
Extracellular Matrix-Related Hubs Genes Have Adverse Effects on Gastric Adenocarcinoma Prognosis Based on Bioinformatics Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microarray Data Collection
2.2. Data Processing
2.3. Identification of Coexpression Modules
2.4. Functional and Pathway Enrichment Analysis
2.5. PPI Network Construction and Analysis
2.6. The Expression and Survival Analysis of Hub Genes
3. Results
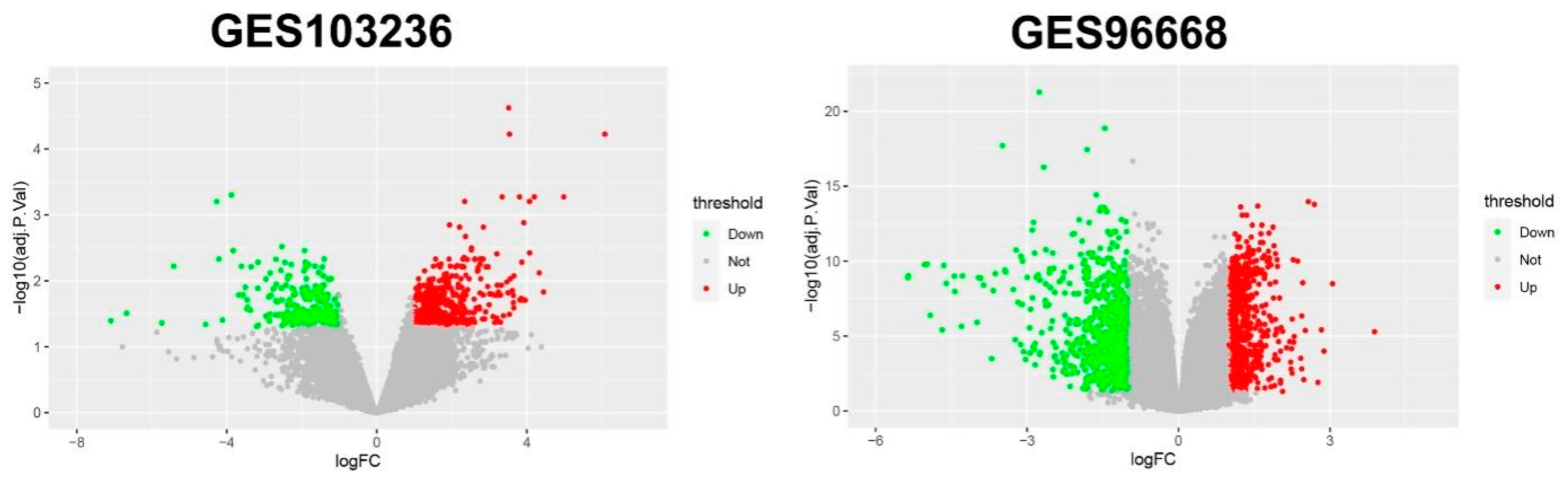
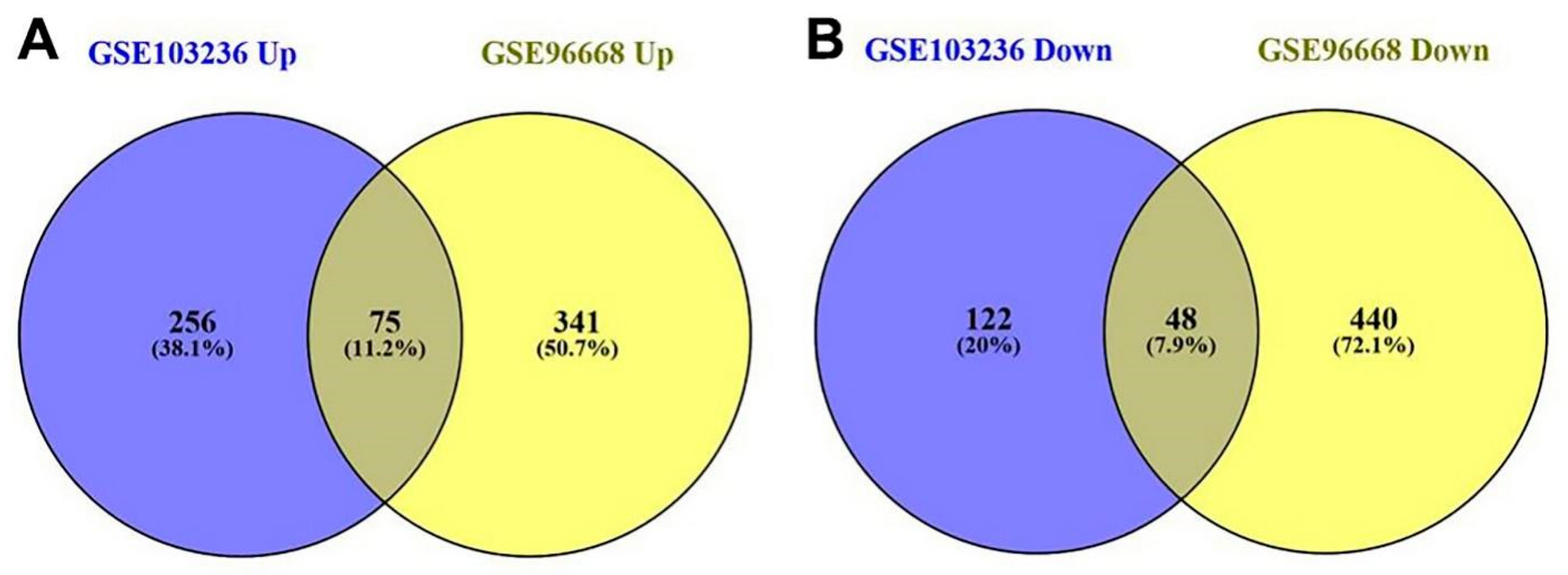
3.1. DEGs and Clusters
3.2. Functional and Pathway Enrichment Analyses
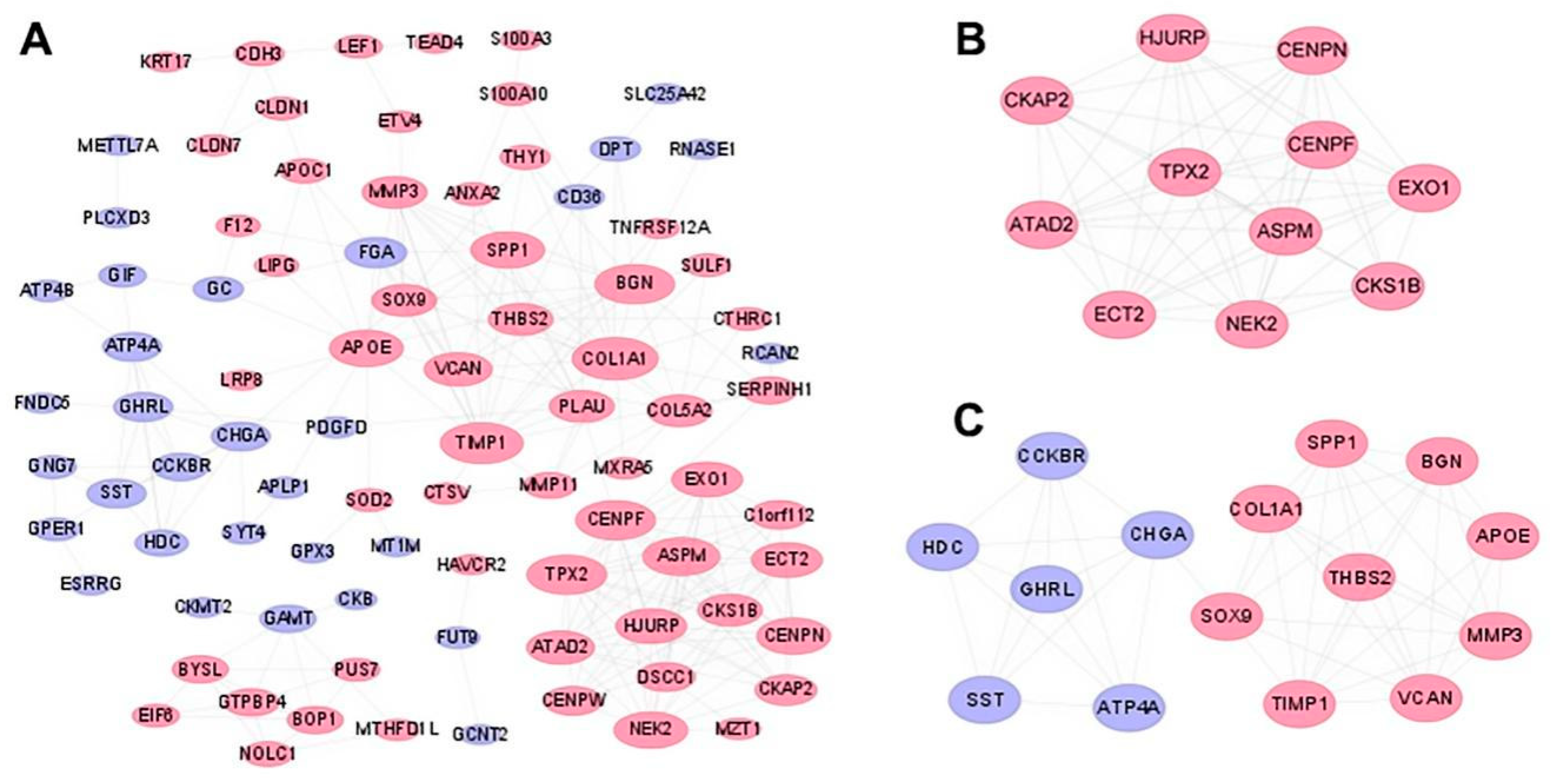
3.3. PPI Network and Module Analyses
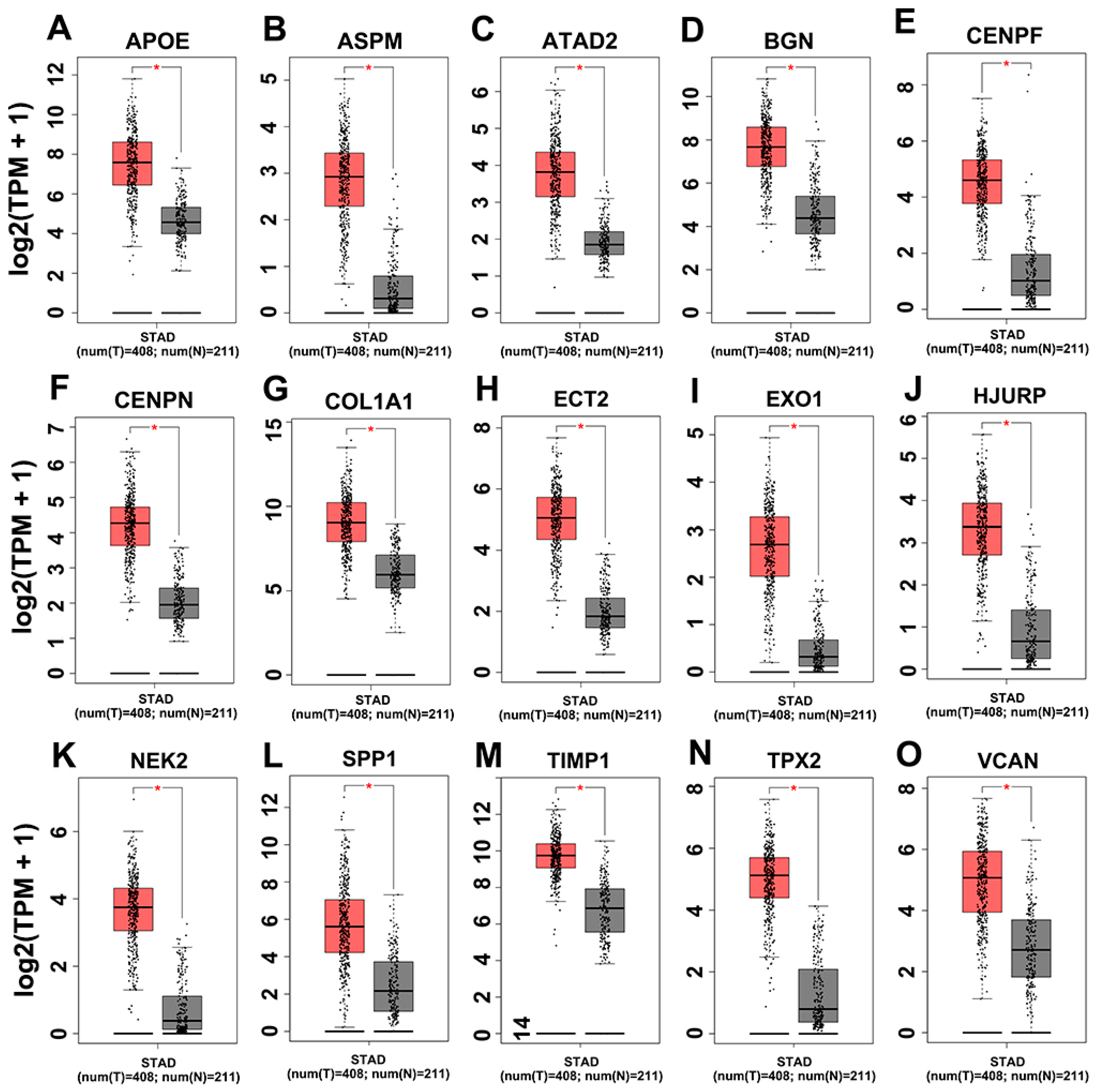
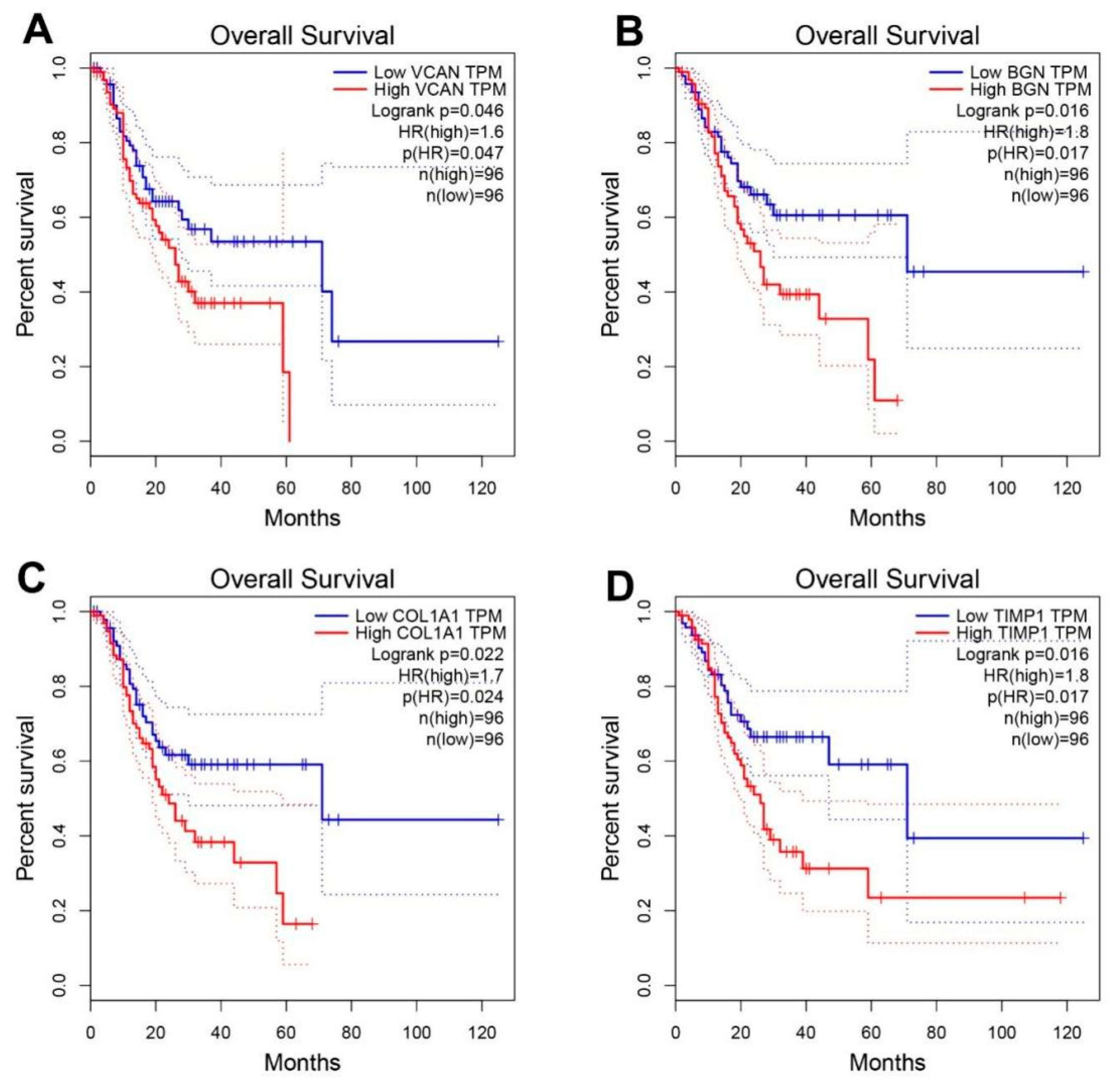
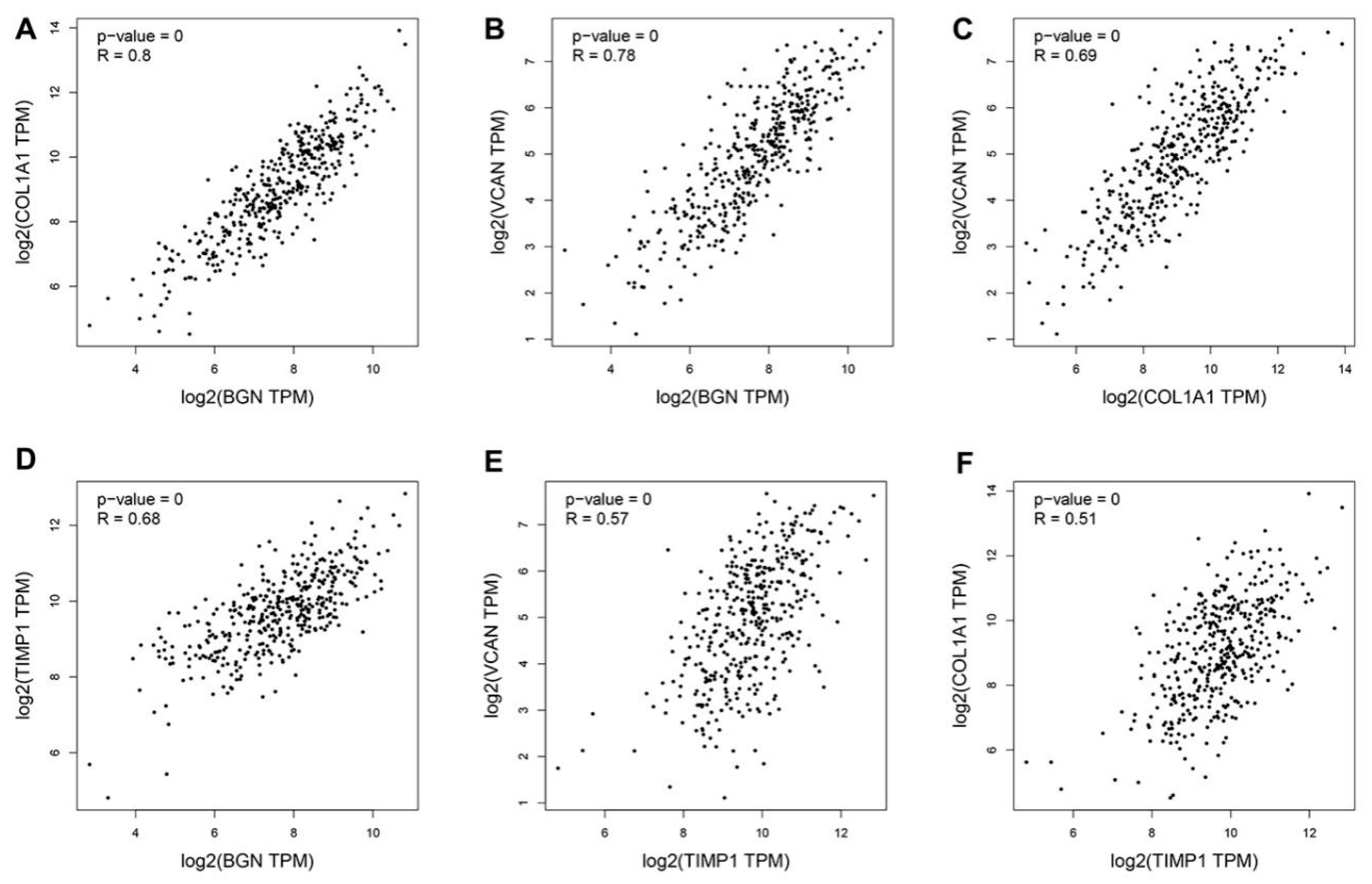
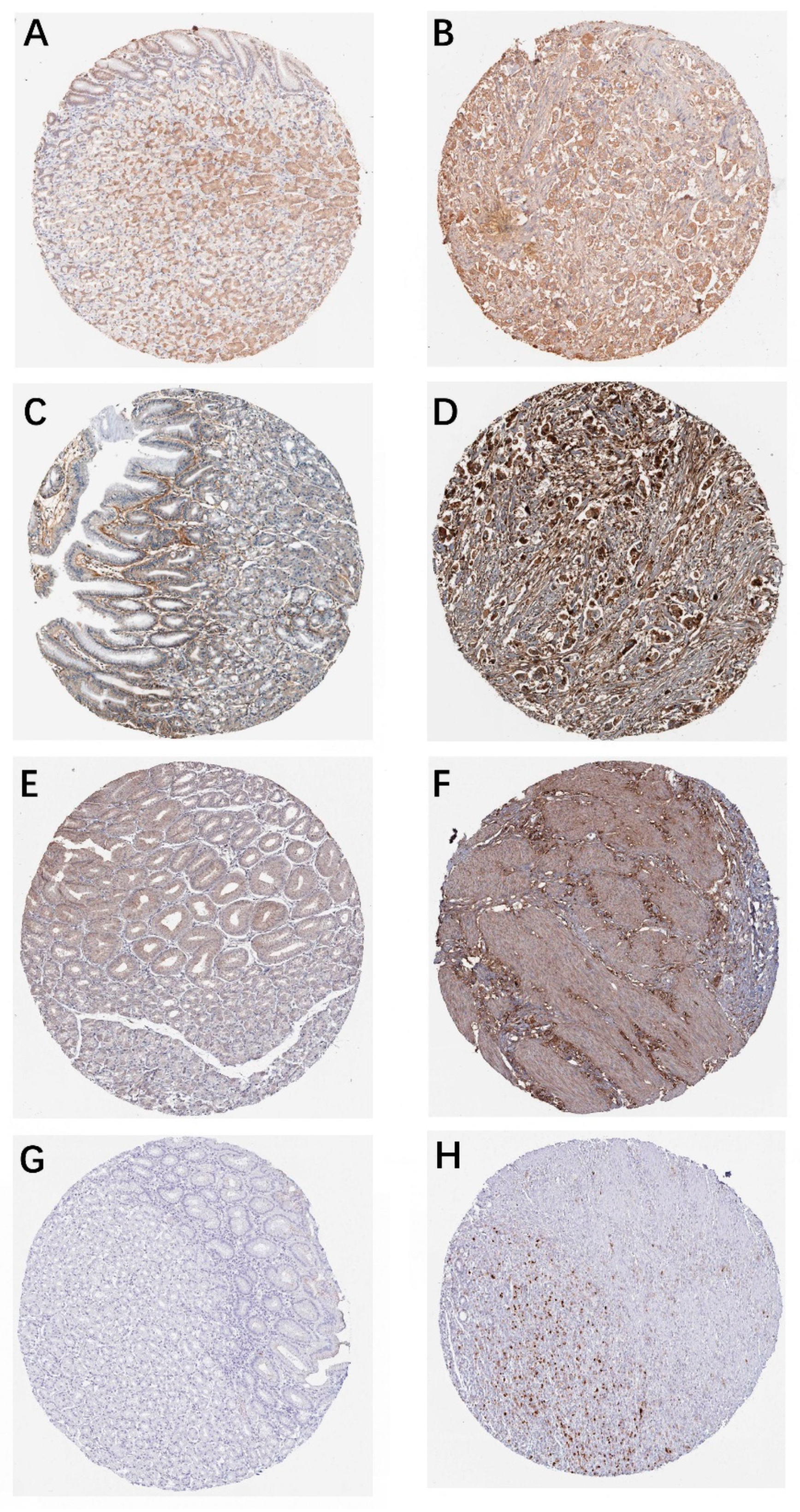
3.4. Hub Genes Survival Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Rugge, M.; Genta, R.M.; Di Mario, F.; El-Omar, E.M.; El-Serag, H.B.; Fassan, M.; Hunt, R.H.; Kuipers, E.J.; Malfertheiner, P.; Sugano, K.; et al. Gastric Cancer as Preventable Disease. Clin. Gastroenterol. Hepatol. 2017, 15, 1833–1843. [Google Scholar] [CrossRef] [Green Version]
- Bass, A.J.; Thorsson, V.; Shmulevich, I.; Reynolds, S.M.; Miller, M.; Bernard, B.; Hinoue, T.; Laird, P.W.; Curtis, C.; Shen, H.; et al. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar]
