
Carotenoid Biosynthetic Genes in Cabbage: Genome-Wide Identification, Evolution, and Expression Analysis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Resource
2.2. Identification and Analysis of Orthologs between B. oleracea and A. thaliana
2.3. Non-Synonymous/Synonymous Substitution (Ka/Ks) Ratios of Gene Pairs among B. oleracea, A. thaliana, and B. rapa
2.4. Expression Analysis of Carotenoid Biosynthetic Genes in B. oleracea
2.5. Exogenous Phytohormone Spraying of Cabbage Leaves
2.6. Phylogenetic Analysis
2.7. Subcellular Localization of PSY in B. oleracea
3. Results
3.1. Identification of Carotenoid Biosynthetic Genes in B. oleracea
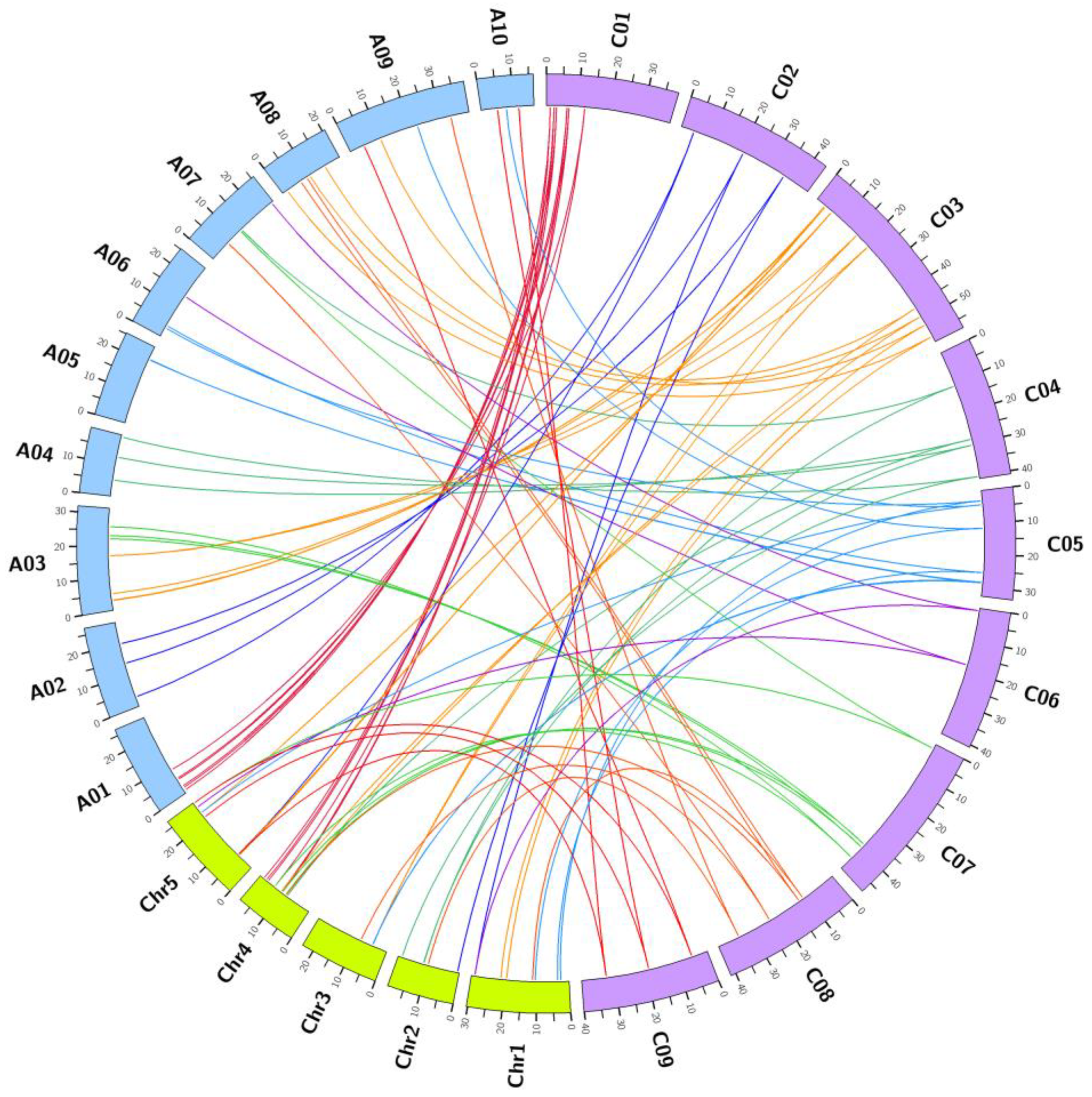
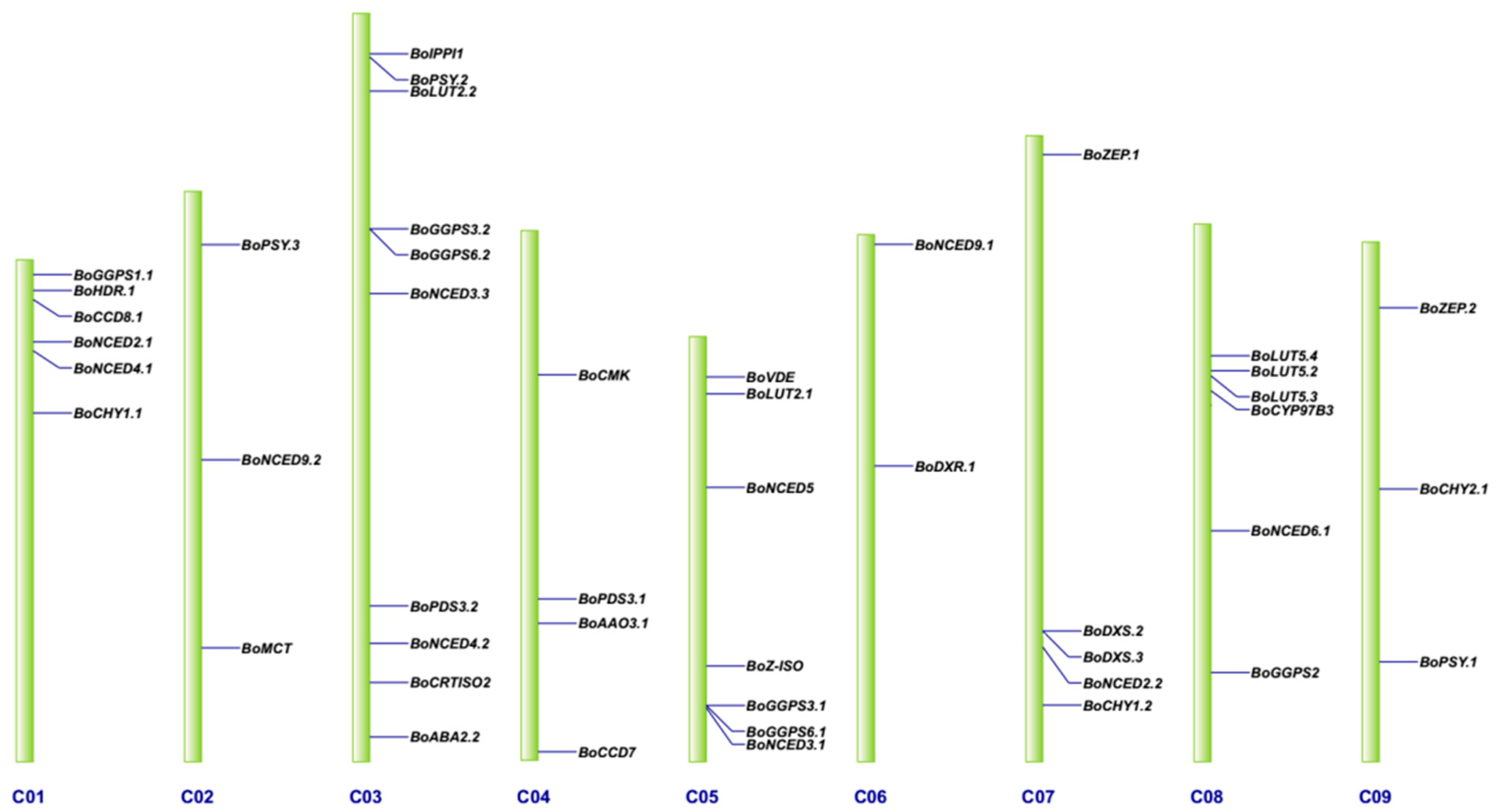
3.2. Genomic Distribution on Chromosomes
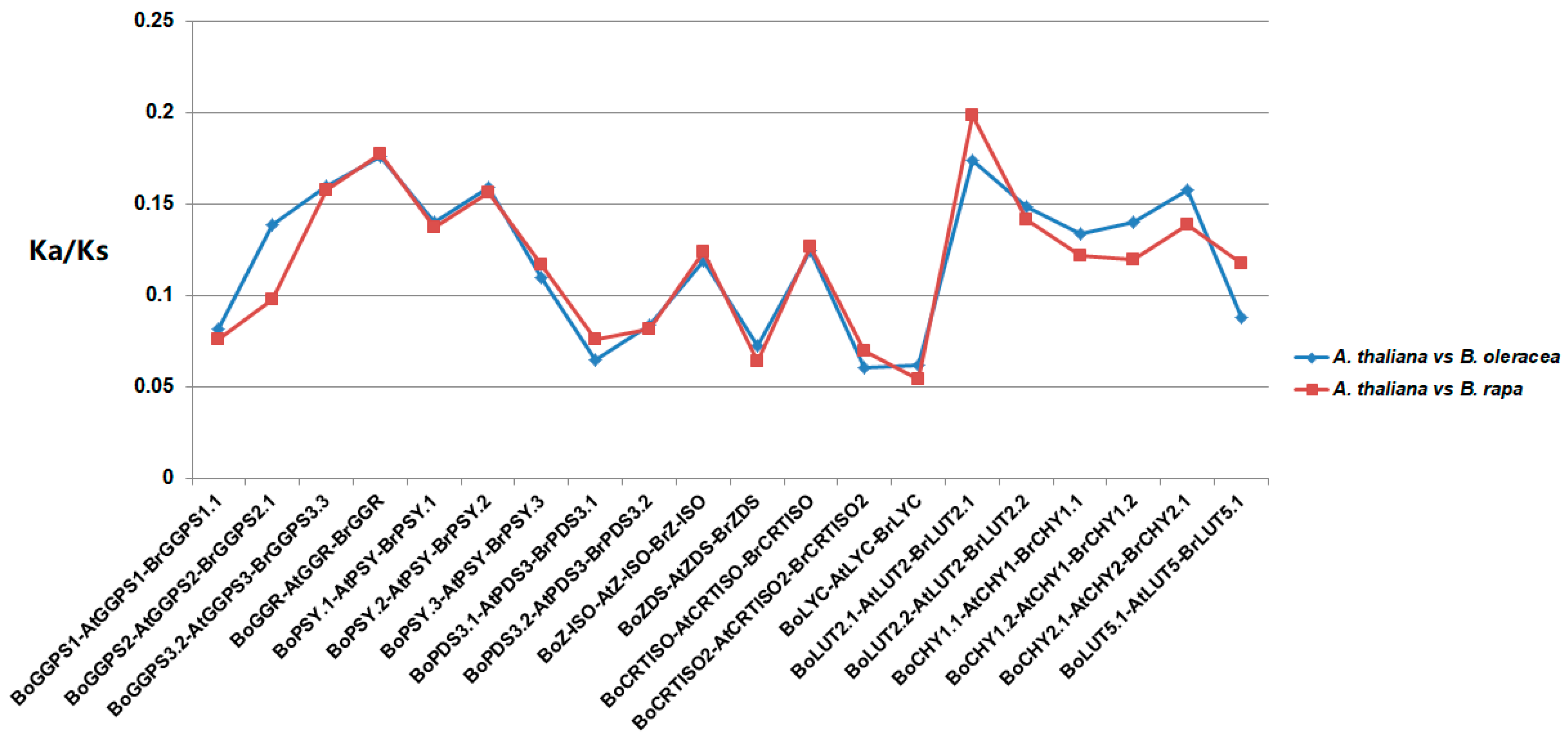
3.3. Comparative Evolutionary Analyses of Orthologous Gene Pairs for Carotenoid Biosynthetic Genes
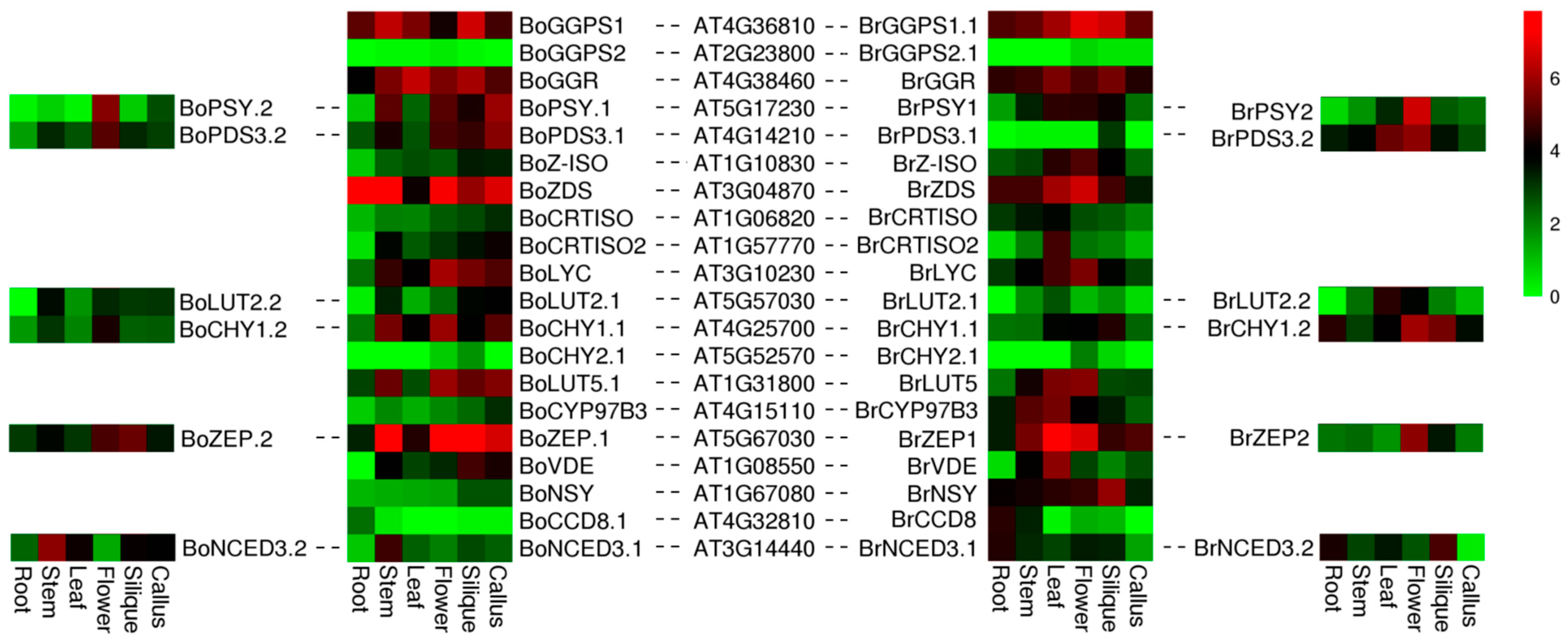
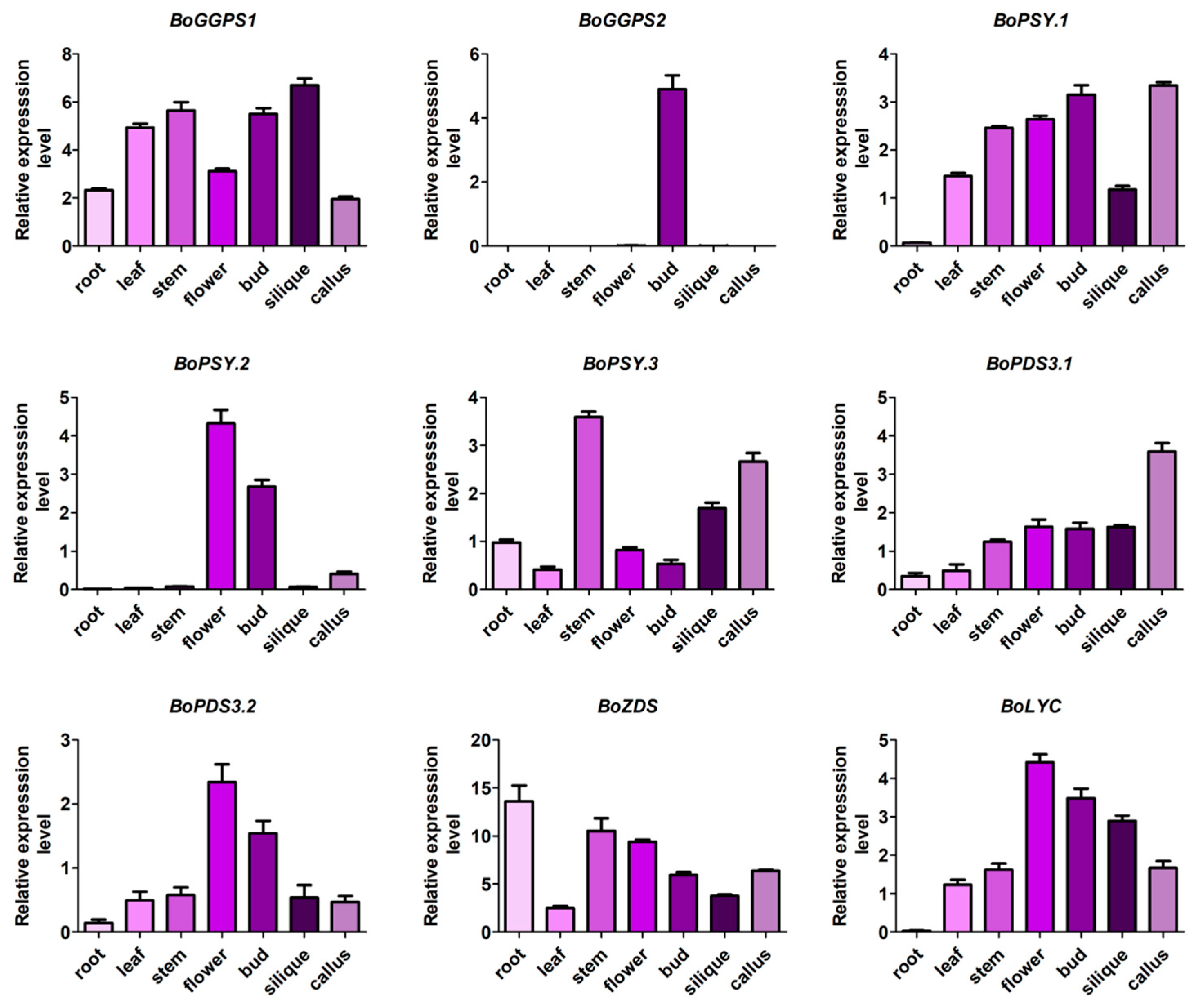
3.4. Expression Analysis of Orthologous for Carotenoid Biosynthetic Genes among B. oleracea, B. rapa and A. thaliana
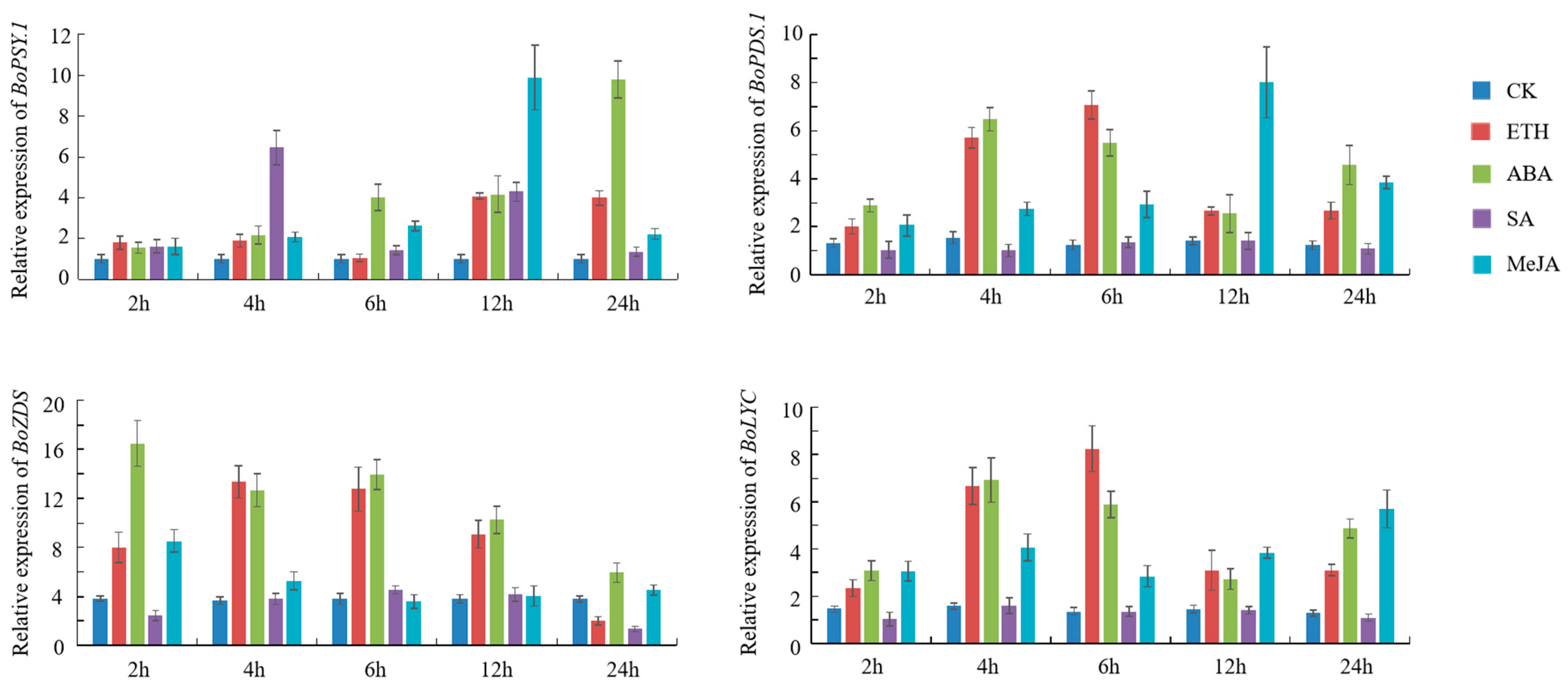
3.5. BoPSY.1, BoPDS.1, BoZDS, and BoLYC Respond to Exogenous Phytohormone Treatments
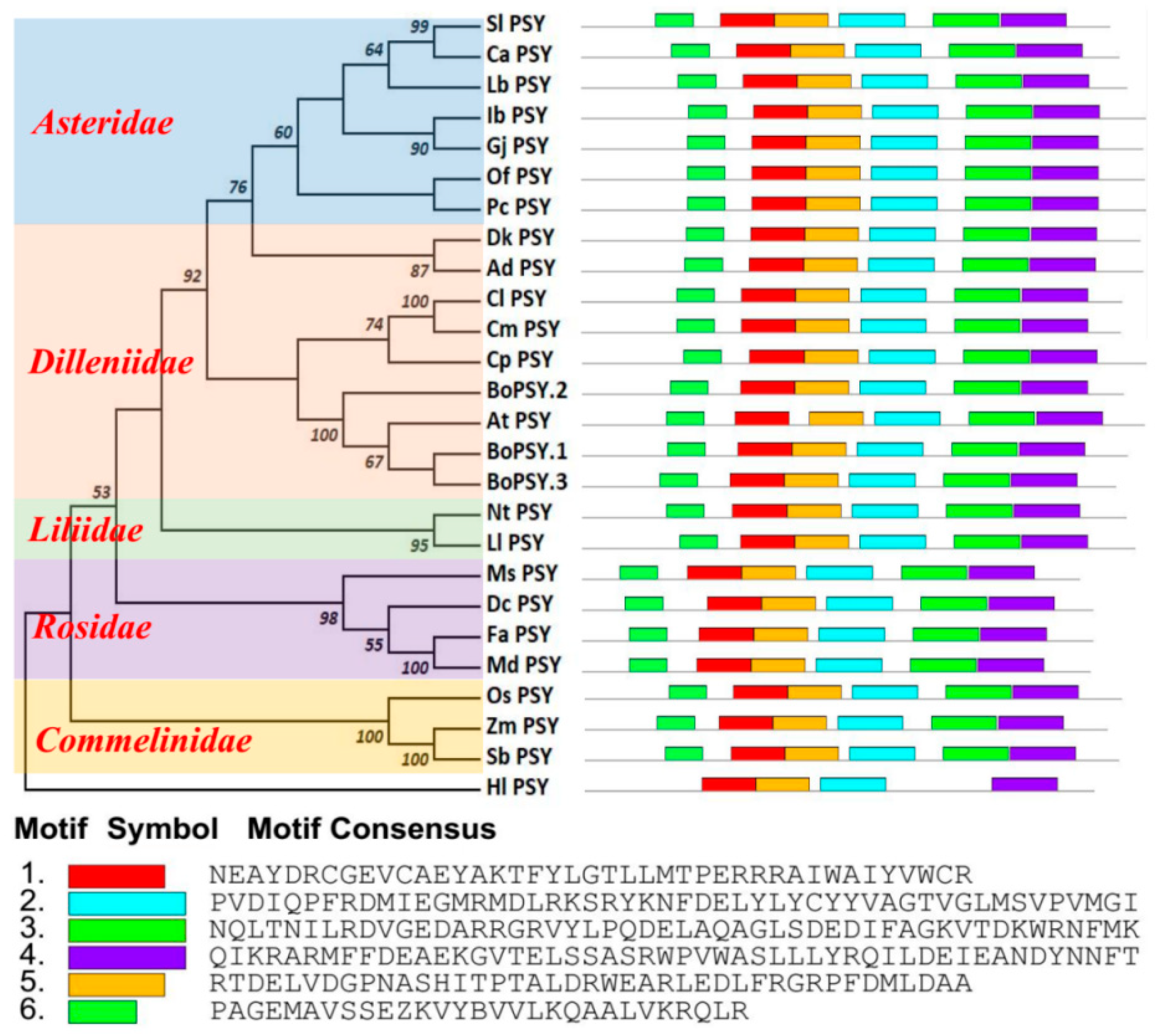
3.6. Evolution of PSY Genes
3.7. Subcellular Localization of BoPSY.1, BoPSY.2, and BoPSY.3
4. Discussion
4.1. Characterization of Carotenoid Biosynthetic Genes in B. oleracea
