
Proteomics Analysis Reveals Diverse Molecular Characteristics between Endocardial and Aortic-Valvular Endothelium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Endothelial Cell Culture and Protein Isolation
2.2. Liquid Chromatography and Mass Spectrometry
2.3. Data Analysis of Proteomics Data
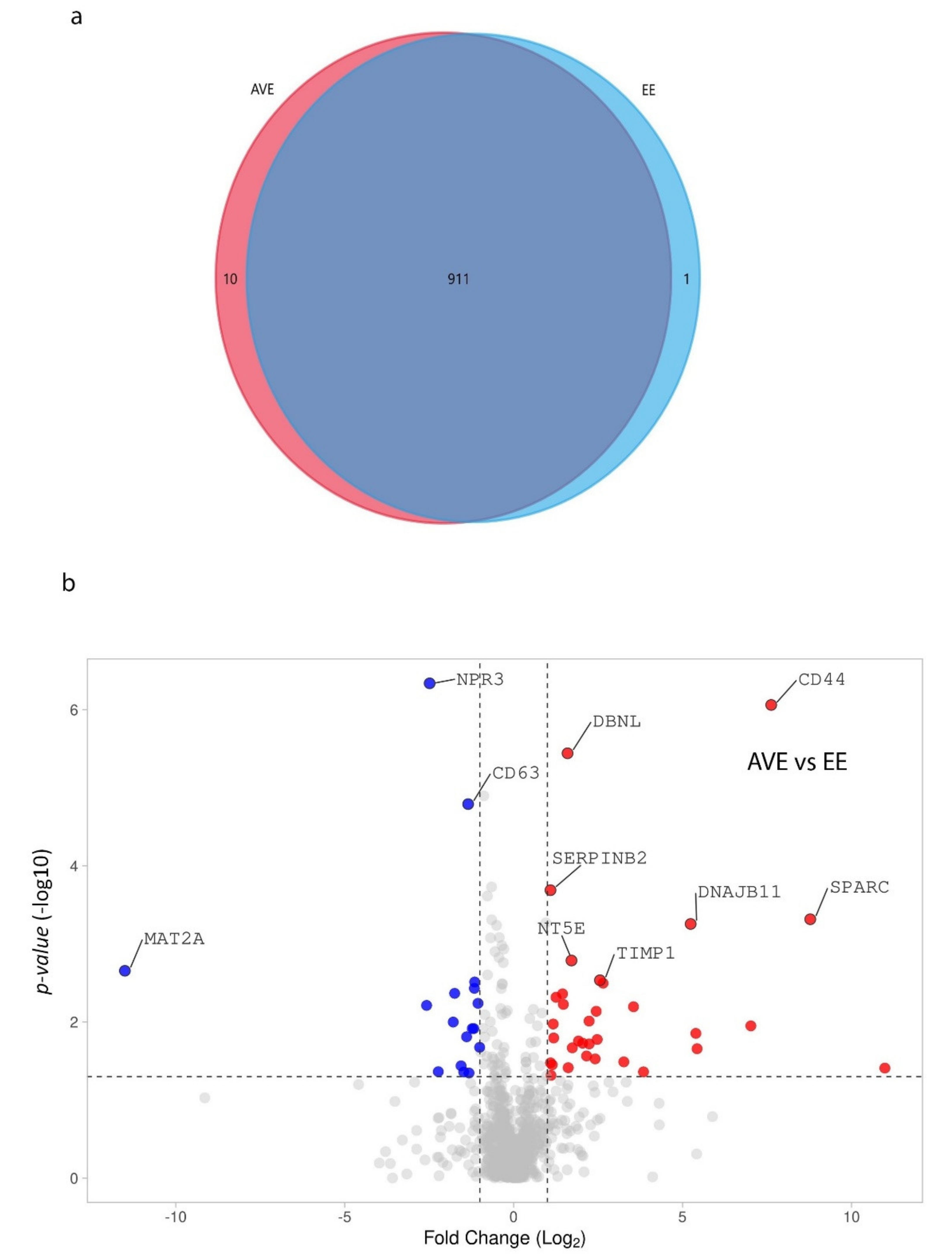
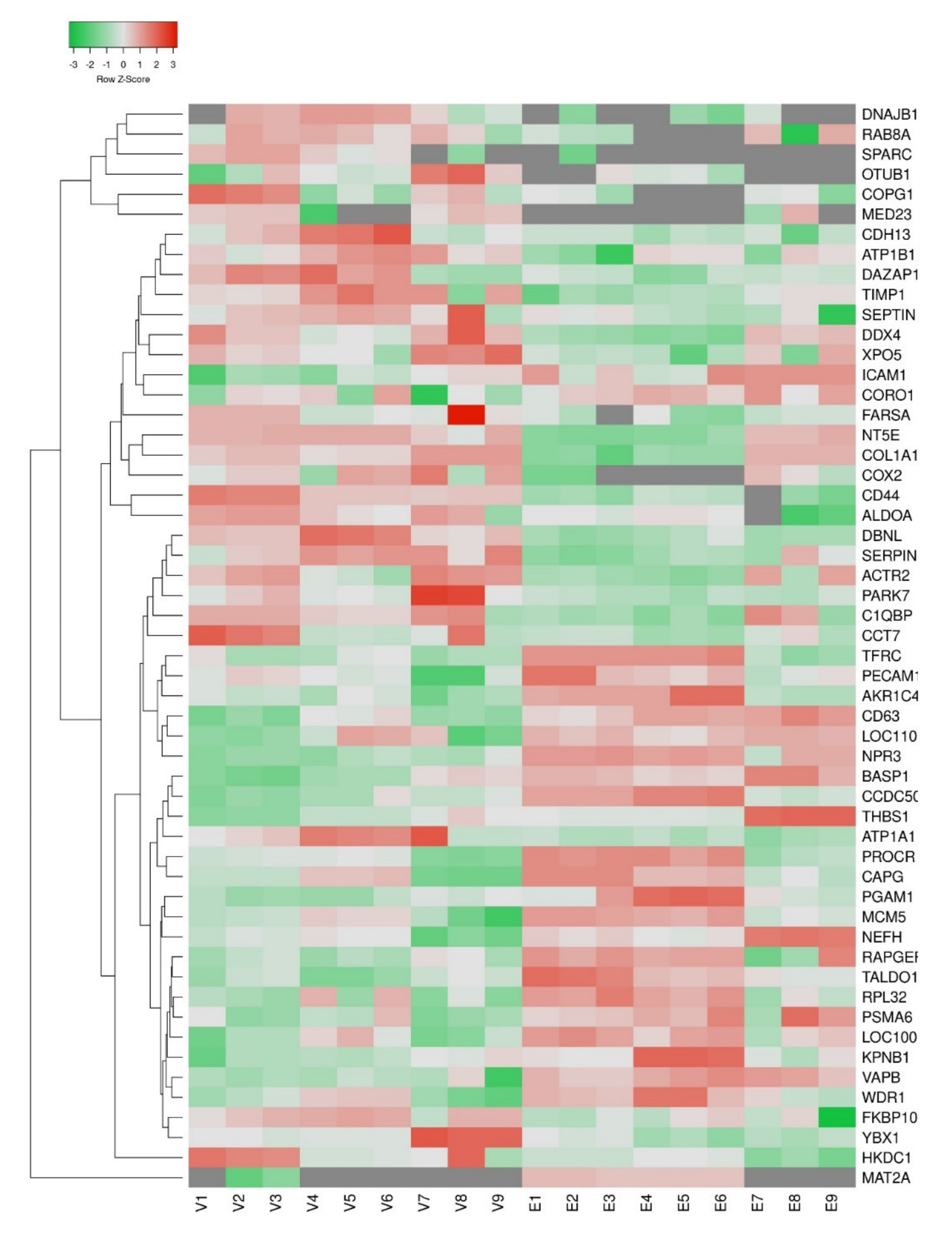
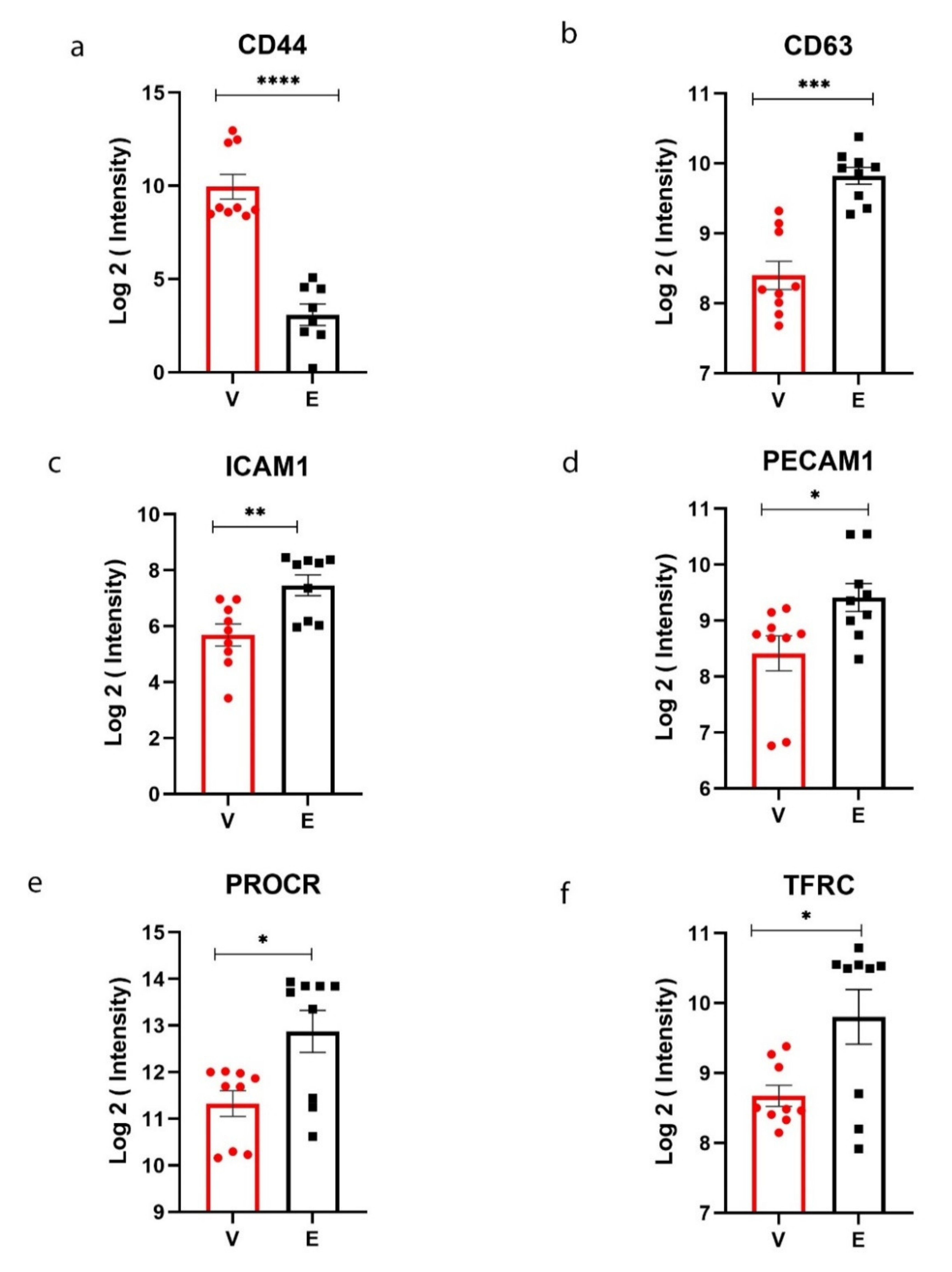
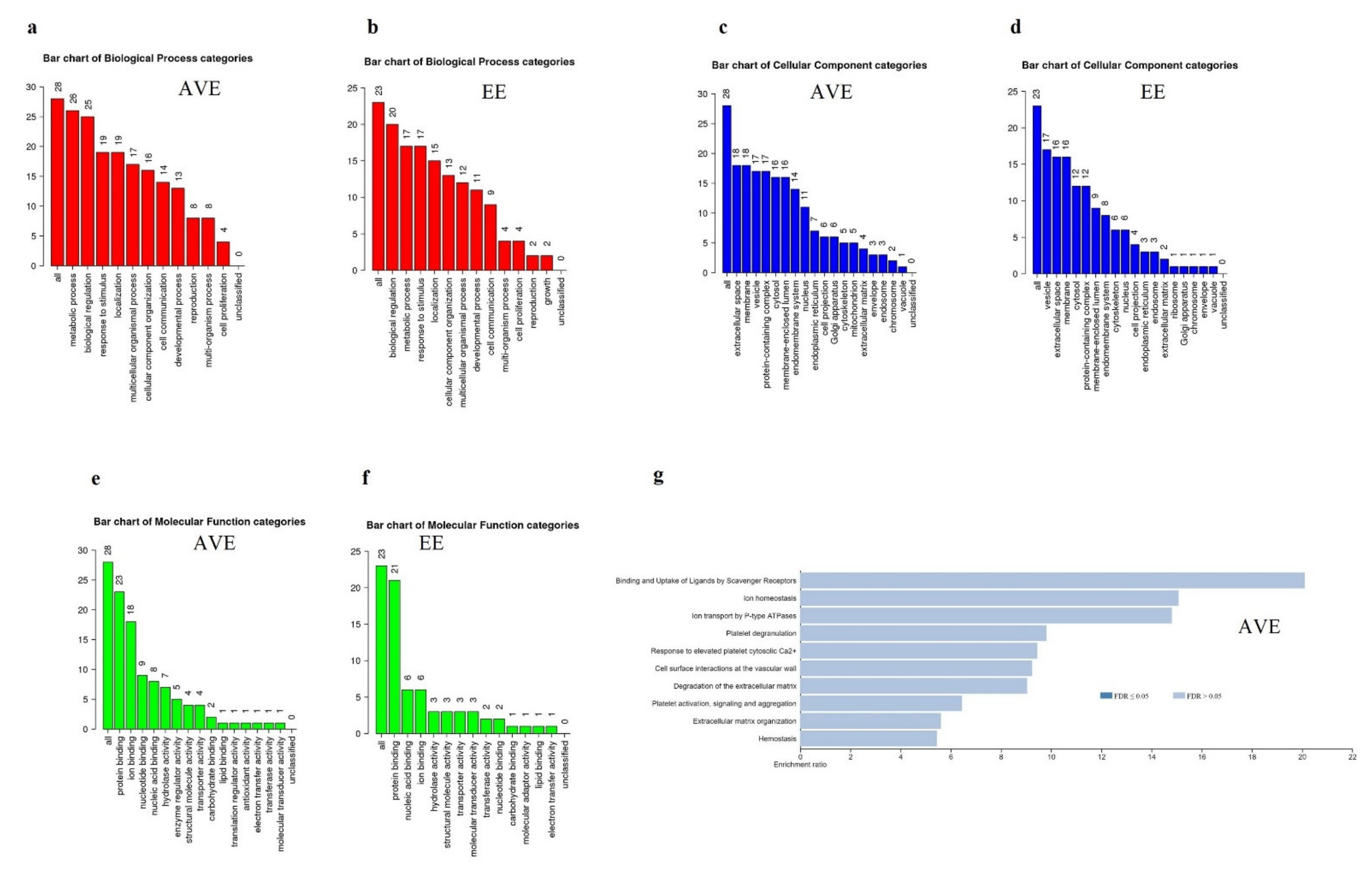
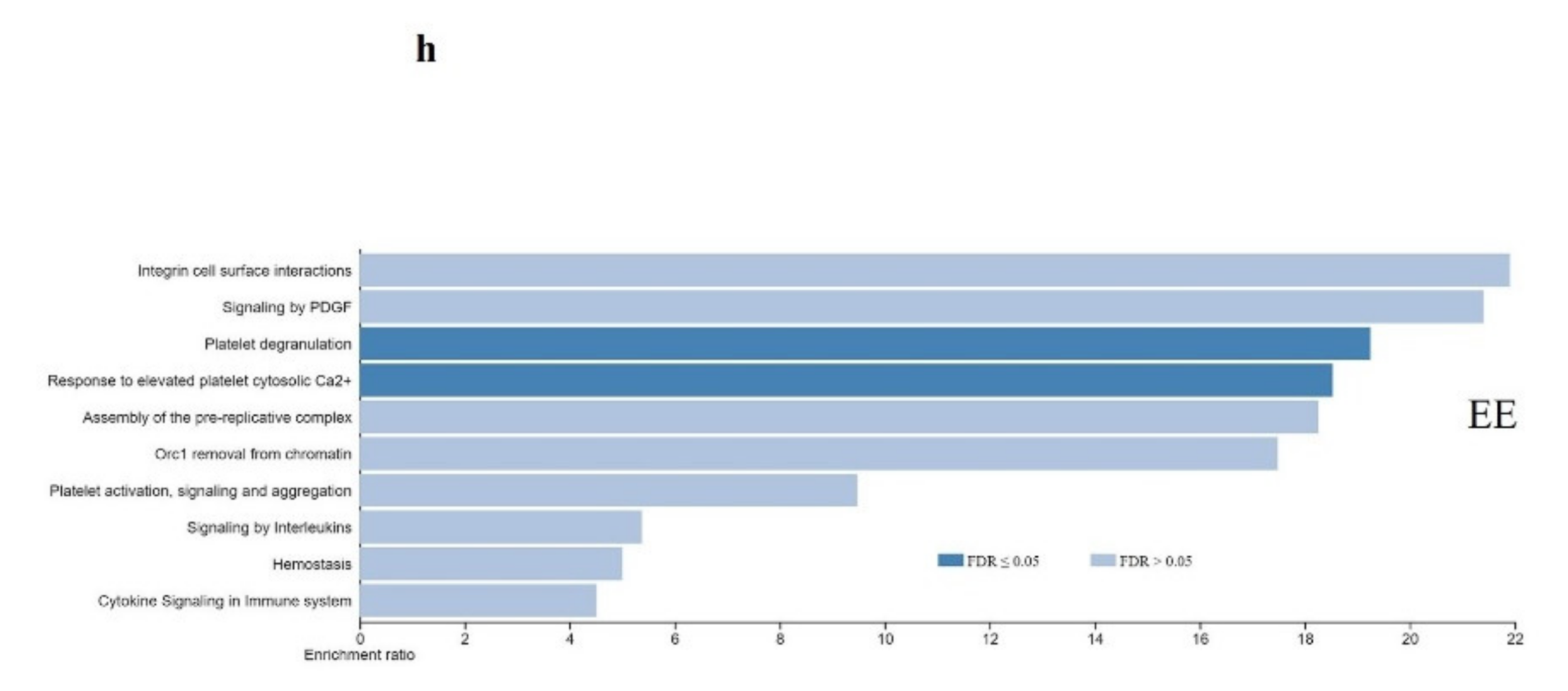
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arciniegas, E.; Ponce, L.; Hartt, Y.; Graterol, A.; Carlini, R.G. Intimal thickening involves transdifferentiation of embryonic endothelial cells. Anat. Rec. 2000, 258, 47–57. [Google Scholar] [CrossRef]
- Leask, R.L.; Jain, N.; Butany, J. Endothelium and valvular diseases of the heart. Microsc. Res. Tech. 2003, 60, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.L. Staphylococcal infective endocarditis. Mayo Clin. Proc. 1982, 57, 106–114. [Google Scholar] [PubMed]
- Tandon, R.; Sharma, M.; Chandrashekhar, Y.; Kotb, M.; Yacoub, M.H.; Narula, J. Revisiting the pathogenesis of rheumatic fever and carditis. Nat. Rev. Cardiol. 