
Mapping the Diverse and Inclusive Future of Parkinson’s Disease Genetics and Its Widespread Impact

 ,
,  and
and
Abstract
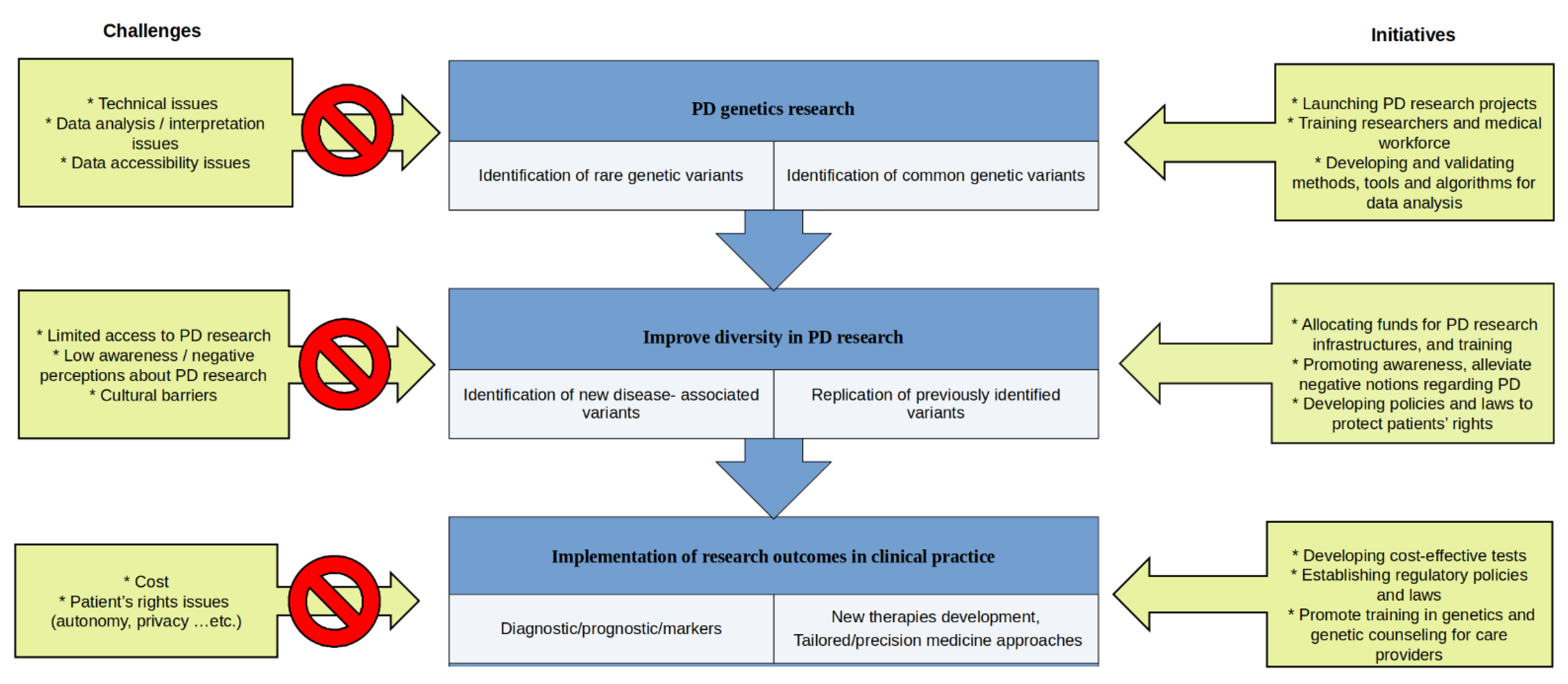
:1. Introduction
2. Parkinson’s Disease Genetics in Underrepresented Populations: Need for Inclusivity
3. Parkinson’s Disease Genetics in the Clinic: Interpretation of Genetic Testing and Genetic Counseling. Challenges and Limitations
4. The New Era of Parkinson’s Disease Genetics: Increasing Knowledge about Disease Etiology
5. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, C.B.; Bedenne, L. Management of Colorectal Polyps: Is All the Effort Worthwhile? J. Gastroenterol. Hepatol. 1990, 5, 144–165. [Google Scholar] [CrossRef]
- Nalls, M.A.; Blauwendraat, C.; Vallerga, C.L.; Heilbron, K.; Bandres-Ciga, S.; Chang, D.; Tan, M.; Kia, D.A.; Noyce, A.J.; Xue, A.; et al. Identification of Novel Risk Loci, Causal Insights, and Heritable Risk for Parkinson’s Disease: A Meta-Analysis of Genome-Wide Association Studies. Lancet Neurol. 2019, 18, 1091–1102. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Nalls, M.A.; Singleton, A.B. The Genetic Architecture of Parkinson’s Disease. Lancet Neurol. 2020, 19, 170–178. [Google Scholar] [CrossRef]
- Deng, H.; Wang, P.; Jankovic, J. The Genetics of Parkinson Disease. Ageing Res. Rev. 2018, 42, 72–85. [Google Scholar] [CrossRef]
- Klein, C.; Westenberger, A. Genetics of Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a008888. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.; Bandres-Ciga, S.; Diez-Fairen, M.; Quinn, J.P.; Billingsley, K.J. Genetic Risk Profiling in Parkinson’s Disease and Utilizing Genetics to Gain Insight into Disease-Related Biological Pathways. Int. J. Mol. Sci. 2020, 21, 7332. [Google Scholar] [CrossRef]
- Foo, J.N.; Chew, E.G.Y.; Chung, S.J.; Peng, R.; Blauwendraat, C.; Nalls, M.A.; Mok, K.Y.; Satake, W.; Toda, T.; Chao, Y.; et al. Identification of Risk Loci for Parkinson Disease in Asians and Comparison of Risk between Asians and Europeans: A Genome-Wide Association Study. JAMA Neurol. 2020, 77, 746–754. [Google Scholar] [CrossRef]
- Iwaki, H.; Blauwendraat, C.; Leonard, H.L.; Kim, J.J.; Liu, G.; Maple-Grødem, J.; Corvol, J.-C.; Pihlstrøm, L.; van Nimwegen, M.; Hutten, S.J.; et al. Genomewide Association Study of Parkinson’s Disease Clinical Biomarkers in 12 Longitudinal Patients’ Cohorts. Mov. Disord. 2019, 34, 1839–1850. [Google Scholar] [CrossRef]
- Blauwendraat, C.; Heilbron, K.; Vallerga, C.L.; Bandres-Ciga, S.; von Coelln, R.; Pihlstrøm, L.; Simón-Sánchez, J.; Schulte, C.; Sharma, M.; Krohn, L.; et al. Parkinson’s Disease Age at Onset Genome-Wide Association Study: Defining Heritability, Genetic Loci, and α-Synuclein Mechanisms. Mov. Disord. 2019, 34, 866–875. [Google Scholar] [CrossRef] [Green Version]