4.2. Exogenous Phytohormones Regulate the Expression of Carotenoid Biosynthetic Genes
4.3. Phytoene Synthase (PSY) Is a Key Enzyme in the Carotenoid Biosynthetic Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lv, H.; Wang, Y.; Han, F.; Ji, J.; Fang, Z.; Zhuang, M.; Li, Z.; Zhang, Y.; Yang, L. A high-quality reference genome for cabbage obtained with SMRT reveals novel genomic features and evolutionary characteristics. Sci. Rep. 2020, 10, 12394. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2007, 7, 213–229. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J. Red cabbage anthocyanins: Profile, isolation, identification, and antioxidant activity. Food Res. Int. 2013, 51, 303–309. [Google Scholar] [CrossRef]
- Alvarez, R.; Vaz, B.; Gronemeyer, H.; de Lera, A.R. Functions, therapeutic applications, and synthesis of retinoids and carotenoids. Chem. Rev. 2014, 114, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.Y.; Niu, J.; Shang, F.; Yang, L.; Chang, T.Y.; Wang, J.J. Characterization of the geranylgeranyl diphosphate synthase gene in Acyrthosiphon pisum (Hemiptera: Aphididae) and its association with carotenoid biosynthesis. Front. Physiol. 2019, 10, 1398. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Bernstein, P.S. Microbial xanthophylls. Appl. Microbiol. Biotechnol. 