2013, 10, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Prasad, Y.; Bhalodkar, N.C. Aortic sclerosis—A marker of coronary atherosclerosis. Clin. Cardiol. 2004, 27, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Butcher, J.T.; Tressel, S.; Johnson, T.; Turner, D.; Sorescu, G.; Jo, H.; Nerem, R.M. Transcriptional profiles of valvular and vascular endothelial cells reveal phenotypic differences: Influence of shear stress. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Simmons, C.A.; Grant, G.R.; Manduchi, E.; Davies, P.F. Spatial heterogeneity of endothelial phenotypes correlates with side-specific vulnerability to calcification in normal porcine aortic valves. Circ. Res. 2005, 96, 792–799. [Google Scholar]
- Eisenberg, L.M.; Markwald, R.R. Molecular regulation of atrioventricular valvuloseptal morphogenesis. Circ. Res. 1995, 77, 1–6. [Google Scholar] [CrossRef]
- Kuruvilla, L.; Kartha, C.C. Molecular mechanisms in endothelial regulation of cardiac function. Mol. Cell Biochem. 2003, 253, 113–123. [Google Scholar] [CrossRef]
- Misfeldt, A.M.; Boyle, S.C.; Tompkins, K.L.; Bautch, V.L.; Labosky, P.A.; Baldwin, H.S. Endocardial cells are a distinct endothelial lineage derived from Flk1+ multipotent cardiovascular progenitors. Dev. Biol. 2009, 333, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Jaleel, A.; Kumar, A.A.; Kumar, G.S.A.; Surendran, A.; Kartha, C.C. Label-free quantitative proteomics analysis reveals distinct molecular characteristics in endocardial endothelium. Mol. Cell Biochem. 2019, 451, 1–10. [Google Scholar] [CrossRef]
- Smith, J.A.; Radomski, M.W.; Schulz, R.; Moncada, S.; Lewis, M.J. Porcine ventricular endocardial cells in culture express the inducible form of nitric oxide synthase. Br. J. Pharmacol. 1993, 108, 1107–1110. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Benito-Martin, A.; Peinado, H. FunRich proteomics software analysis, let the fun begin! Proteomics 2015, 15, 2555–2556. [Google Scholar] [CrossRef]
- Goedhart, J.; Luijsterburg, M.S. VolcaNoseR is a web app for creating, exploring, labeling and sharing volcano plots. Sci. Rep. 2020, 10, 1–5. [Google Scholar] [CrossRef]
- Wunsch, A.M.; Little, C.D.; Markwald, R.R. Cardiac endothelial heterogeneity defines valvular development as demonstrated by the diverse expression of JB3, an antigen of the endocardial cushion tissue. Dev. Biol. 1994, 165, 585–601. [Google Scholar] [CrossRef]