- Popejoy, A.B.; Fullerton, S.M. Genomics Is Failing on Diversity. Nature 2016, 538, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Sirugo, G.; Williams, S.M.; Tishkoff, S.A. The Missing Diversity in Human Genetic Studies. Cell 2019, 177, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Liu, Z.; Li, W.; Wang, Y.; Xie, A. Association between rs823128 Polymorphism and the Risk of Parkinson’s Disease: A Meta-Analysis. Neurosci. Lett. 2018, 665, 110–116. [Google Scholar] [CrossRef]
- Ibanez, L.; Dube, U.; Saef, B.; Budde, J.; Black, K.; Medvedeva, A.; Del-Aguila, J.L.; Davis, A.A.; Perlmutter, J.S.; Harari, O.; et al. Parkinson Disease Polygenic Risk Score Is Associated with Parkinson Disease Status and Age at Onset but Not with α-Synuclein Cerebrospinal Fluid Levels. BMC Neurol. 2017, 17, 198. [Google Scholar] [CrossRef]
- Tekola-Ayele, F.; Rotimi, C.N. Translational Genomics in Low- and Middle-Income Countries: Opportunities and Challenges. Public Health Genomics 2015, 18, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Sarfo, F.S.; Adamu, S.; Awuah, D.; Ovbiagele, B. Tele-Neurology in Sub-Saharan Africa: A Systematic Review of the Literature. J. Neurol. Sci. 2017, 380, 196–199. [Google Scholar] [CrossRef]
- Global Parkinson’s Genetics Program GP2: The Global Parkinson’s Genetics Program. Mov. Disord. 2021, 36, 842–851. [CrossRef]
- Rizig, M.; Okubadejo, N.; Salama, M.; Thomas, O.; Akpalu, A.; Gouider, R.; IPDGC Africa. The International Parkinson Disease Genomics Consortium Africa. Lancet Neurol. 2021, 20, 335. [Google Scholar] [CrossRef]
- Erves, J.C.; Mayo-Gamble, T.L.; Malin-Fair, A.; Boyer, A.; Joosten, Y.; Vaughn, Y.C.; Sherden, L.; Luther, P.; Miller, S.; Wilkins, C.H. Needs, Priorities, and Recommendations for Engaging Underrepresented Populations in Clinical Research: A Community Perspective. J. Community Health 2017, 42, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Sanderson, S.C.; Diefenbach, M.A.; Zinberg, R.; Horowitz, C.R.; Smirnoff, M.; Zweig, M.; Streicher, S.; Jabs, E.W.; Richardson, L.D. Willingness to Participate in Genomics Research and Desire for Personal Results among Underrepresented Minority Patients: A Structured Interview Study. J. Community Genet. 2013, 4, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Nuytemans, K.; Manrique, C.P.; Uhlenberg, A.; Scott, W.K.; Cuccaro, M.L.; Luca, C.C.; Singer, C.; Vance, J.M. Motivations for Participation in Parkinson Disease Genetic Research Among Hispanics versus Non-Hispanics. Front. Genet. 2019, 10, 658. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.G.; Swearingen, C.J.; Shulman, L.M.; Ye, J.; Baumgarten, M.; Tilley, B.C. Minority Enrollment in Parkinson’s Disease Clinical Trials. Parkinsonism Relat. Disord. 2009, 15, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schurz, H.; Müller, S.J.; van Helden, P.D.; Tromp, G.; Hoal, E.G.; Kinnear, C.J.; Möller, M. Evaluating the Accuracy of Imputation Methods in a Five-Way Admixed Population. Front. Genet. 2019, 10, 34. [Google Scholar] [CrossRef]