- Ku, G.Y. Next generation sequencing in gastric or gastroesophageal adenocarcinoma. Transl. Gastroenterol. Hepatol. 2020, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Gao, X.Y.; Peng, X.X.; Chen, M.J.M.; Li, Z.; Wei, B.; Wen, X.Z.; Wei, B.Y.; Dong, Y.; Bu, Z.D.; et al. Multi-omics characterization of molecular features of gastric cancer correlated with response to neoadjuvant chemotherapy. Sci. Adv. 2020, 6, eaay4211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, D.Q.; Liao, W.J.; Kim, K.M.; Shi, M.; Zhou, R.; Yu, Y.F.; Ye, Z.L.; Wu, J.N. Tumor microenvironment evaluation and tumor intrinsic genomic features predict anti-PD-1 response of metastatic gastric cancer: Results from phase II clinical trial and multi-omics data. Cancer Res. 2020, 80 (Suppl. 16), 6655. [Google Scholar]
- Liu, N.N.; Wu, Y.; Cheng, W.P.; Wu, Y.X.; Wang, L.G.; Zhuang, L.W. Identification of novel prognostic biomarkers by integrating multi-omics data in gastric cancer. BMC. Cancer 2021, 21, 460. [Google Scholar] [CrossRef]
- Yoon, S.J.; Kim, J.Y.; Long, N.P.; Min, J.E.; Kim, H.M.; Yoon, J.H.; Anh, N.H.; Park, M.C.; Kwon, S.W.; Lee, S.K. Comprehensive Multi-Omics Analysis Reveals Aberrant Metabolism of Epstein-Barr-Virus-Associated Gastric Carcinoma. Cells 2019, 8, 1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Y.; Cai, H.; Zheng, W.C.; Tong, M.S.; Li, H.D.; Ao, L.; Li, J.; Hong, G.N.; Li, M.Y.; Guan, Q.Z.; et al. An individualized prognostic signature for gastric cancer patients treated with 5-Fluorouracil-based chemotherapy and distinct multi-omics characteristics of prognostic groups. Oncotarget 2016, 7, 8743–8755. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, J.; Wang, C.; Xia, L.; Lu, W. Microenvironment remodeled by tumor and stromal cells elevates fibroblast-derived COL1A1 and facilitates ovarian cancer metastasis. Exp. Cell Res. 2020, 112153. [Google Scholar] [CrossRef]
