
Functional Conservation and Divergence of Five AP1/FUL-like Genes in Marigold (Tagetes erecta L.)

1
Key Laboratory of Horticultural Plant Biology, Ministry of Education, College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Shizishan Street No. 1, Wuhan 430070, China
2
Institute of Vegetable, Wuhan Academy of Agricultural Sciences, Wuhan 430207, China
*
Author to whom correspondence should be addressed.
Genes 2021, 12(12), 2011; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122011
Submission received: 3 November 2021
/
Revised: 10 December 2021
/
Accepted: 13 December 2021
/
Published: 17 December 2021
(This article belongs to the Special Issue Plant Specialized Metabolism: From Genetics to Phenotype)
Abstract
:Members of AP1/FUL subfamily genes play an essential role in the regulation of floral meristem transition, floral organ identity, and fruit ripping. At present, there have been insufficient studies to explain the function of the AP1/FUL-like subfamily genes in Asteraceae. Here, we cloned two euAP1 clade genes TeAP1-1 and TeAP1-2, and three euFUL clade genes TeFUL1, TeFUL2, and TeFUL3 from marigold (Tagetes erecta L.). Expression profile analysis demonstrated that TeAP1-1 and TeAP1-2 were mainly expressed in receptacles, sepals, petals, and ovules. TeFUL1 and TeFUL3 were expressed in flower buds, stems, and leaves, as well as reproductive tissues, while TeFUL2 was mainly expressed in flower buds and vegetative tissues. Overexpression of TeAP1-2 or TeFUL2 in Arabidopsis resulted in early flowering, implying that these two genes might regulate the floral transition. Yeast two-hybrid analysis indicated that TeAP1/FUL proteins only interacted with TeSEP proteins to form heterodimers and that TeFUL2 could also form a homodimer. In general, TeAP1-1 and TeAP1-2 might play a conserved role in regulating sepal and petal identity, similar to the functions of MADS-box class A genes, while TeFUL genes might display divergent functions. This study provides a theoretical basis for the study of AP1/FUL-like genes in Asteraceae species.
1. Introduction
Although flowers show great diversity in morphology, structure, composition, color, and function, they are usually composed of four distinct concentric whorl floral organs: sepals in the outermost whorl, petals in the second whorl, stamens (male reproductive organs) in the third whorl, and carpels (female reproductive organs) in the innermost whorl [1]. The exploration of the mechanism of distinct floral organ formation has undergone a long-term challenge in plant developmental genetics [1,2,3,4]. Based on the study of the floral formation in model plants, such as Arabidopsis thaliana L., Antirrhinum majus L., and Petunia hybrida Vilmorin, the fate of the different floral organs was considered to be determined by a complex regulatory network composed of MADS-box proteins [5].
The MADS-box protein family is one of the most widely studied transcriptional factor families in angiosperm, and this family plays a key role in regulating floral meristem development, floral organ identity, fruit and seed development, vegetative tissue development, and flowering time [6,7,8]. MADS box genes in higher plants are reported to have undergone several duplication events that promote the evolution of morphological complexity of the flower [9], thus allowing MADS-box genes to cluster into several major subfamilies [10,11,12]. One subfamily of MADS-box genes forms the angiosperm-specific APETALA1/FRUITFUL (AP1/FUL) lineage via gene duplication. Phylogenetic analyses reveal that AP1/FUL lineage has undergone numerous duplication events throughout angiosperm diversification [13,14]. AP1 genes diverge into two types of AP1 lineage genes within the core eudicot, namely, euAP1 (Arabidopsis AP1 and Antirrhinum SQUA) and euFUL, which is likely to be part of the whole genome duplication event before the diversification of the core eudicots, and this event is often known as the γ event [15,16]. Contrary to core eudicots, non-core eudicots have only FUL-like clade genes [13,14,17,18]. Within the model plant Arabidopsis, the euAP1 clade is subdivided into AP1 and CAULIFLOWE (CAL) genes [19,20], whose amino acid sequences possess an acidic domain and a farnesylation motif (CaaX) in their 3′ end of the coding sequence [13,21]. AP1 and CAL are accumulated in floral meristems, sepals, and petals primordia [22,23]. In Arabidopsis, ap1 mutation results in the absence of petals, the transformation of sepals into bract-like structures, and the production of secondary flowers from the axils of the first whorl organs [22,24]. Overexpression of AP1 leads to remarkable early flowering and transformation of inflorescence shoot apical meristem into floral meristem [25]. In single Arabidopsis cal mutants, there are no remarkable changes in floral organs, but the cal mutation enhances the repetitive branching pattern in the floral meristem of ap1 mutants [24,26]. In other core eudicots, the ap1 mutation only changes the sepal structure, but it does not affect petal structure [27,28]. Furthermore, ectopic expression of Fortunella crassifolia Swingle FcAP1 in Arabidopsis [29] and heterologous overexpression of the AP1-like gene Betula pendula Roth. BpMADS3 [30] and Pisum sativum L. PEAM4 [31] in Nicotiana tabacum L. cause early flowering with floral meristem development unaffected. In addition, AP1, together with TFL1 and LFY, is the key inflorescence regulator in Arabidopsis [22,32,33]. In general, euAP1 clade genes exhibit a conserved function in specifying the floral meristem and sepal identity in core eudicots.
The euFUL genes show a conserved function in promoting the transition from vegetative meristems to reproductive meristems and in regulating fruit development in core eudicots. The euFUL proteins are characterized by possessing a conserved six-hydrophobic-amino-acid motif (FUL-like motif) in the C domain [13,14]. This motif is considered to be conserved in the entire ancestral gene lineage, and its occurrence is prior to the euFUL/euAP1 duplication, but its function remains unclear [13,14]. In Arabidopsis, euFUL is divided into FUL (or euFULI) and AGAMOUS-like 79 (AGL79 or euFULII) in the duplication event of euFUL clade [34,35,36]. Different from euAP1, FUL is mainly expressed in growing leaves, inflorescence meristems, carpel primordia, and young siliques [25,37]. Arabidopsis FUL is redundant with AP1 and CAL in regulating the floral meristem identity, and it also regulates the flowering time, axillary meristem activation, meristem determinacy, and plant longevity [23,38]. In ful mutants, the cauline leaf development was terminated, and the floral development was disrupted [37,39]. The transcript of AGL79 is detected in roots, but its function remains unclear [34,35]. Recently, only the limited functional analysis of euFUL genes in other core eudicots is available. Similar to the FUL gene function in Arabidopsis, ectopic expression of DEFICIENS-homolog28 (DEFH28, a euFULII gene, Antirrhinum) [40] in Arabidopsis causes early flowering, transformation of inflorescence into a terminal flower, and silique indehiscence. Moreover, overexpression of NtFUL in N. tabacum also results in early flowering and failure in capsule dehiscence [41]. In P. hybrida, silencing PETUNIA FLOWERING GENE (PFG, an euFULⅠ gene) leads to the interruption of inflorescence formation, thus maintaining vegetative growth [42].
Asteraceae is one of the most abundant and widespread families of flowering plants, and it has a specific capitulum consisting of two flower types: the outer are the sterile ray florets and the inner are the fertile disk florets. The specific inflorescence makes Asteraceae a suitable material for studying the evolution and function of MADS-box genes related to floral organ development. Nevertheless, the available functional information on AP1/FUL genes in Asteraceae is restricted to Gerbera hybrida Hort. [43], Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Ling et Shih [44], and Chrysanthemum morifolium Ramat. [45,46]. The expression patterns of AP1/FUL-like genes vary with various Asteraceae species. The euAP1-like genes CDM111 (C. morifolium) and GASQUA3 (G. hybrida) are highly expressed in sepals and petals, while their homologous gene GASQUA1 (G. hybrida) is not expressed in floral meristem or in perianth primordia [43,45,47]. The FUL-like genes GSQUA2 and GSQUA5 are only expressed in florescence and floral organs, while GSUA4 and CDM41 are also expressed in leaves [43,45]. The function of AP1/FUL-like genes in Asteraceae remains unclear. Overexpression of the AP1-like gene CDM111 or FUL-like gene ClM8 in Arabidopsis results in the altered flowering time and inflorescence structure [44,45]. However, overexpression of GSQUA2 (a FUL-like gene, homolog to DEFH28) in gerbera results in a dwarf plant, early flowering, and vegetative abnormality, but it does not affect inflorescence structure [43].
Marigold (T. erecta ) is a popular ornamental plant and economic crop, whose flowers are rich in lutein. As a member of Asteraceae, marigold also has a typical capitulum. Compared with the complex inflorescence structure of G. hybrida, marigold wears a simple inflorescence, which consists of two distinct flower types, namely, the outermost ray flowers and the disk flowers. Furthermore, the ray flowers in marigold retain female pistils, whereas the ray flowers in Helianthus annuus L. are sterile because of only filamentous remnants in aborted stamens and empty ovaries. In addition, the whole life cycle of marigold lasts only 2-3 months from sowing to flowering. Furthermore, in the evolutionary history of the Asteraceae family, marigold undergoes a long evolution process and it belongs to the Tageteae clade [48]. These characteristics make marigold a valuable material for studying the molecular mechanism of marigold inflorescence formation. In our previous work, we have obtained functionally characterized class B (TePI, TeAP3-1, TeAP3-2, TeTM6-1, and TeTM6-2) [49], C (TeAG1 and TeAG2) [50], D (TeAGL11-1 and TeAGL11-2) [50], and E (TeSEP1, TeSEP3-1, TeSEP3-2, TeSEP3-3, and TeSEP4) [51] genes, which are active during marigold inflorescence and floret development and have specific expression patterns in floral organs. In this study, we cloned and characterized five AP1/FUL-like genes in marigold, whose distinct expression patterns, protein interaction patterns, and different phenotypes in Arabidopsis transgenic lines might imply divergent functions of these five genes in regulating the floral meristem development, floral organ identity, and flowering time.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
Marigold (T. erecta, M525B-1) is an inbred line with more than 10 generations of continuous self-crossing, and it has only one whorl of ray florets outside the capitulum [52]. Marigold plants were grown in the experimental field of Huazhong Agricultural University (lat. 30°28’36.5” N, long, 114°21’59.4” E) under natural conditions. To investigate AP1/FUL-like genes expression patterns, the samples of vegetative tissues, flower buds from different stages, and floral organs in the blooming period were collected as described by Ai et al. [49], and were frozen immediately in liquid nitrogen and stored at −80 °C.
Arabidopsis ecotype Columbia (Col-0) plants were used for functional analysis of AP1/FUL-like genes of marigold. Plants were grown in a chamber at 22 °C under long-day conditions (16 h light, 8 h dark) with 70% relative humidity.