- International Parkinson Disease Genomics Consortium (IPDGC) Ten Years of the International Parkinson Disease Genomics Consortium: Progress and Next Steps. J. Parkinson’s Dis. 2020, 10, 19–30. [CrossRef] [Green Version]
- Zabetian, C.P.; Mata, I.F. Latin American Research Consortium on the Genetics of PD (LARGE-PD) LARGE-PD: Examining the Genetics of Parkinson’s Disease in Latin America. Mov. Disord. 2017, 32, 1330–1331. [Google Scholar] [CrossRef]
- Sequeiros, J.; Paneque, M.; Guimarães, B.; Rantanen, E.; Javaher, P.; Nippert, I.; Schmidtke, J.; Kääriäainen, H.; Kristoffersson, U.; Cassiman, J.-J. The Wide Variation of Definitions of Genetic Testing in International Recommendations, Guidelines and Reports. J. Community Genet. 2012, 3, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, R.H.; Lucassen, A.M. Recent Developments in Genetic/genomic Medicine. Clin. Sci. 2019, 133, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Payne, K.; Gavan, S.P.; Wright, S.J.; Thompson, A.J. Cost-Effectiveness Analyses of Genetic and Genomic Diagnostic Tests. Nat. Rev. Genet. 2018, 19, 235–246. [Google Scholar] [CrossRef]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the α-Synuclein Gene Identified in Families with Parkinson’s Disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [Green Version]
- Bandres-Ciga, S.; Diez-Fairen, M.; Kim, J.J.; Singleton, A.B. Genetics of Parkinson’s Disease: An Introspection of Its Journey towards Precision Medicine. Neurobiol. Dis. 2020, 137, 104782. [Google Scholar] [CrossRef]
- Tolosa, E.; Vila, M.; Klein, C.; Rascol, O. LRRK2 in Parkinson Disease: Challenges of Clinical Trials. Nat. Rev. Neurol. 2020, 16, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Youle, R.J. The Roles of PINK1, Parkin, and Mitochondrial Fidelity in Parkinson’s Disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [Green Version]
- Ozelius, L.J.; Senthil, G.; Saunders-Pullman, R.; Ohmann, E.; Deligtisch, A.; Tagliati, M.; Hunt, A.L.; Klein, C.; Henick, B.; Hailpern, S.M.; et al. LRRK2 G2019S as a Cause of Parkinson’s Disease in Ashkenazi Jews. N. Engl. J. Med. 2006, 354, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Zhang, Y.; Sun, Q.; Pan, H.; Tang, B. A Comprehensive Analysis of Population Differences in Variant Distribution in Parkinson’s Disease. Front. Aging Neurosci. 2019, 11, 13. [Google Scholar] [CrossRef]
