71, 0.069 +, 0.93–7.93 | |
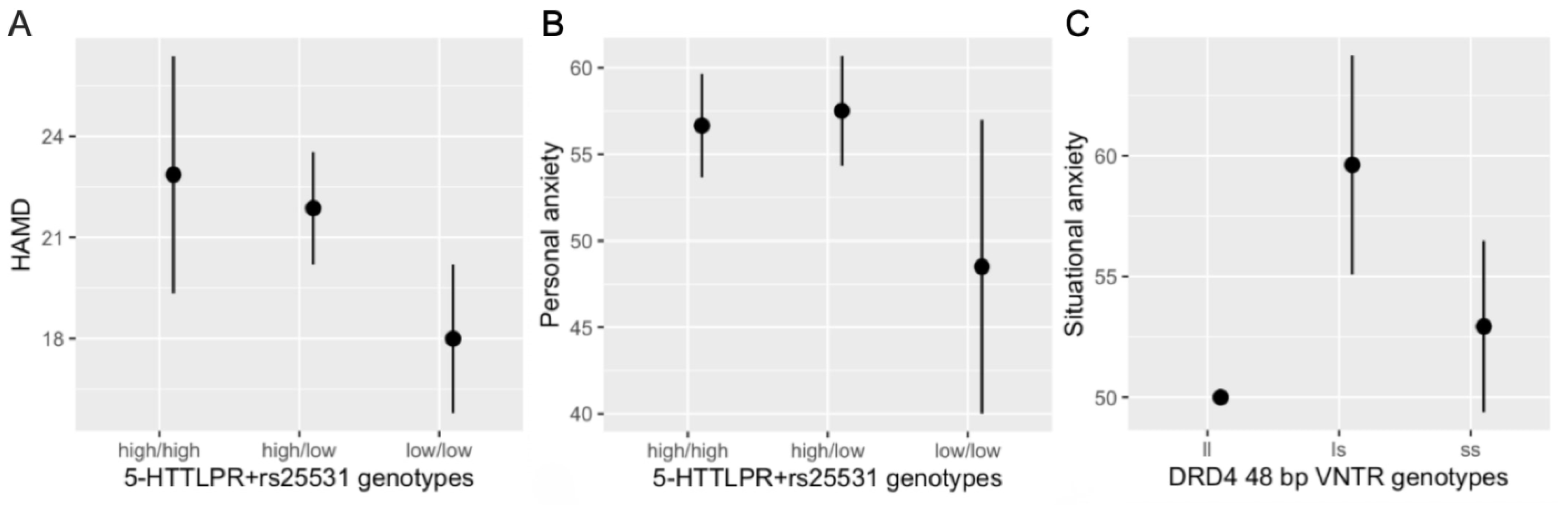
| Scale | Gene | Polymorphism | LRT | P |
|---|---|---|---|---|
| HAMD | SLC6A4 (5HTT) | 5-HTTLPR + rs25531 | 6.29 | 0.044 * |
| Personal anxiety | SLC6A4 (5HTT) | 5-HTTLPR + rs25531 | 7.09 | 0.029 * |
| HTR1B | rs6296 | 5.03 | 0.081 + | |
| Situational anxiety | DRD4 | 48 bp VNTR | 6.46 | 0.039 * |
| SLC6A4 (5HTT) | 5-HTTLPR + rs25531 | 3.85 | 0.146 | |
| HTR1A | rs6295 | 4.26 | 0.119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafikova, E.; Shadrina, M.; Slominsky, P.; Guekht, A.; Ryskov, A.; Shibalev, D.; Vasilyev, V. SLC6A3 (DAT1) as a Novel Candidate Biomarker Gene for Suicidal Behavior. Genes 2021, 12, 861. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060861
Rafikova E, Shadrina M, Slominsky P, Guekht A, Ryskov A, Shibalev D, Vasilyev V. SLC6A3 (DAT1) as a Novel Candidate Biomarker Gene for Suicidal Behavior. Genes. 2021; 12(6):861. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060861
Chicago/Turabian StyleRafikova, Ekaterina, Maria Shadrina, Peter Slominsky, Alla Guekht, Alexey Ryskov, Dmitry Shibalev, and Vasiliy Vasilyev. 2021. "SLC6A3 (DAT1) as a Novel Candidate Biomarker Gene for Suicidal Behavior" Genes 12, no. 6: 861. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060861