2.2. Total RNA Extraction, Isolation, and Bioinformatics Analysis of AP1/FUL-like Genes from Marigold
The total RNA of each sample was isolated with a PLANTpure kit (Aidlab, Beijing, China) according to the manufacturer’s protocol. The quantity and the quality of RNA samples were analyzed by a Nano-Drop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) and by running test gels with ethidium bromide staining. The first-strand cDNA was synthesized by the TRUEscript RT reagent Kit with gDNA Eraser (Aidlab, Beijing, China) with the Oligo-dT primers. Five AP1/FUL-like genes were selected from the transcriptomic data (accession number SRP066084) [49] and full-length transcriptomic data (unpublished), and named TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3, respectively. To verify the accuracy of these five gene sequences, the specific primers TeAP1-1-full-F/R, TeAP1-2-full-F/R, TeFUL1-full-F/R, TeFUL2-full-F/R, and TeFUL3-full-F/R were designed in the 3’ and 5’ terminal region by Primer Premier 5 software (Premier Biosoft International, Palo Alto, CA, USA) (Supplementary Table S1) and used to clone full-length gene sequences. The PCR was programmed using the following parameters: 94 °C for 4 min; 38 cycles of 94 °C for 30 s, 56 °C for 30 s, and 72 °C for 2 min, and a final extension at 72 °C for 10 min. The PCR amplification fragments were purified and then cloned into a pMD18-T vector (Takara, Dalian, China). Positive clones were verified by PCR using M13-F/R universal primers, and 3–5 positive clones were selected and sequenced by the Sangon company in Shanghai.
The Open Reading Frames (ORFs) of these five AP1/FUL-like genes were predicted online (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/orffinder/ access on 2 November 2021) and were blasted against the NCBI to search for their homologous sequences. To identify the conserved motifs of AP1/FUL amino acids, the multiple sequence alignment was performed by using the DNAMAN (v.6.0) software (https://www.lynnon.com access on 2 November 2021) and BoxShade (https://embnet.vital-it.ch/software/BOX_form.html access on 2 November 2021). A total of 36 AP1/FUL-like genes and 4 AGL6-like genes derived from model plants and G. hybrida were downloaded from the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov access on 2 November 2021) for phylogenetic analysis. The gene accession number was listed in Supplementary Table S2. Four AGL6-like proteins from model plants (Arabidopsis and petunia) and Asteraceae species (gerbera) were used as the outgroup. The construction of the phylogenetic tree was based on amino acid alignment with the default settings of MUSCLE in MEGA (v. 7.0). A phylogenetic tree was constructed by the neighbor-joining (NJ) method with bootstrap confidence values of 1000 replicates, and distances were calculated with Poisson corrections for multiple substitutions.
2.3. Gene Expression Analysis
To analyze the expression of five A class genes in marigold, the total RNA from the samples of roots, tender stems, fresh leaves, different sizes of flower buds (0–1, 2–3, 4–5 and 6–7 mm in diameter, respectively), sepals, petals and pistils of ray and disk florets, stamens of disk florets, receptacles, bracts, and ovaries of opened flowers were isolated with PLANTpure kit (Aidlab, Beijing, China) according to the manufacturer’s protocol. The total RNA was reverse-transcribed by the TRUEscript RT reagent Kit with gDNA Eraser (Aidlab, Beijing, China) with the Oli-go-dT primers. The reverse transcription reaction contained total RNA 1 μg, 4 × gDNA Eraser mix 4 μL, 5 × TRUE RT MasterMixⅡ (Aidlab, Beijing, China), and double-distilled water to supply a final volume of 20 μL. The reverse transcription reactions were incubated at 42 °C for 20 min and 85 °C for 5 s. The analysis of expression patterns of AP1/FUL-like genes in different tissues and different development stages of flower buds was performed by quantitative real-time PCR (qRT-PCR). The specific primers were designed within the non-conservative C-terminal region using the Primer Premier 5.0 software to amplify products between 90 and 200 bp (Supplementary Table S1), and the products were named qTeAP1-1, qTeAP1-2, qTeFUL1, qTeFUL2, and qTeFUL3. The specific and unique PCR products for each primer pair were confirmed by 1.2% agarose gel electrophoresis. The qRT-PCR was carried out in an optical 384-well plate in the QuantStudio 6 Flex real-time PCR system (Applied Biosystems, Palo Alto, CA, USA) with SYBR Primix Ex Taq kit (TaKaRa, Dalian, China) according to manufacturer’s instructions. The qRT-PCR data were analyzed in the ABI 7500 Detection System (Applied Biosystems, Palo Alto, CA, USA). The qRT-PCR products were amplified in 10 μL reaction solution containing 1 μL template of the reaction mixture, 5 μL 2 × SYBR Green Master Mix (TaKaRa, Dalian, China), 0.2 μL forward primer and reverse primer (10 μmol/μL for primers), and double-distilled water to supply a final volume of 10 μL. The PCR was performed as follows: 95 °C for 2 min and 40 cycles of 95 °C for 10 s and 60 °C for 20 s. The expression level of each gene was summarized from three replicates for each sample. The house-keeping gene β-actin was used as an internal control for qRT-PCR and the relative expression levels were calculated using the 2–ΔΔCt method [53].
2.4. Yeast Two-Hybrid Assay
The full-length coding sequences of TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 were amplified using primers with specific restriction sites and cloned into the activation domain plasmid pGBKT7 (Clontech, Palo Alto, CA, USA) and into binding domain plasmid pGADT7 (Clontech, Palo Alto, CA, USA), respectively. All constructs were confirmed by sequencing analyses. The primers were presented in Supplementary Table S1. The bait and prey constructs of five class B genes (TeAP3-1, TeAP3-2, TePI, TeTM6-1, and TeTM6-2) and class C + D genes (C: TeAG1 and TeAG2, D: TeAGL11-1 and TeAGL11-2) were previously described by Ai et al. [49] and Zhang et al. [50], respectively. The full-length sequences of six class E genes (TeSEP1, TeSEP3-1, TeSEP3-2, TeSEP3-3, TeSEP4 and TeAGL6) were downloaded from NCBI, and bait and prey recombinants of these six class E genes were also constructed, respectively. Both bait and prey constructs were transformed into yeast cell strain AH109 using LiAc method (Clontech) following the Frozen-EZ Yeast Transformation II Kit protocols (Zymo Research Corp, Irvine, CA, USA). Interaction results between bait proteins and empty AD, between prey proteins and empty BD, between empty BD and empty AD were used as negative controls. The interaction results between pGBKT7-53 and pGADT7-T7 vectors were used as a positive control. Yeast double transformants were plated onto SD medium without tryptophane (Trp) and leucine (Leu) (Sigma, St. Louis, MO, USA, A8056), and medium was incubated at 30 °C for 3-5 days. Positive clones were verified by PCR with general primers AD-R/F or BD-F/R (Supplementary Table S1). Three randomly selected positive yeast cells were spotted onto the X-α-gal-supplemented selection medium without Leu, Trp, histidine (His), and adenine (Ade). The interaction between the tested proteins was analyzed after 3–5-day incubation of the positive yeast cells at 30 °C.
2.5. Vector Construction and Plant Transformation
The full-length coding sequences of TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 were amplified by using primer pairs with specific restriction sites (Supplementary Table S1), and the amplification products were ligated to the pCAMBIA2300s plasmid, which harbored the CaMV35S promoter and kanamycin resistance (Kan) gene (Supplementary Figure S1). The recombinant plasmids were named 35S:TeAP1-1, 35S:TeAP1-2, 35S:TeFUL1, 35S:TeFUL2, and 35S:TeAFUL3, respectively. All the recombinant plasmids were introduced into Escherichia coli DH5a and tested by sequencing. These plasmids were separately transformed into chemically competent Agrobacterium tumefaciens strain GV3101, which was further transformed into wild-type Arabidopsis ecotype Columbia plants by the floral dip method [54]. T1 and T2 generation transgenic plants were selected in solid medium containing 50 μg/mL kanamycin and verified by PCR with a general forward primer of 35S-F and gene-specific reverse primers 35S-TeAP1-1-R, 35S-TeAP1-2-R, 35S-TeFUL1-R, 35S-TeFUL2-R, and 35S-TeFUL3-R (Supplementary Table S1), respectively. The genomic DNA was isolated from the transgenic plants and wild-type Arabidopsis, respectively. The transcript levels of TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 were analyzed by qRT-PCR and semi-quantitative PCR (Semi-PCR). The total RNA of blooming flowers from T1 transgenic plants and wild-type plants was isolated and reverse-transcribed with the above-mentioned reagent kit. The Arabidopsis house-keeping gene EF1α (AtEF1α, AT5G60390) was used as a control for qRT-PCR and semi-PCR. QRT-PCR were performed in the same way as described above, and the relative expression levels were calculated using the 2−ΔΔCt method. The semi-PCR was performed as follows: 94 °C for 4 min, 24–26 cycles of 94 °C for 10 s, 60 °C for 30 s and 72 °C for 30 s, final extension for 5 min at 72 °C. The 24-26 cycles of semi-PCR were designed for the house-keeping gene EF1α, and 30–32 cycles of semi-PCR were designed for exogenous genes. Phenotype changes of T1 and T2 generation transgenic plants were analyzed. To testify the segregation tests, 16 kanamycin-resistant transgenic plants of the T2 generation lines that fitted a segregation ratio of 3:1 were chosen to record main morphological traits. The transcript levels of some endogenous genes of T3 generations were analyzed.
2.6. Expression Analysis of Endogenous Genes in Transgenic Plants
In order to investigate the conserved functions of AP1/FUL-like genes in marigold and to reveal the mechanism underlying phenotypic changes of transgenic lines 35S:TeAP1-2 and 35S:TeFUL2, the transcript levels of some AP1-regulated endogenous genes (including LFY, FT, SEP3, SOC1, SVP, TFL1, AGL24, and SPL9) were analyzed by qRT-PCR [55]. Total RNA was isolated from 10-day-old T3 transgenic lines 35S:TeAP1-2 and 35S:TeFUL2 and wild-type Arabidopsis 10-day-old seedlings. Reverse transcription and qRT-PCR were performed in the same way as described above. The gene-specific primers are listed in Supplementary Table S1.