2005, 68, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Banos, M.; Garbayo, I.; Vilchez, C.; Bonete, M.J.; Martinez-Espinosa, R.M. Carotenoids from Haloarchaea and their potential in biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente, B.; Martinez-Garcia, J.F.; Stange, C.; Rodriguez-Concepcion, M. Illuminating colors: Regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49–55. [Google Scholar] [CrossRef]
- DellaPenna, D.; Pogson, B.J. Vitamin synthesis in plants: Tocopherols and carotenoids. Annu. Rev. Plant Biol. 2006, 57, 711–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassett, R.G.; Coombes, J.S. Astaxanthin in cardiovascular health and disease. Molecules 2012, 17, 2030–2048. [Google Scholar] [CrossRef] [PubMed]
- Mordente, A.; Guantario, B.; Meucci, E.; Silvestrini, A.; Lombardi, E.; Martorana, G.E.; Giardina, B.; Bohm, V. Lycopene and cardiovascular diseases: An update. Curr. Med. Chem. 2011, 18, 1146–1163. [Google Scholar] [CrossRef]
- Sommer, A.; Vyas, K.S. A global clinical view on vitamin A and carotenoids. Am. J. Clin. Nutr. 2012, 96, 1204S–1206S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.S.; Chaves, G.V.; Stenzel, A.P.; Pereira, S.E.; Saboya, C.J.; Ramalho, A. Improvement of anthropometric and biochemical, but not of vitamin A, status in adolescents who undergo Roux-en-Y gastric bypass: A 1-year follow up study. Surg. Obes. Relat. Dis. 2017, 13, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Carotenoids of biotechnological importance. In Biotechnology of Isoprenoids, Advances in Biochemical Engineering/Biotechnology; Schrader, J., Bohlmann, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 148, pp. 449–467. [Google Scholar] [CrossRef]