- Cornejo-Olivas, M.; Torres, L.; Velit-Salazar, M.R.; Inca-Martinez, M.; Mazzetti, P.; Cosentino, C.; Micheli, F.; Perandones, C.; Dieguez, E.; Raggio, V.; et al. Variable Frequency of Variants in the Latin American Research Consortium on the Genetics of Parkinson’s Disease (LARGE-PD), a Case of Ancestry. NPJ Parkinson’s Dis. 2017, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Kestenbaum, M.; Alcalay, R.N. Clinical Features of LRRK2 Carriers with Parkinson’s Disease. Adv. Neurobiol. 2017, 14, 31–48. [Google Scholar]
- Trinh, J.; Guella, I.; Farrer, M.J. Disease Penetrance of Late-Onset Parkinsonism: A Meta-Analysis. JAMA Neurol. 2014, 71, 1535–1539. [Google Scholar] [CrossRef] [PubMed]
- Rosborough, K.; Patel, N.; Kalia, L.V. α-Synuclein and Parkinsonism: Updates and Future Perspectives. Curr. Neurol. Neurosci. Rep. 2017, 17, 31. [Google Scholar] [CrossRef]
- Lunati, A.; Lesage, S.; Brice, A. The Genetic Landscape of Parkinson’s Disease. Rev. Neurol. 2018, 174, 628–643. [Google Scholar] [CrossRef]
- Kasten, M.; Klein, C. The Many Faces of α-Synuclein Mutations. Mov. Disord. 2013, 28, 697–701. [Google Scholar] [CrossRef]
- Fuchs, J.; Nilsson, C.; Kachergus, J.; Munz, M.; Larsson, E.-M.; Schüle, B.; Langston, J.W.; Middleton, F.A.; Ross, O.A.; Hulihan, M.; et al. Phenotypic Variation in a Large Swedish Pedigree due to SNCA Duplication and Triplication. Neurology 2007, 68, 916–922. [Google Scholar] [CrossRef]
- Ryan, B.J.; Hoek, S.; Fon, E.A.; Wade-Martins, R. Mitochondrial Dysfunction and Mitophagy in Parkinson’s: From Familial to Sporadic Disease. Trends Biochem. Sci. 2015, 40, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Kasten, M.; Hartmann, C.; Hampf, J.; Schaake, S.; Westenberger, A.; Vollstedt, E.-J.; Balck, A.; Domingo, A.; Vulinovic, F.; Dulovic, M.; et al. Genotype-Phenotype Relations for the Parkinson’s Disease Genes Parkin, PINK1, DJ1: MDSGene Systematic Review. Mov. Disord. 2018, 33, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Niemann, N.; Jankovic, J. Juvenile Parkinsonism: Differential Diagnosis, Genetics, and Treatment. Parkinsonism Relat. Disord. 2019, 67, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Ligaard, J.; Sannæs, J.; Pihlstrøm, L. Deep Brain Stimulation and Genetic Variability in Parkinson’s Disease: A Review of the Literature. NPJ Parkinson’s Dis. 2019, 5, 18. [Google Scholar] [CrossRef]
- Riboldi, G.M.; Di Fonzo, A.B. Gaucher Disease, and Parkinson’s Disease: From Genetic to Clinic to New Therapeutic Approaches. Cells 2019, 8, 364. [Google Scholar] [CrossRef] [Green Version]
- Cilia, R.; Tunesi, S.; Marotta, G.; Cereda, E.; Siri, C.; Tesei, S.; Zecchinelli, A.L.; Canesi, M.; Mariani, C.B.; Meucci, N.; et al. Survival and Dementia in GBA-Associated Parkinson’s Disease: The Mutation Matters. Ann. Neurol. 2016, 80, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Do, J.; McKinney, C.; Sharma, P.; Sidransky, E. Glucocerebrosidase and Its Relevance to Parkinson Disease. Mol. Neurodegener. 2019, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Hruska, K.S.; LaMarca, M.E.; Scott, C.R.; Sidransky, E. Gaucher Disease: Mutation and Polymorphism Spectrum in the Glucocerebrosidase Gene (GBA). Hum. Mutat. 2008, 29, 567–583. [Google Scholar] [CrossRef]