3. Results
3.1. Isolation and Phylogenetic Analysis of TeAP1/FUL-like Genes
The full-length sequences of five AP1/FUL-like genes were amplified by using gene-specific primers. In this study, the cDNA of ray floret sepals was used as a template to amplify the full-length sequences of TeAP1-1 and TeAP1-2. The cDNA from different sizes of flower buds (0–1 mm in diameter and 3–4 mm in diameter) was used as a template to clone full-length sequences of TeFUL1, TeFUL2, and TeFUL3. In order to further identify the putative homologs of AP1 and FUL genes, we blasted nucleotide sequences of these five genes against NCBI. The blast search results indicated that two different AP1-like genes and three different FUL-like genes were detected with the two AP1-like genes designated as TeAP1-1 (Acc. No. MT394170), TeAP1-2 (Acc. No. MT394171), and three FUL-like genes designated as TeFUL1 (Acc. No. MT394172), TeFUL2 (Acc. No. MT394173), and TeFUL3 (Acc. No. MT394174), respectively. Sequence analysis revealed that the five putative proteins encoded by these five genes were composed of 246, 247, 235, 235, and 242 amino acids, respectively. The putative TeAP1-1 and TeAP2-1 proteins shared more than 89% amino-acid identity, and the identity between these two marigold putative proteins and one Arabidopsis AP1 clade protein was lower than 60% at the amino acid level (Supplementary Table S3). Three marigold putative FUL homologous proteins shared relatively low identity, and the identity between these three marigold putative FUL homologous proteins and two Arabidopsis FUL clade proteins was lower than 55% at amino acid level (Supplementary Table S3). Multiple sequence alignment and conservation analysis of AP1/FUL proteins indicated that all TeAP1/FUL proteins contained one conserved MADS domain, one less conserved I domain, one K domain, and one variable C-terminal domain (Figure 1). The putative proteins of TeAP1-1 and TeAP1-2 possessed one typical euAP1-motif (CFPS) containing both an acidic domain and a farnesylation motif (CaaX, shown at their C termini) (Figure 1). In addition, the characteristic FUL motif was shared by the three TeFUL proteins (Figure 1).
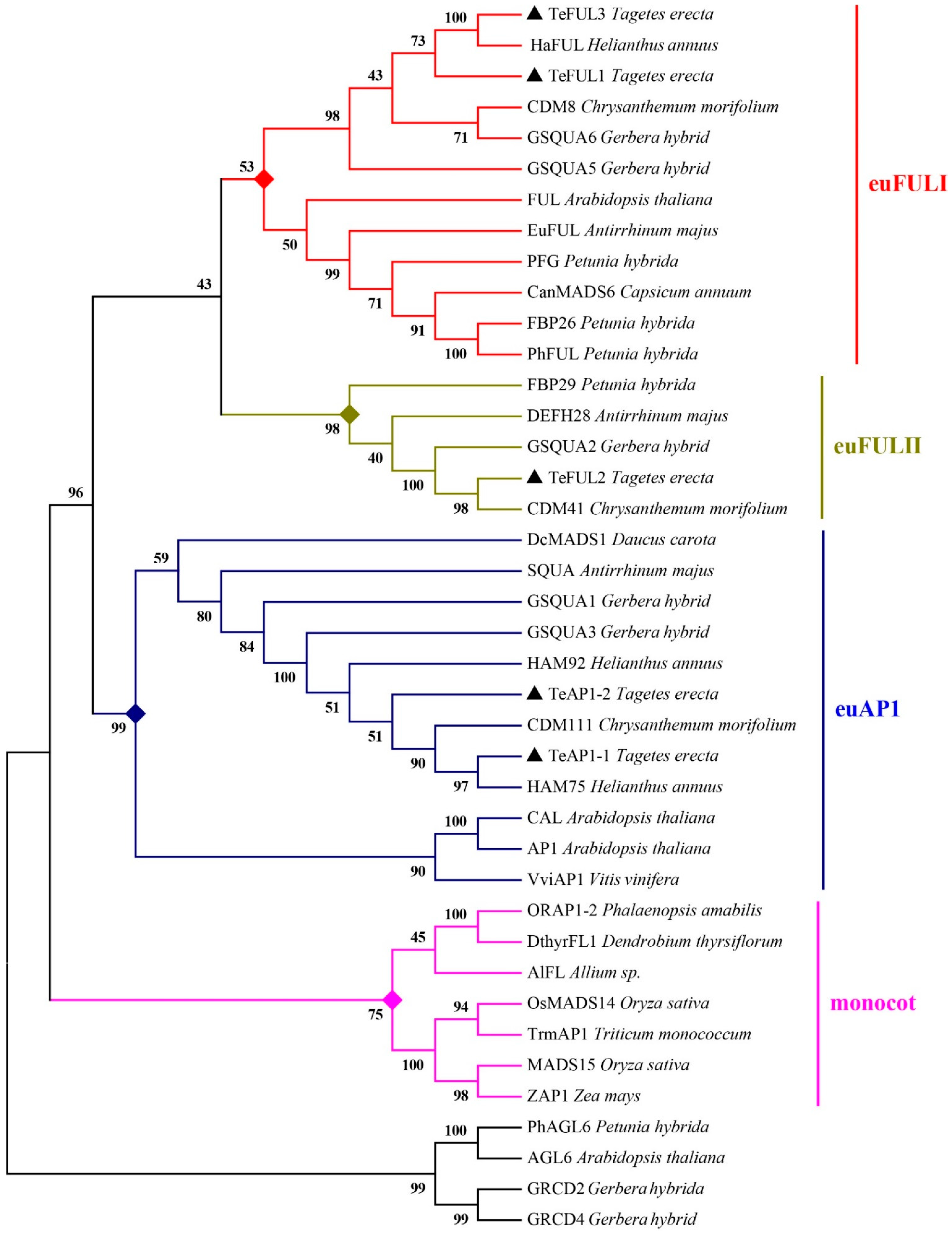
To investigate the relationship between TeAP1/FUL genes and other members of AP1 and FUL clades, a phylogenetic analysis was carried out by using amino acid sequences of the AP1/FUL clade from other plant species and those of AGL6 subfamily proteins acting as an outgroup (Figure 2). TeAP1-1 and TeAP1-2 were orthologous to Arabidopsis AP1 and Antirrhinum SQUA. TeFUL1 and TeFUL3 were phylogenetically close to euFULI, and TeFUL2 was orthologous to the Antirrhinum protein DEFH28 belonging to euFULII protein. Notably, TeFUL1 and HaFUL (H. annuus) shared 78.39% amino-acid identity, and TeAP1-1 and HAM75 (H. annuus) were more closely related to each other with over 97.45% amino-acid identity. The high homology might indicate their functional similarity.
3.2. Expression Analysis of TeAP1/FUL-like Genes in Marigold
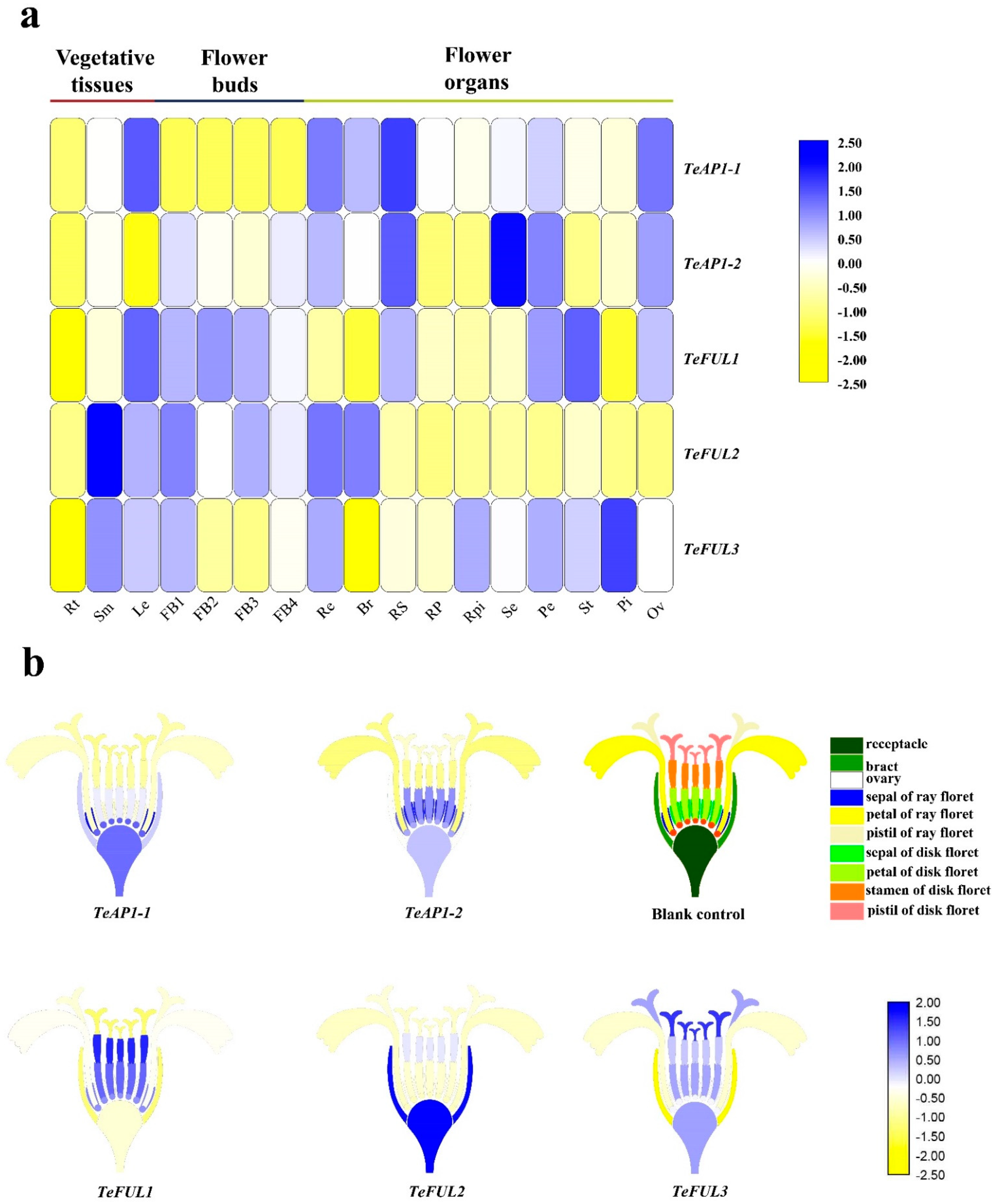
The expression patterns for these five AP1/FUL genes in different vegetative tissues, floral organs, and different development stages of flower buds were examined by qRT-PCR. TeAP1-1 was mainly expressed in leaves, receptacles, bracts, sepals of ray florets, petals of disk florets, and ovaries but not expressed in flower buds and roots (Figure 3 and Supplementary Figure S2). Compared to TeAP1-1, TeAP1-2 was weakly expressed in different development stages of flower buds but was highly expressed in receptacles, sepals of ray and disk florets, petals of disk florets, and ovaries (Figure 3a and Supplementary Figure S2). TeFUL1 and TeFUL3 shared a similar expression pattern, and they were widely expressed in vegetative and reproductive tissues (Figure 3a and Supplementary Figure S2). Some differences in expression levels in some tissues were also detected between TeFUL1 and TeFUL3. For example, TeFUL1 was expressed mainly in petals of disk florets, stamens, ovaries and sepals of ray florets, while TeFUL3 was highly expressed in all floral organs of two-type florets and receptacles (Figure 3b and Supplementary Figure S2). Contrary to TeFUL1 and TeFUL3, TeFUL2 was highly expressed in vegetative tissues, flower buds, receptacles, and bracts, and it was weakly expressed in floral organs (Figure 3a and Supplementary Figure S2).