- Obulesu, M.; Dowlathabad, M.R.; Bramhachari, P.V. Carotenoids and Alzheimer’s disease: An insight into therapeutic role of retinoids in animal models. Neurochem. Int. 2011, 59, 535–541. [Google Scholar] [CrossRef]
- Eisenreich, W.; Bacher, A.; Arigoni, D.; Rohdich, F. Biosynthesis of isoprenoids via the non-mevalonate pathway. Cell. Mol. Life Sci. 2004, 61, 1401–1426. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Concepcion, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; O’Connell, A.P. Gene expression analysis of strawberry achene and receptacle maturation using DNA microarrays. J. Exp. Bot. 2002, 53, 2073–2087. [Google Scholar] [CrossRef] [Green Version]
- Scolnik, P.A.; Bartley, G.E. Nucleotide sequence of an Arabidopsis cDNA for phytoene synthase. Plant Physiol. 1994, 104, 1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsch, R.; Medina, J.; Giuliano, G.; Beyer, P.; Von Lintig, J. Structural and functional characterization of the phytoene synthase promoter from Arabidopsis thaliana. Planta 2003, 216, 523–534. [Google Scholar] [CrossRef]
- Naing, A.H.; Kyu, S.Y.; Pe, P.P.W.; Park, K.I.; Lee, J.M.; Lim, K.B.; Kim, C.K. Silencing of the phytoene desaturase (PDS) gene affects the expression of fruit-ripening genes in tomatoes. Plant Methods 2019, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Scolnik, P.A.; Bartley, G.E. Phytoene desaturase from Arabidopsis. Plant Physiol. 1993, 103, 1475. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, F.; Wurtzel, E.T. Isolation and characterization of the Z-ISO gene encoding a missing component of carotenoid biosynthesis in plants. Plant Physiol. 2010, 153, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kreunen, S.S.; Cuttriss, A.J.; DellaPenna, D.; Pogson, B.J. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis. Plant Cell 2002, 14, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, F.X.; Chamovitz, D.; Misawa, N.; Gantt, E.; Hirschberg, J. Cloning and functional expression in Escherichia coli of a cyanobacterial gene for lycopene cyclase, the enzyme that catalyzes the biosynthesis of β-carotene. FEBS Lett. 1993, 328, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.X.; Pogson, B.; Sun, Z.R.; McDonald, K.A.; DellaPenna, D.; Gantt, E. Functional analysis of the β and ε lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Lydiate, D.J.; Young, L.W.; Schafer, U.A.; Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by downregulating lycopene ε cyclase. Transgenic Res. 2008, 17, 573–585. [Google Scholar] [CrossRef]