- Rabelink, T.J.; van den Berg, B.M.; Garsen, M.; Wang, G.; Elkin, M.; van der Vlag, J. Heparanase: Roles in cell survival, extracellular matrix remodelling and the development of kidney disease. Nat. Rev. Nephrol. 2017, 13, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Chivu, E.M.; Necula, L.G.; Dragu, D.; Badea, L.; Dima, S.O.; Tudor, S.; Nastase, A.; Popescu, I.; Diaconu, C.C. Identification of potential biomarkers for early and advanced gastric adenocarcinoma detection. Hepatogastroenterology 2010, 57, 1453–1464. [Google Scholar]
- Bornschein, J.; Wernisch, L.; Secrier, M.; Miremadi, A.; Perner, J.; MacRae, S.; O’Donovan, M.; Newton, R.; Menon, S.; Bower, L.; et al. Transcriptomic profiling reveals three molecular phenotypes of adenocarcinoma at the gastroesophageal junction. Int. J. Cancer 2019, 145, 3389–3401. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets-update. Nucleic. Acids. Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.F.; Li, C.W.; Kang, B.X.; Gao, G.; Li, C.; Zhang, Z.M. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic. Acids. Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer Statistics in China, 2015. Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Schurch, C.M.; Bhate, S.S.; Barlow, G.L.; Phillips, D.J.; Noti, L.; Zlobec, I.; Chu, P.; Black, S.; Demeter, J.; McIlwain, D.R. Coordinated cellular neighborhoods orchestrate antitumoral immunity at the colorectal cancer invasive front. Cell 