3.3. Interactions between TeAP1/FUL Proteins and Other MADS-Box Proteins in Marigold
A yeast two-hybrid analysis was performed to evaluate the interaction strength between AP1/FUL proteins and class B, class C, class D, or class E proteins. The marigold proteins were individually fused to the binding domain and the activation domain and then were pairwise recombined in both directions. No autoactivation was observed among these five proteins (Supplementary Figure S3a). As shown in Table 1 and Supplementary Figure S3b–e, TeAP1/FUL proteins only interacted with SEPATELLA (SEP) proteins, but they did not interact with class B (TePI, TeAP3-1, TeAP3-2, TeTM6-1 and TeTM6-2), class C (TeAG1 and TeAG2), and class D (TeAGL11-1 and TeAGL11-2) proteins. TeAP1-1 and TeAP1-2 exhibited a similar protein interaction pattern, both of which interacted with class E proteins TeSEP3-2 and TeSEP3-3 to form heterodimers (Table 1, Supplementary Figure S3b–e). Contrary to two TeAP1 proteins, three TeFUL proteins displayed different protein interaction patterns. TeFUL1 only interacted with TeAGL6. TeFUL2 interacted with TeSEP3-2, TeAGL6, and itself. TeFUL3 interacted with TeSEP1, TeSEP3-1, TeSEP3-2, TeSEP3-3, and TeSEP4 to form heterodimers (Table 1, Supplementary Figure S3b,e).
3.4. Early Flowering Caused by Ectopic Expression of TeAP1-2 and TeFUL2 in Arabidopsis
To explore the potential functions of TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 genes, functional analyses were performed by overexpressing these five genes in Arabidopsis with the cauliflower mosaic virus 35S promoter. After kanamycin selection and PCR verification, a total of 63, 31, 42, 26, and 45 independent T1 transgenic plants (namely, 35S:TeAP1-1, 35S:TeAP1-2, 35S:TeFUL1, 35S:TeFUL2, and 35S:TFUL3) were obtained, respectively. The transcript level analysis revealed that five AP1/FUL-like genes (TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3) were successfully expressed in Arabidopsis plants (Figure 4l,m, and Supplementary Figure S4a–f). Compared with wild-type plants, the transgenic plants containing 35S:TeAP1-1, 35S:TeFUL1, and 35S:TeFUL3 exhibited no visual phenotypical changes. However, fourteen 35S:TeFUL1 and eleven 35S:TeFUL3 transgenic plants displayed early flowering. According to the phenotypic alterations, 2–6 T2 transgenic lines were, respectively, selected from the Arabidopsis transgenic plant whose progenies showed a 3:1 segregation ratio for kanamycin resistance, which may indicate a single-copy insertion of transgenes. Sixteen T2 transgenic plants for each line were used to investigate the flowering time and floral phenotypes.
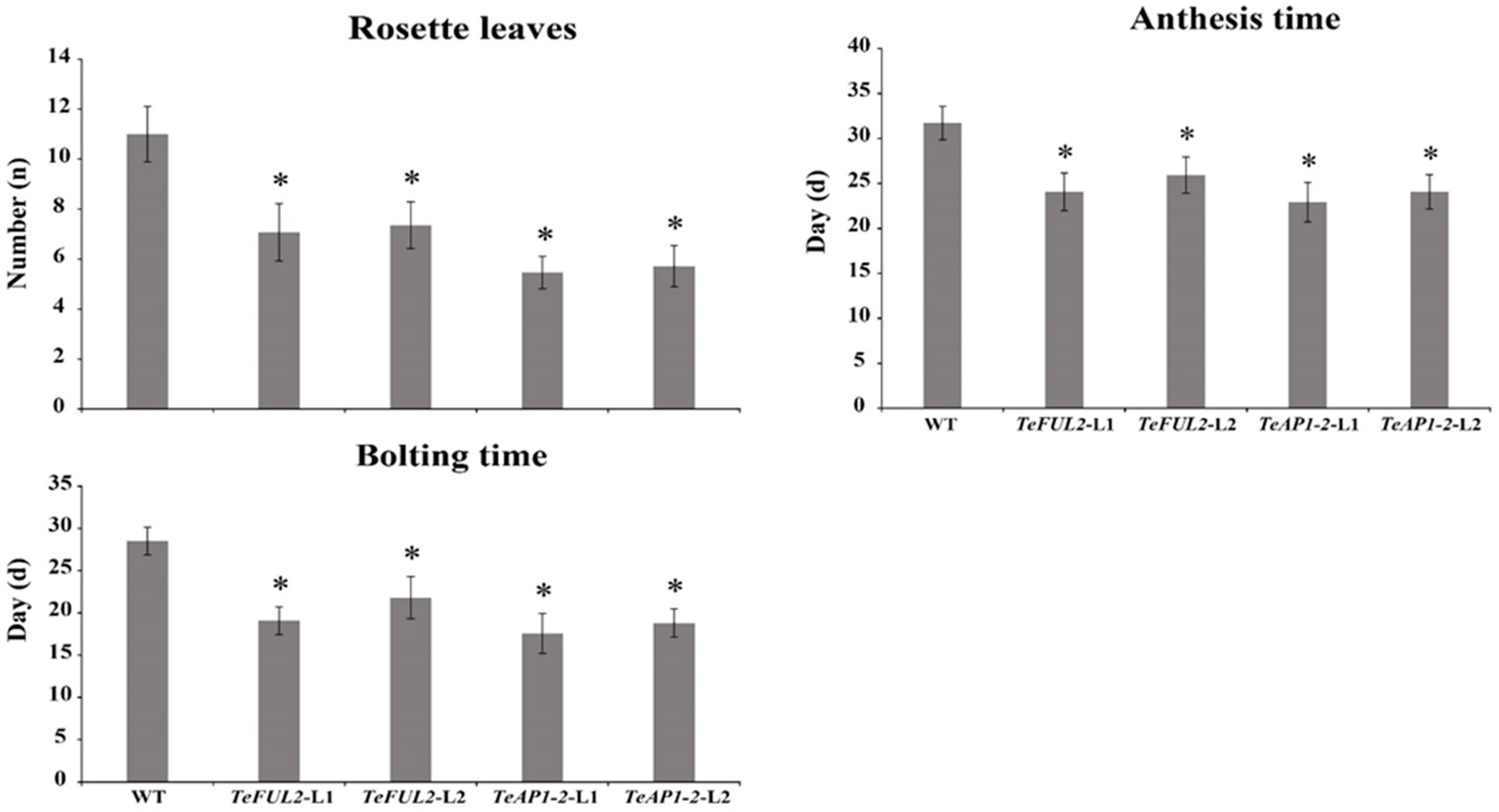
Compared with the wild-type plants, overexpression of TeAP1-2 and TeFUL2 in Arabidopsis caused obvious early flowering (Figure 4d,e and Figure 5). According to the statistics, wild-type Arabidopsis flowered in ten to thirteen (11.17 ± 1.11) rosette leaves, while most transgenic lines flowered in five to eight rosette leaves under the same conditions (Figure 5). In addition, ectopic expression of TeAP1-2 also led to the wavy shape of the last two rosette leaves and the curling of cauline leaves (Figure 4a–c). However, overexpression of TeAP1-2 and TeFUL2 in Arabidopsis could not affect flower development (Figure 4f–k).
3.5. Expression Analysis of Endogenous Genes in Transgenic Plants
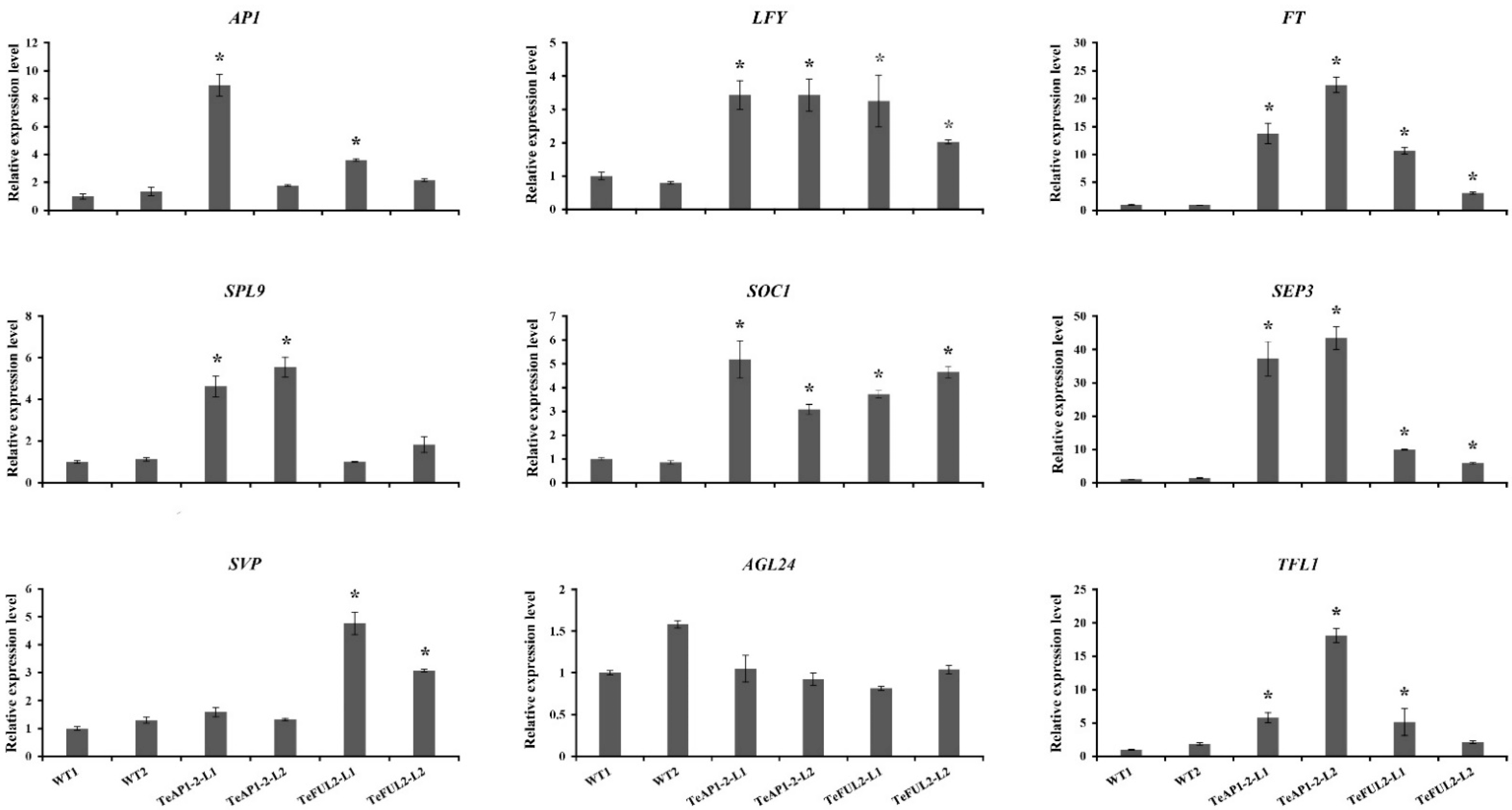
To reveal the mechanism underlying phenotypic changes of transgenic lines 35S:TeAP1-2 and 35S:TeFUL2, the expression levels of AP1-regulated endogenous genes were analyzed when the T3 transgenic and wild-type seedlings were 10 days old. As shown in Figure 6, TeAP1-2 and TeFUL2 displayed a similar function in regulating the expression level of AP1 downstream genes. For example, the expressions of AP1, FT, LFY, SOC1, SPE3, and TFL1 in transgenic lines 35S:TeAP1-2 and 35S:TeFUL2 were obviously higher than those in wild-type plants. The expression level of AGL24 showed no remarkable changes in both transgenic lines 35S:AP1-2 and 35S:FUL2. It should be noted that TeAP1-2 and TeFUL2 specifically regulated some downstream genes. For instance, SPL9 was significantly upregulated in transgenic lines 35S:TeAP1-2, while it exhibited no change in transgenic lines 35S:FUL2. In contrast to SPL9, SVP was significantly highly expressed in transgenic lines 35S:FUL2, but there was no significant change in transgenic lines 35S:AP1-2.