- Gomez-Garcia Mdel, R.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol. 2008, 50, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G. Plant carotenoids: Genomics meets multi-gene engineering. Curr. Opin. Plant Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantini, E.; Falcone, G.; Frusciante, S.; Giliberto, L.; Giuliano, G. Dissection of tomato lycopene biosynthesis through virus-induced gene silencing. Plant Physiol. 2013, 163, 986–998. [Google Scholar] [CrossRef]
- Kachanovsky, D.E.; Filler, S.; Isaacson, T.; Hirschberg, J. Epistasis in tomato color mutations involves regulation of phytoene synthase 1 expression by cis-carotenoids. Proc. Natl. Acad. Sci. USA 2012, 109, 19021–19026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, P.D.; Römer, S.; Shipton, C.A.; Mills, P.B.; Kiano, J.W.; Misawa, N.; Drake, R.G.; Schuch, W.; Bramley, P.M. Evaluation of transgenic tomato plants expressing an additional phytoene synthase in a fruit-specifific manner. Proc. Natl. Acad. Sci. USA 2002, 99, 1092–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Xu, X.; Hu, X.; Liu, Y.; Cao, H.; Chan, H.; Gong, Z.; Yuan, Y.; Luo, Y.; Feng, B.; et al. SlMYB72 regulates the metabolism of chlorophylls, carotenoids, and flavonoids in tomato fruit. Plant Physiol. 2020, 183, 854–868. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Luo, D.; Lin, A.; Zhang, C.; Shan, L.; He, P.; Li, B.; Zhang, Q.; Hua, B.; Yuan, Z.; et al. A tomato B-box protein SlBBX20 modulates carotenoid biosynthesis by directly activating Phytoene Synthase 1, and is targeted for 26S proteasome-mediated degradation. New Phytol. 