2020, 183, 838. [Google Scholar] [CrossRef]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, L.W.; Heegaard, A.M.; Vetter, U.; Vogel, W.; Just, W.; Termine, J.D.; Young, M.F. Human biglycan gene. Putative promoter, intron-exon junctions, and chromosomal localization. J. Biol. Chem. 1991, 266, 14371–14377. [Google Scholar] [CrossRef]
- Liu, Y.; Li, W.; Li, X.; Tai, Y.; Lu, Q.; Yang, N.; Jiang, J. Expression and significance of biglycan in endometrial cancer. Arch. Gynecol. Obstet. 2014, 289, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Aprile, G.; Avellini, C.; Reni, M.; Mazzer, M.; Foltran, L.; Rossi, D.; Cereda, S.; Iaiza, E.; Fasola, G.; Piga, A. Biglycan expression and clinical outcome in patients with pancreatic adenocarcinoma. Tumor Biol. 2013, 34, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ohga, N.; Hida, Y.; Maishi, N.; Kawamoto, T.; Kitayama, K.; Akiyama, K.; Osawa, T.; Kondoh, M.; Matsuda, K.; et al. Biglycan is a specific marker and an autocrine angiogenic factor of tumour endothelial cells. Brit. J. Cancer 2012, 106, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Duan, Y.T.; Li, J.F.; Su, L.P.; Yan, M.; Zhu, Z.G.; Liu, B.; Yang, Q. Biglycan enhances gastric cancer invasion by activating FAK signaling pathway. Oncotarget 2014, 5, 1885–1896. [Google Scholar] [CrossRef] [Green Version]