- Krohn, L.; Grenn, F.P.; Makarious, M.B.; Kim, J.J.; Bandres-Ciga, S.; Roosen, D.A.; Gan-Or, Z.; Nalls, M.A.; Singleton, A.B.; Blauwendraat, C.; et al. Comprehensive Assessment of PINK1 Variants in Parkinson’s Disease. Neurobiol. Aging 2020, 91, 168.e1–168.e5. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Payne, K.; Walls, B.; Wojcieszek, J. Approach to Assessment of Parkinson Disease with Emphasis on Genetic Testing. Med. Clin. N. Am. 2019, 103, 1055–1075. [Google Scholar] [CrossRef]
- Goldman, J.S. Predictive Genetic Counseling for Neurodegenerative Diseases: Past, Present, and Future. Cold Spring Harb. Perspect. Med. 2019, 23, a036525. [Google Scholar] [CrossRef] [PubMed]
- Crook, A.; Jacobs, C.; Newton-John, T.; O’Shea, R.; McEwen, A. Genetic Counseling and Testing Practices for Late-Onset Neurodegenerative Disease: A Systematic Review. J. Neurol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, R.N.; Kehoe, C.; Shorr, E.; Battista, R.; Hall, A.; Simuni, T.; Marder, K.; Wills, A.-M.; Naito, A.; Beck, J.C.; et al. Genetic Testing for Parkinson Disease: Current Practice, Knowledge, and Attitudes among US and Canadian Movement Disorders Specialists. Genet. Med. 2020, 22, 574–580. [Google Scholar] [CrossRef] [Green Version]
- Rajan, R.; Divya, K.P.; Kandadai, R.M.; Yadav, R.; Satagopam, V.P.; Madhusoodanan, U.K.; Agarwal, P.; Kumar, N.; Ferreira, T.; Kumar, H.; et al. Genetic Architecture of Parkinson’s Disease in the Indian Population: Harnessing Genetic Diversity to Address Critical Gaps in Parkinson’s Disease Research. Front. Neurol. 2020, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Loesch, D.P.; Horimoto, A.R.V.R.; Heilbron, K.; Sarihan, E.I.; Inca-Martinez, M.; Mason, E.; Cornejo-Olivas, M.; Torres, L.; Mazzetti, P.; Cosentino, C.; et al. Characterizing the Genetic Architecture of Parkinson’s Disease in Latinos. Ann. Neurol. 2021, 90, 353–365. [Google Scholar] [CrossRef]
- Tipton, P.W.; Jaramillo-Koupermann, G.; Soto-Beasley, A.I.; Walton, R.L.; Soler-Rangel, S.; Romero-Osorio, Ó.; Díaz, C.; Moreno-López, C.L.; Ross, O.A.; Wszolek, Z.K.; et al. Genetic Characterization of Parkinson’s Disease Patients in Ecuador and Colombia. Parkinsonism Relat. Disord. 2020, 75, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Williams, U.; Bandmann, O.; Walker, R. Parkinson’s Disease in Sub-Saharan Africa: A Review of Epidemiology, Genetics and Access to Care. J. Mov. Disord. 2018, 11, 53–64. [Google Scholar] [CrossRef]