4. Discussion
The study of many AP1/FUL-like genes from various species has demonstrated that AP1/FUL genes play key roles in flowering time, flower and fruit development. Like APETALLA3 (AP3, B class gene) and AGAMOUS (AG, C class gene), the AP1/FUL genes underwent several duplication events, resulting in the occurrence of euAP1 and euFUL clade in core eudicots [13,14]. In this study, five marigold AP1/FUL-like genes were obtained. Sequence alignment analysis indicated that all these five TeAP1/FUL-like proteins were typical MIKC proteins, and they contained a conserved motif at their C terminal domain (Figure 1). TeAP1-1 and TeAP1-2 were clustered into euAP1 clade proteins harboring an acidic domain and a farnesylation motif (Figure 1), and the TeFUL1, TeFUL2, and TeFUL3 possessed a conserved FUL motif (Figure 1), which suggested these three marigold FUL-like proteins were members of FUL clade proteins [13,14]. Such changes in the 3′ end of coding sequence have been explained by a frameshift mutation in ancestral AP1/FUL-like genes [13,56] and are responsible for gene-specific functions.
Our phylogenetic analysis indicated that TeAP1-1 and TeAP1-2 were members of the AP1 clade, and they seemed to be homologous to Antirrhinum SQUA, which was previously reported to be involved in regulating the floral meristem development and specifying the sepal and petal identity [57]. TeFUL1, TeFUL2, and TeFUL3 were clustered into the FUL clade, and TeFUL1 and TeFUL3 proteins were close to the euFULI group. TeFUL2 belonged to the euFULII group (Figure 2). TeFUL2 was orthologous to the antirrhinum DEFH28 involved in regulating floral meristem development, fruit development, and flowering time [40]. Gene expression analysis indicated that TeFUL2 was mainly expressed at the early stage of inflorescence development (Figure 3 and Supplementary Figure S2), and the expression pattern of TeFUL2 was similar to that of the early function genes Arabidopsis FUL [23] and petunia PFG [42], implying a role of TeFUL2 in meristem identity. However, TeFUL1 and TeFUL3 were expressed in vegetative tissues, different stages of flower buds, and floral organs (Figure 3 and Supplementary Figure S2). Based on these findings, it could be speculated that TeFUL2 and TeFUL1 (or TeFUL3) might arise from gene duplication and that this duplication event might cause the change in their expression patterns. Many previous studies reveal that functional divergence is caused by gene duplication, which further drives evolution [10,58]. Therefore, we speculated that the duplication events and transcript pattern differences of TeFUL genes might imply the functional divergence of these genes in marigold.
4.1. Conserved Function of AP1/FUL Genes in Early Flowering
Functional analysis of the AP1/FUL-like genes in core eudicots and non-core eudicots reveal that AP1/FUL-like genes display conserved roles in regulating the flowering time. For example, overexpression of AP1 or FUL in Arabidopsis both leads to early flowering [29,59]. Furthermore, a similar phenotype is also observed in the case of ectopic overexpression of AP1-like or FUL-like genes from the Asteraceae species, such as C. morifolium (CDM111) [45], C. lavandulifolium (ClM8) [44] and G. hybrida (GSUQA2) [43]. In this study, heterologous expression of TeAP1-2 and TeFUL2 in Arabidopsis resulted in early flowering without affecting floral organ identity (Figure 4d–k and Figure 5). In addition, ectopic expression of TeAP1-2 also led to the wave-shaped rosette leaf and curled cauline leaf (Figure 4b,c), which was similar to the function of the AP1/FUL-like gene MBP20 [60]. The MADS-box transcription factors possess a DNA-binding domain to regulate their downstream gene expressions [58]. Therefore, we speculated that the early flowering phenotypes observed in 35S:TeAP1-2 and 35S:TeFUL2 transgenic lines might be related to the change in endogenous gene expression levels. In this study, AP1, FT, LFY, SOC1, and SEP3 were significantly upregulated in 10-day-old seedlings of transgenic lines containing 35S:TeAP1-2 and 35S:TeFUL2 fusion vectors (Figure 6), suggesting TeAP1-2 and TeFUL2 might share the overlapping regulation network of a series of downstream genes in Arabidopsis. Remarkably, the TFL1 was significantly activated in transgenic seedlings overexpressing TeAP1-2 or TeFUL2 (Figure 6), which was consistent with the previous report that overexpression of the FUL-like gene PlacFL2 from Platanus acerifolia Willd. obviously activated the TFL1 expression [59]. However, in Arabidopsis, the TFL1 inhibits AP1 activities through transcriptional repression [55,61]. Therefore, our results require to be further investigated. In Arabidopsis, AP1 directly represses SVP, AGL24, and SOC1 to partially specify floral meristem identities [62]. However, in our study, no remarkable change in the expression level of AGL24 was observed in transgenic lines 35S:TeAP1-2 and 35S:TeFUL2 (Figure 6). Additionally, the expression level of the flowering repressor gene SVP was significantly activated in transgenic lines 35S:TeFUL2, but not in transgenic lines 35S:TeAP1-2 (Figure 6). In contrast to SVP, SPL9 was significantly upregulated in transgenic lines 35S:TeAP1-2, but not in transgenic lines 35S:TeFUL2 (Figure 6). These results revealed that TeAP1-2 and TeFUL2 had divergent functions in regulating downstream genes, which was further supported by their difference in protein interaction patterns (Table 1, Supplementary Figure S3b–e).
4.2. Potential Redundant Function of TeAP1-1 and TeAP1-2 as Class A Genes
In Arabidopsis, AP1 is an early-acting gene, and it functions as a class A gene to specify sepal and petal identity [22,63]. AP1 is expressed in floral meristems and developing sepal and petal primordia [22,23,26,64]. However, in other core eudicots, the AP1-like genes can also be expressed in bracts and reproductive organs [31,45,65,66]. Similarly, both TeAP1-1 and TeAP1-2 were highly expressed in sepals of two-type florets and petals of disk florets, bracts, receptacles, and ovaries (Figure 3 and Supplementary Figure S2). Previous studies have revealed that the AP1 gene is involved in the specification of floral meristem (FM) identity and its high expression in inflorescence meristems and inflorescence branch meristem of Cornus species tends to form closed inflorescences [22,25,67,68]. In this study, TeAP1-2 was relatively highly expressed in flower buds, implying TeAP1-2 might regulate the head flower formation. According to the floral quartet model, the combinations of class A and E proteins specify the sepal identity [5,69]. In this study, TeAP1-1 and TeAP1-2 shared a similar protein interaction pattern to form heterodimers with TeSEP3-2 and TeSEP3-3 (Table 1, Supplementary Figure S3b,e). In Arabidopsis, AP1 only interacted with SEP to form a heterodimer. Furthermore, in the Asteraceae species, the AP1-like proteins C. morifolium CDM111 [45,46], G. hybrida GSQUA1, and GSQUA3 [70] also had a limited protein interaction pattern. In other words, they only interacted with SEP proteins to form heterodimers. Additionally, the results of protein-protein interaction also imply that the TeSEP3 proteins played a glue role in regulating floral organ development. Taken together, as class A genes, TeAP1-1 and TeAP1-2 might play a redundant role.
4.3. Divergent Functions among TeFULs Genes
It is well-known that FUL-like genes play important roles in the transition from vegetative meristems to reproductive meristems and in fruit development in many core eudicots and non-core eudicots. In the model plant Arabidopsis, FUL regulates the cell differentiation during fruit development [37,39,61] and participates in specifying floral meristem identity together with AP1 and CAL [23]. In basal eudicots, the Aquilegia coerulea Pall. FUL-like genes regulate leaf morphogenesis and inflorescence development [8]. Additionally, in monocots, Oryza sativa L. homologs genes OsMADS14 and OsMADS15 are involved in specifying the meristem identity, palea and lodicule identity [7]. In contrast to the AP1-like genes, the FUL-like genes are widely expressed in vegetative and reproductive tissues [6,37,59].