2019, 221, 279–294. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wang, X.; Zhang, X.; Guo, M.; Liu, T. Unraveling the target genes of RIN transcription factor during tomato fruit ripening and softening. J. Sci. Food Agric. 2017, 97, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Joung, J.G.; McQuinn, R.; Chung, M.Y.; Fei, Z.; Tieman, D.; Klee, H.; Giovannoni, J. Combined transcriptome, genetic diversity and metabolite profiling in tomato fruit reveals that the ethylene response factor SlERF6 plays an important role in ripening and carotenoid accumulation. Plant J. 2012, 70, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.H.; Mahoney, N.E.; Goodman, N.; Pavlath, A.E. Regulation of lycopene formation in cell suspension culture of VFNT tomato (Lycopersicon esculentum) by CPTA, growth regulators, sucrose, and temperature. J. Exp. Bot. 1995, 46, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, L.; Zhang, M.; Zhang, Y.; Wang, Q. Effect of carbon dioxide enrichment on health-promoting compounds and organoleptic properties of tomato fruits grown in greenhouse. Food Chem. 2014, 153, 157–163. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Hittinger, C.T.; Carroll, S.B. Gene duplication and the adaptive evolution of a classic genetic switch. Nature 2007, 449, 677–681. [Google Scholar] [CrossRef]