- Morris, A.P. Transethnic Meta-Analysis of Genomewide Association Studies. Genet. Epidemiol. 2011, 35, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Asimit, J.L.; Hatzikotoulas, K.; McCarthy, M.; Morris, A.P.; Zeggini, E. Trans-Ethnic Study Design Approaches for Fine-Mapping. Eur. J. Hum. Genet. 2016, 24, 1330–1336. [Google Scholar] [CrossRef] [Green Version]
- Bandres-Ciga, S.; Saez-Atienzar, S.; Kim, J.J.; Makarious, M.B.; Faghri, F.; Diez-Fairen, M.; Iwaki, H.; Leonard, H.; Botia, J.; Ryten, M.; et al. Large-Scale Pathway Specific Polygenic Risk and Transcriptomic Community Network Analysis Identifies Novel Functional Pathways in Parkinson Disease. Acta Neuropathol. 2020, 140, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Nalls, M.A.; Bras, J.; Hernandez, D.G.; Keller, M.F.; Majounie, E.; Renton, A.E.; Saad, M.; Jansen, I.; Guerreiro, R.; Lubbe, S.; et al. NeuroX, a Fast and Efficient Genotyping Platform for Investigation of Neurodegenerative Diseases. Neurobiol. Aging 2015, 36, 1605.e7–1605.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blauwendraat, C.; Faghri, F.; Pihlstrom, L.; Geiger, J.T.; Elbaz, A.; Lesage, S.; Corvol, J.-C.; May, P.; Nicolas, A.; Abramzon, Y.; et al. NeuroChip, an Updated Version of the NeuroX Genotyping Platform to Rapidly Screen for Variants Associated with Neurological Diseases. Neurobiol. Aging 2017, 57, 247.e9–247.e13. [Google Scholar] [CrossRef] [Green Version]
- Benner, C.; Spencer, C.C.A.; Havulinna, A.S.; Salomaa, V.; Ripatti, S.; Pirinen, M. FINEMAP: Efficient Variable Selection Using Summary Data from Genome-Wide Association Studies. Bioinformatics 2016, 32, 1493–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kichaev, G.; Yang, W.-Y.; Lindstrom, S.; Hormozdiari, F.; Eskin, E.; Price, A.L.; Kraft, P.; Pasaniuc, B. Integrating Functional Data to Prioritize Causal Variants in Statistical Fine-Mapping Studies. PLoS Genet. 2014, 10, e1004722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustos, B.I.; Billingsley, K.; Blauwendraat, C.; Raphael Gibbs, J.; Gan-Or, Z.; Krainc, D.; Singleton, A.B.; Lubbe, S.J. For the International Parkinson’s Disease Genomics Consortium (IPDGC). Genome-Wide Contribution of Common Short-Tandem Repeats to Parkinson’s Disease Genetic Risk. medRxiv 2021. [Google Scholar] [CrossRef]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional Mapping and Annotation of Genetic Associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- Faghri, F.; Hashemi, S.H.; Leonard, H.; Scholz, S.W.; Campbell, R.H.; Nalls, M.A.; Singleton, A.B. Predicting Onset, Progression, and Clinical Subtypes of Parkinson Disease Using Machine Learning. bioRxiv 2018, 338913. [Google Scholar] [CrossRef]
- Makarious, M.B.; Leonard, H.L.; Vitale, D.; Iwaki, H.; Sargent, L.; Dadu, A.; Violich, I.; Hutchins, E.; Saffo, D.; Bandres-Ciga, S.; et al. Multi-Modality Machine Learning Predicting Parkinson’s Disease. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bressan, E.; Reed, X.; Bansal, V.; Hutchins, E.; Cobb, M.M.; Webb, M.G.; Alsop, E.; Grenn, F.P.; Illarionova, A.; Savytska, N.; et al. The Foundational Data Initiative for Parkinson’s Disease (FOUNDIN-PD): Enabling Efficient Translation from Genetic Maps to Mechanism. bioRxiv 2021. [Google Scholar] [CrossRef]