In our study, TeFUL1 and TeFUL3 were expressed in stems and leaves as well as in reproductive tissues (Figure 3 and Supplementary Figure S2), which was in line with the typical FUL-like expression pattern [6,37,59], implying that TeFUL1 and TeFUL3 might play a role as FUL genes. Furthermore, ectopic expression of TeFUL1 or TeFUL3 in Arabidopsis led to no visible phenotype changes. In Arabidopsis, FUL functions redundantly with CAL and AP1 to specify the floral meristem identity, and single ful mutation has no ability to affect floral organ identity [23]. In general, we speculated that TeFUL1 and TeFUL3 might function redundantly in regulating the floral meristem identity, or that TeFUL1 and TeFUL3 need to work together with AP1-like genes to regulate the floral meristem development. However, the striking difference in the protein interaction pattern was observed between TeFUL1 and TeFUL3 (Table 1, Supplementary Figure S3b–e). TeFUL1 only interacted with TeAGL6, while TeFUL3 interacted with TeSEP1, TeSEP3-1, TeSEP3-2, TeSEP3-3, and TeSEP4 to form heterodimers (Table 1, Supplementary Figure S3b–e). Different protein interaction patterns might be related to their different conserved regions at C domains (Figure 1). The above results suggested that TeFUL1 and TeFUL3 might be partially functionally redundant, but they might have their own specific functions in regulating floral organ identity.
In contrast to TeFUL1 and TeFUL3, TeFUL2 was highly expressed in flower buds and vegetative tissues, and weakly expressed or unexpressed in floral organs and ovules (Figure 3 and Supplementary Figure S2). Additionally, TeFUL2 could form a homodimer by itself; meanwhile, it could form heterodimers with TeAGL6 and TeSEP3-2 (Table 1, Supplementary Figure S3b,e). Ectopic expression of TeFUL2 in Arabidopsis also led to early flowering with fewer rosette leaves (Figure 5), which was consistent with the phenotype of the plants overexpressing euFULII (DEFH28) clade genes from core eudicots and non-core eudicots [34,43]. The above results suggested that TeFUL1 and TeFUL3 might lose some functions, but these functions might have been retained in TeFUL2. Overexpression of Antirrhinum DEFH28 (euFULII clade genes) in Arabidopsis resulted in early flowering, two to four carpel formations, and failure of silique dehiscence [34]. However, ectopic expression of TeFUL2 in Arabidopsis did not affect floral organ identity and silique dehiscence (Figure 4), which was in line with the study results of G. hybrida GSQUA2 [43]. In general, TeFUL2 might retain a conserved role in regulating the meristem transition rather than fruit ripping.
5. Conclusions
In conclusion, marigold has five AP1/FUL-like genes, two of which are clustered into the euAP1 clade and three to the FUL-like clade. Based on the analyses of gene expression and protein interaction patterns, TeAP1-1 and TeAP1-2 are likely to play a redundant role in regulating sepal and petal identity, which is similar to the function of class A genes. Additionally, ectopic expression of TeAP1-2 resulted in early flowering, implying that TeAP1-2 might be involved in the regulation of floral transition. However, three FUL-like genes display divergent functions. TeFUL1 and TeFUL3 are more functionally close to euFUL genes, whereas TeFUL2 is more functionally close to antirrhinum DEFH28 belonging to the euFULII gene. Our results will provide a theoretical basis for the study of class A genes in Asteraceae. Considering the great difference in the florescence structure between marigold and Arabidopsis, this study will be helpful for understanding the function of AP1/FUL genes in Asteraceae species.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/genes12122011/s1, Figure S1: The map of pCAMBIA2300s, Figure S2: Expression levels of TeAP1/FUL in different tissues and organs, Figure S3: Interactions of TeAP1/FUL proteins with class B, C, D, and E proteins of marigold by yeast two-hybrid assays, Figure S4: Expression of TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 in seedlings of T1 transgenic plants, Table S1: Sequence of primers, Table S2: Sequence information of AP1/FUL-like proteins used to construct evolutionary tree, Table S3: Amino acid sequence alignment of TeAP1/FUL proteins.
Author Contributions
Conceived and designed the experiments: Y.H.; C.Z. Performed the experiments: C.Z. Analyzed the data: C.Z.; Y.S. Contributed reagents/materials/analysis tools: C.Z.; X.Y. Wrote the paper: C.Z.; Y.H. Plant cultivation: C.Z.; X.Y.; H.L. Revised the paper: C.Z.; Y.H.; M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from the National Natural Science Foundation of China (31672181), the National Key R&D Program of China (2018YFD1000400), and the Fundamental Research Funds for the Central Universities (2662019PY072).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy considerations.
Acknowledgments
The authors are grateful to all past and present colleagues in our lab for constructive discussion and technical support.
Conflicts of Interest
The authors declare that no competing commercial or financial interest exist.
Abbreviations
| AP1 | APETALA1 |
| FUL | FRUITFULL |
| AGL79 | AGAMOUS-like 79 |
| DEFH28 | DEFICIENS-homolog28 |
| PFG | PETUNIA FLOWERING GENE |
| qRT-PCR | quantitative real-time PCR |
| Semi-PCR | semi-quantitative PCR |
References
- Irish, V.F. The flowering of Arabidopsis flower development. Plant J. 2010, 61, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Meyerowitz, E.M.; Smyth, D.R.; Bowman, J.L. Abnormal flowers and pattern formation in floral. Development 1989, 106, 209–217. [Google Scholar] [CrossRef]
- Schwarzsommer, Z.; Huijser, P.; Nacken, W.; Saedler, H.; Sommer, H. Genetic Control of Flower Development by Homeotic Genes in Antirrhinum majus. Science 1990, 250, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31. [Google Scholar] [CrossRef]
- Theissen, G.; Saedler, H. Plant biology: Floral quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef]
- Wang, S.F.; Lu, G.; Hou, Z.; Luo, Z.D.; Wang, T.T.; Li, H.X.; Zhang, J.H.; Ye, Z.B. Members of the tomato FRUITFULL MADS-box family regulate style abscission and fruit ripening. J. Exp. Bot. 2014, 65, 3005–3014. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Shi, X.W.; Lin, X.L.; Liu, Y.; Chong, K.; Theisen, G.; Meng, Z. The ABCs of flower development: Mutational analysis of AP1/FUL-like genes in rice provides evidence for a homeotic (A)-function in grasses. Plant J. 2017, 89, 310–324. [Google Scholar] [CrossRef]
- Pabonmora, N.; Sharma, B.; Holappa, L.; Kramer, E.M.; Litt, A. The Aquilegia FRUITFULL-like genes play key roles in leaf morphogenesis and inflorescence development. Plant J. 2013, 74, 197–212. [Google Scholar] [CrossRef]
- Hands, P.; Vosnakis, N.; Betts, D.; Irish, V.F.; Drea, S. Alternate transcripts of a floral developmental regulator have both distinct and redundant functions in opium poppy. Ann. Bot. 2011, 107, 1557–1566. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.; Theissen, G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Martinezcastilla, L.P.; Alvarezbuylla, E.R. Adaptive evolution in the Arabidopsis MADS-box gene family inferred from its complete resolved phylogeny. Proc. Natl. Acad. Sci. USA 2003, 100, 13407–13412. [Google Scholar] [CrossRef] [Green Version]
- Hileman, L.C.; Sundstrom, J.F.; Litt, A.; Chen, M.; Shumba, T.; Irish, V.F. Molecular and Phylogenetic Analyses of the MADS-Box Gene Family in Tomato. Mol. Biol. Evol. 2006, 23, 2245–2258. [Google Scholar] [CrossRef] [PubMed]
- Litt, A.; Irish, V.F. Duplication and Diversification in the APETALA1/FRUITFULL Floral Homeotic Gene Lineage: Implications for the Evolution of Floral Development. Genetics 2003, 165, 821–833. [Google Scholar] [CrossRef]
- Shan, H.Y.; Zhang, N.; Liu, C.J.; Xu, G.X.; Zhang, J.; Chen, Z.D.; Kong, H.Z. Patterns of gene duplication and functional diversification during the evolution of the AP1/SQUA subfamily of plant MADS-box genes. Mol. Phylogenet. Evol. 2007, 44, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Litt, A.; Kramer, E.M. The ABC model and the diversification of floral organ identity. Semin. Cell Dev. Biol. 2010, 21, 129–137. [Google Scholar] [CrossRef]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Preston, J.C.; Kellogg, E.A. Reconstructing the evolutionary history of paralogous APETALA1/FRUITFULL-like genes in grasses (Poaceae). Genetics 2006, 174, 421–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, J.C.; Kellogg, E.A. Conservation and divergence of APETALA1/FRUITFULL-like gene function in grasses: Evidence from gene expression analyses. Plant J. 2007, 52, 69–81. [Google Scholar] [CrossRef]
- Lowman, A.C.; Purugganan, M.D. Duplication of the Brassica oleracea APETALA1 floral homeotic gene and the evolution of domesticated cauliflower. J. Hered. 1999, 90, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, E.R.; Garcia-Ponce, B.; Garay-Arroyo, A. Unique and redundant functional domains of APETALA1 and CAULIFLOWER, two recently duplicated Arabidopsis thaliana floral MADS-box genes. J. Exp. Bot. 2006, 57, 3099–3107. [Google Scholar] [CrossRef] [Green Version]
- Yalovsky, S.; Rodriguez-Concepcion, M.; Bracha, K.; Toledo-Ortiz, G.; Gruissem, W. Prenylation of the floral transcription factor APETALA1 modulates its function. Plant Cell 2000, 12, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Mandel, M.A.; Gustafsonbrown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, C.; Gu, Q.; Martienssen, R.A.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [CrossRef]
- Bowman, J.L.; Alvarez, J.P.; Weigel, D.; Meyerowitz, E.M.; Smyth, D.R. Control of flower development in Arabidopsis thaliana by APETALA1 and interacting genes. Development 1993, 119, 721–743. [Google Scholar] [CrossRef]
- Mandel, M.A.; Yanofsky, M.F. A gene triggering flower formation in Arabidopsis. Nature 1995, 377, 522–524. [Google Scholar] [CrossRef]
- Kempin, S.A.; Savidge, B.; Yanofsky, M.F. Molecular basis of the cauliflower phenotype in Arabidopsis. Science 1995, 267, 522–525. [Google Scholar] [CrossRef]
- Benlloch, R.; Derfurth, I.; Ferrandiz, C.; Cosson, V.; Beltran, J.P.; Canas, L.A.; Kondorosi, A.; Madueno, F.; Ratet, P. Isolation of mtpim Proves Tnt1 a Useful Reverse Genetics Tool in Medicago truncatula and Uncovers New Aspects of AP1-like Functions in Legumes. Plant Physiol. 2006, 142, 972–983. [Google Scholar] [CrossRef] [Green Version]