- Said, N.; Theodorescu, D. RhoGI2 suppresses bladder cancer metastasis via reduction of inflammation in the tumor microenvironment. Oncoimmunology 2012, 1, 1175–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogels, M.; Braster, R.; Nijland, P.G.; Gul, N.; van de Luijtgaarden, W.; Fijneman, R.J.A.; Meijer, G.A.; Jimenez, C.R.; Beelen, R.H.J.; van Egmond, M. Carcinoma origin dictates differential skewing of monocyte function. Oncoimmunology 2012, 1, 798–809. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, X.; Wu, J.; Quan, W.; Ma, L.; Yang, F.; Wu, K.; Wan, H. Tumor-produced versican V1 enhances hCAP18/LL-37 expression in macrophages through activation of TLR2 and vitamin D3 signaling to promote ovarian cancer progression in vitro. PLoS ONE 2013, 8, e566162. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Huang, W.; Tian, D.; Zhang, L.; Qi, X.; Chen, Z.; Shang, X.; Nie, Y.; Wu, K. Forkhead box Q1 promotes hepatocellular carcinoma metastasis by transactivating ZEB2 and versicanV1 expression. Hepatology 2014, 59, 958–973. [Google Scholar] [CrossRef]
- Pukkila, M.J.; Kosunen, A.S.T.; Virtaniemi, J.A.; Kumpulainen, E.J.; Johansson, R.T.; Kellokoski, J.K.; Nuutinen, J.; Kosma, V. Versican expression in pharyngeal squamous cell carcinoma: An immunohistochemical study. J. Clin. Pathol. 2004, 57, 735–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, K.; Nelson, C.M.; Nandadasa, S.; Aramaki-Hattori, N.; Lindner, D.J.; Alban, T.; Inagaki, J.; Ohtsuki, T.; Oohashi, T.; Apte, S.S.; et al. Stromal Versican Regulates Tumor Growth by Promoting Angiogenesis. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gilkes, D.M.; Semenza, G.L.; Wirtz, D. Hypoxia and the extracellular matrix: Drivers of tumour metastasis. Nat. Rev. Cancer 2014, 14, 430–439. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, C.M.; Selvarajan, S.; Mudgett, J.; Werb, Z. Stromelysin-1 regulates adipogenesis during mammary gland involution. J. Cell Biol. 2001, 152, 693–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, F.; Teixeira, A.L.; Nogueira, I.; Morais, M.; Maia, J.; Bodo, C.; Ferreira, M.; Vieira, I.; Silva, J.; Lobo, J.; et al. Plasma extracellular vesicle-derived TIMP-1 mRNA as a prognostic biomarker in clear cell renal cell carcinoma: A pilot study. Int. J. Mol. Sci. 2020, 21, 462413. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEGs | List of Gene Symbols |
|---|---|
| Upregulated DEGs | ANXA2, APOC1, APOE, ASPM, ATAD2, AUNIP, BGN, BOP1, BYSL, C1orf112, CDH3, CEMIP, CENPF, CENPN, CENPW, CKAP2, CKS1B, CLDN1, CLDN7, COL1A1, COL5A2, CTHRC1, CTSV, DSCC1, ECT2, EIF6, ESM1, ETV4, EXO1, F12, FAM72D, FOXD2, GJB2, GTPBP4, HAVCR2, HJURP, HMGA1, IGF2BP3, KRT17, LEF1, LIPG, LRFN4, LRP8, MMP11, MMP3, MTHFD1L, MXRA5, MZT1, NEK2, NFE2L3, NOLC1, NPM3, PLAU, PMEPA1, PUS7, RIPK2, S100A10, S100A3, SEH1L, SERPINH1, SNX10, SOD2, SOX9, SPP1, SULF1, TEAD4, THBS2, THY1, TIMP1, TMEM158, TNFRSF12A, TPX2, UPP1, VCAN, ZFAS1 |
| Downregulated DEGs | ADHFE1, APLP1, APOBEC2, ARHGEF37, ATP4A, ATP4B, C16orf89, C2orf40, CCKBR, CD36, CHGA, CKB, CKMT2, DPT, ESRRG, FGA, FNDC5, FUT9, GAMT, GC, GCNT2, GHRL, GIF, GNG7, GPER1, GPX3, HDC, LIFR, MAL, METTL7A, MT1M, MYRIP, PDGFD, PLCXD3, PNPLA7, PPP2R3A, RCAN2, RERGL, RGN, RNASE1, RPRM, SIGLEC11, SLC25A4, SLC25A42, SLC2A12, SORBS2, SST, SYT4 |
| Category | Term | Count | p Value |
|---|---|---|---|