{kind=link}
| Gene | Year of Discovery | Reported Variants | Frequency | Inheritance | Confidence as a PD Gene |
|---|---|---|---|---|---|
| SNCA * | 1997, 2003 | Missense or multiplication | Very rare | Dominant | Very high |
| PRKN * | 1998 | Missense or loss of function | Rare | Recessive | Very high |
| UCHL1 | 1998 | Missense | Unclear | Dominant | Low |
| PARK7 * | 2003 | Missense | Very rare | Recessive | Very high |
| LRRK2 * | 2004 | Missense | Common | Dominant | Very high |
| PINK1 * | 2004 | Missense or loss of function | Rare | Recessive | Very high |
| POLG | 2004 | Missense or loss of function | Rare | Dominant | High |
| HTRA2 | 2005 | Missense | Unclear | Dominant | Low |
| ATP13A2 * | 2006 | Missense or loss of function | Very rare | Recessive | Very high |
| FBXO7 * | 2008 | Missense | Very rare | Recessive | Very high |
| GIGYF2 | 2008 | Missense | Unclear | Dominant | Low |
| GBA * | 2009 | Missense or loss of function | Common | Dominant (incomplete penetrance) | Very high |
| PLA2G6 * | 2009 | Missense or loss of function | Rare | Recessive | Very high |
| EIF4G1 | 2011 | Missense | Unclear | Dominant | Low |
| VPS35 * | 2011 | Missense | Very rare | Dominant | Very high |
| DNAJC6 | 2012 | Missense or loss of function | Very rare | Recessive | High |
| SYNJ1 | 2013 | Missense or loss of function | Very rare | Recessive | High |
| DNAJC13 | 2014 | Missense | Unclear | Dominant | Low |
| TMEM230 | 2016 | Missense | Unclear | Dominant | Low |
| VPS13C | 2016 | Missense or loss of function | Rare | Recessive | High |
| LRP10 | 2018 | Missense or loss of function | Unclear | Dominant | Low |
| NUS1 | 2018 | Missense | Unclear | Recessive | Low |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsayed, I.; Martinez-Carrasco, A.; Cornejo-Olivas, M.; Bandres-Ciga, S. Mapping the Diverse and Inclusive Future of Parkinson’s Disease Genetics and Its Widespread Impact. Genes 2021, 12, 1681. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111681
Elsayed I, Martinez-Carrasco A, Cornejo-Olivas M, Bandres-Ciga S. Mapping the Diverse and Inclusive Future of Parkinson’s Disease Genetics and Its Widespread Impact. Genes. 2021; 12(11):1681. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111681
Chicago/Turabian StyleElsayed, Inas, Alejandro Martinez-Carrasco, Mario Cornejo-Olivas, and Sara Bandres-Ciga. 2021. "Mapping the Diverse and Inclusive Future of Parkinson’s Disease Genetics and Its Widespread Impact" Genes 12, no. 11: 1681. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111681