- Vrebalov, J.; Ruezinsky, D.; Padmanabhan, V.; White, R.; Medrano, D.; Drake, R.; Schuch, W.; Giovannoni, J.J. A MADS-Box Gene Necessary for Fruit Ripening at the Tomato Ripening-Inhibitor (Rin) Locus. Science 2002, 296, 343–346. [Google Scholar] [CrossRef]
- Duan, Y.; Fan, J.; Guo, W. Regeneration and characterization of transgenic kumquat plants containing the Arabidopsis APETALA1 gene. Plant Cell Tiss. Org. 2010, 100, 273–281. [Google Scholar] [CrossRef]
- Elo, A.; Lemmetyinen, J.; Turunen, M.; Tikka, L.; Sopanen, T. Three MADS-box genes similar to APETALA1 and FRUITFULL from silver birch (Betula pendula). Physiol. Plant. 2001, 112, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Berbel, A.; Navarro, C.; Ferrandiz, C.; Canas, L.A.; Madueno, F.; Beltran, J.P. Analysis of PEAM4, the pea AP1 functional homologue, supports a model for AP1-like genes controlling both floral meristem and floral organ identity in different plant species. Plant J. 2001, 25, 441–451. [Google Scholar] [CrossRef]
- Shannon, S.; Meeks-Wagner, D.R. A mutation in the Arabidopsis TFL1 gene affects inflorescence meristem development. Plant Cell 1991, 3, 877–892. [Google Scholar] [CrossRef]
- Schultz, E.A.; Haughn, G.W. LEAFY, a homeotic gene that regulates inflorescence development in Arabidopsis. Plant Cell 1991, 3, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bodt, S.; Raes, J.; Florquin, K.; Rombauts, S.; Rouze, P.; Theisen, G.; De Peer, Y.V. Genomewide Structural Annotation and Evolutionary Analysis of the Type I MADS-Box Genes in Plants. J. Mol. Evol. 2003, 56, 573–586. [Google Scholar] [CrossRef]
- Parenicova, L.; De Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.; Ingram, R.; Kater, M.M.; Davies, B. Molecular and Phylogenetic Analyses of the Complete MADS-Box Transcription Factor Family in Arabidopsis: New Openings to the MADS World. Plant Cell 2003, 15, 1538–1551. [Google Scholar] [CrossRef] [Green Version]
- Blazquez, M.A.; Ferrandiz, C.; Madueno, F.; Parcy, F. How Floral Meristems are Built. Plant Mol. Biol. 2006, 60, 855–870. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Ferrandiz, C.; Yanofsky, M.F.; Martienssen, R.A. The FRUITFULL MADS-box gene mediates cell differentiation during Arabidopsis fruit development. Development 1998, 125, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Melzer, S.; Lens, F.; Gennen, J.; Vanneste, S.; Rohde, A.; Beeckman, T. Flowering-time genes modulate meristem determinacy and growth form in Arabidopsis thaliana. Nat. Genet. 2008, 40, 1489–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrandiz, C. Regulation of fruit dehiscence in Arabidopsis. J. Exp. Bot. 2002, 53, 2031–2038. [Google Scholar] [CrossRef] [Green Version]
- Muller, B.M.; Saedler, H.; Zachgo, S. The MADS-box gene DEFH28 from Antirrhinum is involved in the regulation of floral meristem identity and fruit development. Plant J. 2001, 28, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smykal, P.; Gennen, J.; De Bodt, S.; Ranganath, V.; Melzer, S. Flowering of strict photoperiodic Nicotiana varieties in non-inductive conditions by transgenic approaches. Plant Mol. Biol. 2007, 65, 233–242. [Google Scholar] [CrossRef]
- Immink, R.G.H.; Hannapel, D.J.; Ferrario, S.; Busscher, M.; Franken, J.; Campagne, M.M.L.; Angenent, G.C. A petunia MADS box gene involved in the transition from vegetative to reproductive development. Development 1999, 126, 5117–5126. [Google Scholar] [CrossRef] [PubMed]
- Ruokolainen, S.; Ng, Y.P.; Broholm, S.K.; Albert, V.A.; Elomaa, P.; Teeri, T.H. Characterization of SQUAMOSA-like genes in Gerbera hybrida, including one involved in reproductive transition. BMC Plant Biol. 2010, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, Y.P.; Fu, J.X.; Qi, S.; Ma, H.Z.; Dai, S.L. Isolation and functional analysis of the ClM8-FRUITFULL-like MADS-box gene from Chrysanthemum lavandulifolium. Sci. Hortic. 2013, 161, 125–133. [Google Scholar] [CrossRef]
- Shchennikova, A.V.; Shulga, O.A.; Immink, R.; Skryabin, K.G.; Angenent, G.C. Identification and characterization of four chrysanthemum MADS-box genes, belonging to the APETALA1/FRUITFULL and SEPALLATA3 subfamilies. Plant Physiol. 2004, 134, 1632–1641. [Google Scholar] [CrossRef] [Green Version]
- Goloveshkina, E.N.; Shchennikova, A.V.; Kamionskaya, A.M.; Skryabin, K.G.; Shulga, O.A. Influence of ectopic expression of Asteraceae MADS box genes on plant ontogeny in tobacco. Plant Cell Tiss.Org. 2012, 109, 61–71. [Google Scholar] [CrossRef]
- Yu, D.; Kotilainen, M.; Pollanen, E.; Mehto, M.; Elomaa, P.; Helariutta, Y.; Albert, V.A.; Teeri, T.H. Organ identity genes and modified patterns of flower development in Gerbera hybrida (Asteraceae). Plant J. 1999, 17, 51–62. [Google Scholar] [CrossRef]
- Mandel, J.R.; Dikow, R.B.; Siniscalchi, C.M.; Thapa, R.; Watson, L.E.; Funk, V.A. A fully resolved backbone phylogeny reveals numerous dispersals and explosive diversifications throughout the history of Asteraceae. Proc. Natl. Acad. Sci. USA 2019, 116, 14083–14088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, Y.; Zhang, C.L.; Sun, Y.L.; Wang, W.N.; He, Y.H.; Bao, M.Z. Characterization and functional analysis of five MADS-box B class genes related to floral organ identification in Tagetes erecta. PLoS ONE 2017, 12, e0169777. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wei, L.D.; Wang, W.J.; Qi, W.Q.; Cao, Z.; Li, H.; Bao, M.Z.; He, Y.H. Identification, characterization and functional analysis of AGAMOUS subfamily genes associated with floral organs and seed development in Marigold (Tagetes erecta). BMC Plant Biol. 2020, 20, 439. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wei, L.D.; Yu, X.M.; Li, H.; Wang, W.J.; Wu, S.Z.; Duan, F.; Bao, M.Z.; Chan, Z.L.; He, Y.H. Functional conservation and divergence of SEPALLATA-like genes in the development of two-type florets in marigold. Plant Sci. 2021, 110938. [Google Scholar] [CrossRef]
- He, Y.H.; Ning, G.G.; Sun, Y.L.; Hu, Y.; Zhao, X.Y.; Bao, M.Z. Cytological and mapping analysis of a novel male sterile type resulting from spontaneous floral organ homeotic conversion in marigold (Tagetes erecta L.). Mol. Breed. 2010, 26, 19–29. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, K.; Wellmer, F.; Muino, J.M.; Ferrier, T.; Wuest, S.E.; Kumar, V.; Serranomislata, A.; Madueno, F.; Krajewski, P.; Meyerowitz, E.M. Orchestration of Floral Initiation by APETALA1. Science 2010, 328, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbussche, M.; Theissen, G.; De Peer, Y.V.; Gerats, T. Structural diversification and neo-functionalization during floral MADS-box gene evolution by C-terminal frameshift mutations. Nucleic Acids Res. 2003, 31, 4401–4409. [Google Scholar] [CrossRef]
- Huijser, P.; Klein, J.; Lonnig, W.; Meijer, H.; Saedler, H.; Sommer, H. Bracteomania, an inflorescence anomaly, is caused by the loss of function of the MADS-box gene squamosa in Antirrhinum majus. EMBO J. 1992, 11, 1239–1249. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.H.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.S.; Lu, S.J.; Yi, S.S.; Han, H.J.; Zhou, Q.; Cai, F.F.; Bao, M.Z.; Liu, G.F. Identification and characterization of FRUITFULL-like genes from Platanus acerifolia, a basal eudicot tree. Plant Sci. 2019, 280, 206–218. [Google Scholar] [CrossRef]
- Burko, Y.; Shleizerburko, S.; Yanai, O.; Shwartz, I.; Zelnik, I.D.; Jacobhirsch, J.; Kela, I.; Eshedwilliams, L.; Ori, N. A Role for APETALA1/FRUITFULL Transcription Factors in Tomato Leaf Development. Plant Cell 2013, 25, 2070–2083. [Google Scholar] [CrossRef] [Green Version]
- Liljegren, S.J.; Gustafson-Brown, C.; Pinyopich, A.; Ditta, G.S.; Yanofsky, M.F. Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate. Plant Cell 1999, 11, 1007–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhou, J.; Brachadrori, K.; Yalovsky, S.; Ito, T.; Yu, H. Specification of Arabidopsis floral meristem identity by repression of flowering time genes. Development 2007, 134, 1901–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrandiz, C.; Liljegren, S.J.; Yanofsky, M.F. Negative Regulation of the SHATTERPROOF Genes by FRUITFULL during Arabidopsis Fruit Development. Science 2000, 289, 436–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, M.A.; Koornneef, M.; Putterill, J. Flowering on time: Genes that regulate the floral transition: Workshop on the molecular basis of flowering time control. EMBO Rep. 2001, 2, 1078–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sather, D.N.; Golenberg, E.M. Duplication of AP1 within the Spinacia oleracea L. AP1/FUL clade is followed by rapid amino acid and regulatory evolution. Planta 2009, 229, 507–521. [Google Scholar] [PubMed]
- Hernandes-Lopes, J.; Sousa-Baena, M.S.; Lemos, R.C.C.; Correa, T.C.S.; Van Sluys, M.A.; Melo-de-Pinna, G.F.A. Toward understanding inflorescence development and architecture in Passiflora: Insights from comparative anatomy and expression of APETALA1. Am. J. Bot. 2019, 106, 1173–1189. [Google Scholar] [CrossRef]
- Irish, V.F.; Sussex, I.M. Function of the apetala-1 gene during Arabidopsis floral development. Plant Cell 1990, 2, 741–753. [Google Scholar]
- Ma, Q.; Liu, X.; Franks, R.G.; Xiang, Q.J. Alterations of CorTFL1 and CorAP1 expression correlate with major evolutionary shifts of inflorescence architecture in Cornus (Cornaceae)—A proposed model for variation of closed inflorescence forms. New Phytol. 2017, 216, 519–535. [Google Scholar] [CrossRef]
- Theissen, G. Development of floral organ identity: Stories from the MADS house. Curr. Opin. Plant Biol. 2001, 4, 75–85. [Google Scholar] [CrossRef]
- Ruokolainen, S.; Ng, Y.P.; Albert, V.A.; Elomaa, P.; Teeri, T.H. Large scale interaction analysis predicts that the Gerbera hybrida floral E function is provided both by general and specialized proteins. BMC Plant Biol. 2010, 10, 129. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Alignment of marigold AP1/FUL-like amino acid sequence from model plants (Arabidopsis and Antirrhinum) and Asteraceae species. The MADS domain is marked with a bold red line. The I domain is marked with a bold black line. The K domain is marked with a bold yellow line. The C domain is marked with a bold blue line. The FUL protein motif is marked with a red box. EuAP1-like proteins contain both an acidic domain (shown in yellow dotted box), and a farnesylation motif (shown in blue dotted box) at their C termini. The whole domain marked with a red dotted line represents the euAP1 motif.