- Oppenheimer-Marks, N.; Davis, L.S.; Lipsky, P.E. Human T lymphocyte adhesion to endothelial cells and transendothelial migration. Alteration of receptor use relates to the activation status of both the T cell and the endothelial cell. J. Immunol. 1990, 145, 140–148. [Google Scholar]
- Okumura, N.; Hirano, H.; Numata, R.; Nakahara, M.; Ueno, M.; Hamuro, J.; Kinoshita, S.; Koizumi, N. Cell Surface Markers of Functional Phenotypic Corneal Endothelial Cells. Investig. Opthalmol. Vis. Sci. 2014, 55, 7610–7618. [Google Scholar] [CrossRef] [Green Version]
- Ponta, H.; Sherman, L.S.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.M.; Chen, Y.; Zheng, L.R.; Tao, Q.M.; Hu, S.J. [Effect of radiofrequency ablation on endothelial function and platelet activation]. Zhonghua Nei Ke Za Zhi 2003, 42, 400–402. [Google Scholar] [PubMed]
- Zibara, K.; Chettab, K.; McGregor, B.; Poston, R.; McGregor, J. Increased ICAM-1 and PECAM-1 transcription levels in the heart of Apo-E deficient mice in comparison to wild type (C57BL6). Thromb. Haemost. 2001, 85, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.A.; Baldwin, H.S.; DeLisser, H.; Mickanin, C.; Shen, H.M.; Kennedy, G.; Chen, A.; Edelman, J.M.; Albelda, S.M. Cell adhesion receptors and early mammalian heart development: An overview. C. R. Acad. Sci. Ser. III Sci. 1993, 316, 838–859. [Google Scholar]