| Biological Processes | GO:0007155~cell adhesion | 13 | 7.92 × 10−5 |
| GO:0008284~positive regulation of cell proliferation | 10 | 5.02 × 10−3 | |
| GO:0043066~negative regulation of apoptotic process | 8 | 3.84 × 10−2 | |
| GO:0070374~positive regulation of ERK1 and ERK2 cascade | 7 | 1.35 × 10−3 | |
| GO:0030335~positive regulation of cell migration | 7 | 1.75 × 10−3 | |
| GO:0030198~extracellular matrix organization | 7 | 2.40 × 10−3 | |
| GO:0007067~mitotic nuclear division | 7 | 7.52 × 10−3 | |
| GO:0008283~cell proliferation | 7 | 4.15 × 10−2 | |
| GO:0030199~collagen fibril organization | 6 | 6.88 × 10−6 | |
| GO:0006629~lipid metabolic process | 6 | 4.68 × 10−3 | |
| Molecular Function | GO:0005515~protein binding | 69 | 4.98 × 10−2 |
| GO:0005509~calcium ion binding | 14 | 1.01 × 10−3 | |
| GO:0003682~chromatin binding | 7 | 4.77 × 10−2 | |
| GO:0042393~histone binding | 5 | 9.13 × 10−3 | |
| GO:0005201~extracellular matrix structural constituent | 4 | 1.03 × 10−2 | |
| GO:0008900~hydrogen:potassium-exchanging ATPase activity | 2 | 1.99 × 10−2 | |
| GO:0030284~estrogen receptor activity | 2 | 3.30 × 10−2 | |
| GO:0071813~lipoprotein particle binding | 2 | 3.30 × 10−2 | |
| GO:0004111~creatine kinase activity | 2 | 3.95 × 10−2 | |
| GO:0060228~phosphatidylcholine-sterol O-acyltransferase activator activity | 2 | 3.95 × 10−2 | |
| Cellular component | GO:0070062~extracellular exosome | 30 | 7.08 × 10−3 |
| GO:0005615~extracellular space | 27 | 4.46 × 10−7 | |
| GO:0005576~extracellular region | 27 | 1.20 × 10−5 | |
| GO:0048471~perinuclear region of cytoplasm | 12 | 2.49 × 10−3 | |