Figure 1.
Alignment of marigold AP1/FUL-like amino acid sequence from model plants (Arabidopsis and Antirrhinum) and Asteraceae species. The MADS domain is marked with a bold red line. The I domain is marked with a bold black line. The K domain is marked with a bold yellow line. The C domain is marked with a bold blue line. The FUL protein motif is marked with a red box. EuAP1-like proteins contain both an acidic domain (shown in yellow dotted box), and a farnesylation motif (shown in blue dotted box) at their C termini. The whole domain marked with a red dotted line represents the euAP1 motif.

Figure 2.
Phylogenetic tree based on the amino-acid alignment of TeAP1/FUL proteins. The tree was generated with the MEGA v7.0 software, using the neighbor-joining (NJ) method and 1000 bootstrap replicates. The TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 are marked with black triangles.
Figure 2.
Phylogenetic tree based on the amino-acid alignment of TeAP1/FUL proteins. The tree was generated with the MEGA v7.0 software, using the neighbor-joining (NJ) method and 1000 bootstrap replicates. The TeAP1-1, TeAP1-2, TeFUL1, TeFUL2, and TeFUL3 are marked with black triangles.

Figure 3.
Expression levels of TeAP1/FUL genes in different tissues and organs of marigold. (a) Heatmap of relative expression of TeAP1/FUL genes by qRT-PCR in different tissues and organs. Rt: root; Sm: stems; Le: leaves; FB1-FB4: flower buds were 0–1, 2–3, 4–5 and 6–7 mm in diameter, respectively; Re: receptacle; Br: bract; RS: sepal of ray floret; RP: petal of ray floret; RPi: pistil of ray floret; Se: sepal of disk floret; Pe: petal of disk floret; St: stamen of disk floret; Pi: pistil of disk floret; Ov: ovary. (b) Heatmap of TeAP1/FUL genes in the inflorescence of marigold based on the relative expression by qRT-PCR. Blank control: structural model of capitulum in marigold; different colors represent different floral organs.
Figure 3.
Expression levels of TeAP1/FUL genes in different tissues and organs of marigold. (a) Heatmap of relative expression of TeAP1/FUL genes by qRT-PCR in different tissues and organs. Rt: root; Sm: stems; Le: leaves; FB1-FB4: flower buds were 0–1, 2–3, 4–5 and 6–7 mm in diameter, respectively; Re: receptacle; Br: bract; RS: sepal of ray floret; RP: petal of ray floret; RPi: pistil of ray floret; Se: sepal of disk floret; Pe: petal of disk floret; St: stamen of disk floret; Pi: pistil of disk floret; Ov: ovary. (b) Heatmap of TeAP1/FUL genes in the inflorescence of marigold based on the relative expression by qRT-PCR. Blank control: structural model of capitulum in marigold; different colors represent different floral organs.

Figure 4.
Abnormal morphology of transgenic Arabidopsis plants of constitutively expressed TeAP1-2 and TeFUL2 genes. (a) The wild-type seedling. (b) The transgenic seedlings with severely curled rosette leave in 35S:TeAP1-2 transgenic lines. (c) The curled cauline leaves of 35S:TeAP1-2 transgenic lines. (d) Wild-type (left) and early flowering transgenic plant (right) of 35S:TeAP1-2 transgenic lines; WT: wild-type line 1; L1: 35S:TeAP1-2 line 1; L2: 35S:TeAP1-2 line 2; L3: 35S:TeAP1-2 line 3. (e) Wild-type (left) and early flowering transgenic plant (right) of 35S:TeFUL2 transgenic lines; WT: wild-type line 1; L1: 35S:TeFUL2 line 1; L2: 35S:TeFUL2 line 2; L3: 5S:TeFUL2 line 3. (f) The flower of wild-type Arabidopsis plants. (g) The anatomy of wild-type Arabidopsis flower. (h) The flower of 35S:TeAP1-2 transgenic plants. (i) The anatomy of 35S:TeAP1-2 transgenic plant flowers. (j) The flower of 35S:TeFUL2 transgenic plants. (k) The anatomy of 35S:TeFUL2 transgenic plant flowers. (a–e), bar = 5 mm; (f–k), bar = 500 μm. (l) Expression of TeAP1-2 in seedlings of T1 transgenic plants by semi-RT-PCR. The picture above is the expression level of TeAP1-2 in transgenic lines, the band size was 106 bp; the picture below the constitutive gene is the Arabidopsis keeping-house gene AtEF1α. (m) Expression of TeFUL2 in seedlings of T1 transgenic plants by semi-RT-PCR. The picture above is the expression level of TeFUL2 in transgenic lines, the band size was 127 bp; the picture below the constitutive gene is Arabidopsis house-keeping gene AtEF1α.
Figure 4.
Abnormal morphology of transgenic Arabidopsis plants of constitutively expressed TeAP1-2 and TeFUL2 genes. (a) The wild-type seedling. (b) The transgenic seedlings with severely curled rosette leave in 35S:TeAP1-2 transgenic lines. (c) The curled cauline leaves of 35S:TeAP1-2 transgenic lines. (d) Wild-type (left) and early flowering transgenic plant (right) of 35S:TeAP1-2 transgenic lines; WT: wild-type line 1; L1: 35S:TeAP1-2 line 1; L2: 35S:TeAP1-2 line 2; L3: 35S:TeAP1-2 line 3. (e) Wild-type (left) and early flowering transgenic plant (right) of 35S:TeFUL2 transgenic lines; WT: wild-type line 1; L1: 35S:TeFUL2 line 1; L2: 35S:TeFUL2 line 2; L3: 5S:TeFUL2 line 3. (f) The flower of wild-type Arabidopsis plants. (g) The anatomy of wild-type Arabidopsis flower. (h) The flower of 35S:TeAP1-2 transgenic plants. (i) The anatomy of 35S:TeAP1-2 transgenic plant flowers. (j) The flower of 35S:TeFUL2 transgenic plants. (k) The anatomy of 35S:TeFUL2 transgenic plant flowers. (a–e), bar = 5 mm; (f–k), bar = 500 μm. (l) Expression of TeAP1-2 in seedlings of T1 transgenic plants by semi-RT-PCR. The picture above is the expression level of TeAP1-2 in transgenic lines, the band size was 106 bp; the picture below the constitutive gene is the Arabidopsis keeping-house gene AtEF1α. (m) Expression of TeFUL2 in seedlings of T1 transgenic plants by semi-RT-PCR. The picture above is the expression level of TeFUL2 in transgenic lines, the band size was 127 bp; the picture below the constitutive gene is Arabidopsis house-keeping gene AtEF1α.

Figure 5.
Statistics for main morphological traits of the control and transgenic plants. * Significant difference at p < 0.05.
Figure 5.
Statistics for main morphological traits of the control and transgenic plants. * Significant difference at p < 0.05.

Figure 6.
qRT-PCR analysis of endogenous flowering related genes in 10-D-old seedlings of Arabidopsis wild-type and 35S:TeAP1-2 and 35S:TeFUL2 transgenic lines. WT1: wild-type line 1; WT2: wild-type line 2; * expression level of endogenous genes in transgenic plants was 2 times higher or 0.5 times lower than that in wild-type plants.
Figure 6.
qRT-PCR analysis of endogenous flowering related genes in 10-D-old seedlings of Arabidopsis wild-type and 35S:TeAP1-2 and 35S:TeFUL2 transgenic lines. WT1: wild-type line 1; WT2: wild-type line 2; * expression level of endogenous genes in transgenic plants was 2 times higher or 0.5 times lower than that in wild-type plants.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Interactions of Marigold TeAP1/FUL proteins with classes A, B, C, D, and E proteins detected by yeast two-hybrid assays.
Table 1.
Interactions of Marigold TeAP1/FUL proteins with classes A, B, C, D, and E proteins detected by yeast two-hybrid assays.
| AD BD | Class A Proteins | Class B Proteins | Class C Proteins | Class D Proteins | Class E Proteins | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TeAP1-1 | TeAP1-2 | TeFUL1 | TeFUL2 | TeFUL3 | TePI | TeAP3-1 | TeAP3-2 | TeTM6-1 | TeTM6-2 | TeAG1 | TeAG2 | TeAGL11-1 | TeAGL11-2 | TeSEP1 | TeSEP3-1 | TeSEP3-2 | TeSEP3-3 | TeSEP4 | TeAGL6 | |
| TeAP1-1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ++ | + | - | - |
| TeAP1-2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + | - | - |
| TeFUL1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + |
| TeFUL2 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + |
| TeFUL3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ++ | + | ++ | ++ | - | - |
| TePI | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAP3-1 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAP3-2 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeTM6-1 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeTM6-2 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAG1 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAG2 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAGL11-1 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAGL11-2 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeSEP1 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeSEP3-1 | - | - | - | - | + | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeSEP3-2 | + | - | - | + | + | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeSEP3-3 | - | - | - | - | + | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeSEP4 | - | - | - | - | ++ | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| TeAGL6 | - | - | - | - | - | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
Note: ++, strong interaction; +, weak interaction; -, no interaction; /, not tested.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, C.; Sun, Y.; Yu, X.; Li, H.; Bao, M.; He, Y. Functional Conservation and Divergence of Five AP1/FUL-like Genes in Marigold (Tagetes erecta L.). Genes 2021, 12, 2011. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122011
AMA Style
Zhang C, Sun Y, Yu X, Li H, Bao M, He Y. Functional Conservation and Divergence of Five AP1/FUL-like Genes in Marigold (Tagetes erecta L.). Genes. 2021; 12(12):2011. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122011
Chicago/Turabian StyleZhang, Chunling, Yalin Sun, Xiaomin Yu, Hang Li, Manzhu Bao, and Yanhong He. 2021. "Functional Conservation and Divergence of Five AP1/FUL-like Genes in Marigold (Tagetes erecta L.)" Genes 12, no. 12: 2011. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122011
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.