- Cervero, J.; Montes, R.; Espana, F.; Esmon, C.T.; Hermida, J. Limited ability to activate protein C confers left atrial endocardium a thrombogenic phenotype: A role in cardioembolic stroke? Stroke 2011, 42, 2622–2624. [Google Scholar] [CrossRef]
- Schoner, A.; Tyrrell, C.; Wu, M.; Gelow, J.M.; Hayes, A.A.; Lindner, J.R.; Thornburg, K.L.; Hasan, W. Endocardial Endothelial Dysfunction Progressively Disrupts Initially Anti then Pro-Thrombotic Pathways in Heart Failure Mice. PLoS ONE 2015, 10, e0142940. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, W.A.; Brandon, M.R.; Hunt, S.V.; Williams, A.F.; Gatter, K.C.; Mason, D.Y. Transferrin receptor on endothelium of brain capillaries. Nat. Cell Biol. 1984, 312, 162–163. [Google Scholar] [CrossRef]
- Back, M.; Gasser, T.C.; Michel, J.B.; Caligiuri, G. Biomechanical factors in the biology of aortic wall and aortic valve diseases. Cardiovasc. Res. 2013, 99, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Tsang, H.G.; Clark, E.L.; Markby, G.R.; Bush, S.J.; Hume, D.A.; Corcoran, B.M.; MacRae, V.E.; Summers, K.M. Expression of Calcification and Extracellular Matrix Genes in the Cardiovascular System of the Healthy Domestic Sheep (Ovis aries). Front. Genet. 2020, 11, 919. [Google Scholar] [CrossRef]