| GO:0005783~endoplasmic reticulum | 12 | 1.96 × 10−2 | |
| GO:0005730~nucleolus | 12 | 2.45 × 10−2 | |
| GO:0009986~cell surface | 10 | 9.16 × 10−3 | |
| GO:0031012~extracellular matrix | 9 | 7.28 × 10−4 | |
| GO:0005578~proteinaceous extracellular matrix | 8 | 1.90 × 10−3 | |
| GO:0045121~membrane raft | 7 | 2.34 × 10−3 | |
| GO:0005581~collagen trimer | 6 | 3.41 × 10−4 |
| Category | Term | Count | p Value |
|---|---|---|---|
| KEGG PATHWAY | hsa04971:Gastric acid secretion | 4 | 2.03 × 10−2 |
| hsa04512:ECM-receptor interaction | 5 | 4.98× 10−3 | |
| hsa04151:PI3K-Akt signaling pathway | 7 | 5.59 × 10−2 | |
| hsa00330:Arginine and proline metabolism | 3 | 6.02 × 10−2 | |
| hsa04510:Focal adhesion | 5 | 8.17 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alatan, H.; Chen, Y.; Zhou, J.; Wang, L. Extracellular Matrix-Related Hubs Genes Have Adverse Effects on Gastric Adenocarcinoma Prognosis Based on Bioinformatics Analysis. Genes 2021, 12, 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071104
Alatan H, Chen Y, Zhou J, Wang L. Extracellular Matrix-Related Hubs Genes Have Adverse Effects on Gastric Adenocarcinoma Prognosis Based on Bioinformatics Analysis. Genes. 2021; 12(7):1104. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071104
Chicago/Turabian StyleAlatan, Husile, Yinwei Chen, Jinghua Zhou, and Li Wang. 2021. "Extracellular Matrix-Related Hubs Genes Have Adverse Effects on Gastric Adenocarcinoma Prognosis Based on Bioinformatics Analysis" Genes 12, no. 7: 1104. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071104