- Gomez-Stallons, M.V.; Tretter, J.T.; Hassel, K.; Gonzalez-Ramos, O.; Amofa, D.; Ollberding, N.J.; Mazur, W.; Choo, J.K.; Smith, J.M.; Kereiakes, D.J.; et al. Calcification and extracellular matrix dysregulation in human postmortem and surgical aortic valves. Heart 2019, 105, 1616–1621. [Google Scholar] [CrossRef]
- Greene, C.L.; Jaatinen, K.J.; Wang, H.; Koyano, T.K.; Bilbao, M.S.; Woo, Y.J. Transcriptional Profiling of Normal, Stenotic, and Regurgitant Human Aortic Valves. Genes 2020, 11, 789. [Google Scholar] [CrossRef]
- Lerman, D.; Prasad, S.; Alotti, N. Calcific Aortic Valve Disease: Molecular Mechanisms and Therapeutic Approaches. Eur. Cardiol. Rev. 2015, 10, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Brutsaert, D.L. Cardiac Endothelial-Myocardial Signaling: Its Role in Cardiac Growth, Contractile Performance, and Rhythmicity. Physiol. Rev. 2003, 83, 59–115. [Google Scholar] [CrossRef]
- Noireaud, J.; Andriantsitohaina, R. Recent Insights in the Paracrine Modulation of Cardiomyocyte Contractility by Cardiac Endothelial Cells. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Boyce, S.; Lwaleed, B.A.; Kazmi, R.S. Homeostasis of Hemostasis: The Role of Endothelium. Semin. Thromb. Hemost. 2015, 41, 549–555. [Google Scholar] [CrossRef]
- Verhamme, P.; Hoylaerts, M.F. The Pivotal Role of the Endothelium in Haemostasis and Thrombosis. Acta Clin. Belg. 2006, 61, 213–219. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aneesh Kumar, A.; Ajith Kumar, G.S.; Satheesh, G.; Surendran, A.; Chandran, M.; Kartha, C.C.; Jaleel, A. Proteomics Analysis Reveals Diverse Molecular Characteristics between Endocardial and Aortic-Valvular Endothelium. Genes 2021, 12, 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071005
Aneesh Kumar A, Ajith Kumar GS, Satheesh G, Surendran A, Chandran M, Kartha CC, Jaleel A. Proteomics Analysis Reveals Diverse Molecular Characteristics between Endocardial and Aortic-Valvular Endothelium. Genes. 2021; 12(7):1005. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071005
Chicago/Turabian StyleAneesh Kumar, A., G. S. Ajith Kumar, Gopika Satheesh, Arun Surendran, Mahesh Chandran, Chandrasekharan C. Kartha, and Abdul Jaleel. 2021. "Proteomics Analysis Reveals Diverse Molecular Characteristics between Endocardial and Aortic-Valvular Endothelium" Genes 12, no. 7: 1005. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071005