- Spillane, C.; Schmid, K.J.; Laoueille-Duprat, S.; Pien, S.; Escobar-Restrepo, J.M.; Baroux, C.; Gagliardini, V.; Page, D.R.; Wolfe, K.H.; Grossniklaus, U. Positive darwinian selection at the imprinted MEDEA locus in plants. Nature 2007, 448, 349–352. [Google Scholar] [CrossRef]
- Town, C.D.; Cheung, F.; Maiti, R.; Crabtree, J.; Haas, B.J.; Wortman, J.R.; Hine, E.E.; Althoff, R.; Arbogast, T.S.; Tallon, L.J.; et al. Comparative genomics of Brassica oleracea and Arabidopsis thaliana reveal gene loss, fragmentation, and dispersal after polyploidy. Plant Cell 2006, 18, 1348–1359. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome triplication found across the tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.Y.; Zhu, B.Z.; Zhu, H.L.; Zhang, Y.L.; Xie, Y.H.; Li, Y.C.; Luo, Y.B. Effect of suppression of ethylene biosynthesis on flavor products in tomato fruits. Russ. J. Plant Physiol. 2007, 54, 80–88. [Google Scholar] [CrossRef]
- Sun, L.; Yuan, B.; Zhang, M.; Wang, L.; Cui, M.; Wang, Q.; Leng, P. Fruit-specific RNAi-mediated suppression of SlNCED1 increases both lycopene and β-carotene contents in tomato fruit. J. Exp. Bot. 2012, 63, 3097–3108. [Google Scholar] [CrossRef]
- McQuinn, R.P.; Gapper, N.E.; Gray, A.G.; Zhong, S.; Tohge, T.; Fei, Z.; Fernie, A.R.; Giovannoni, J.J. Manipulation of ZDS in tomato exposes carotenoid- and ABA-specific effects on fruit development and ripening. Plant Biotechnol. J. 2020, 18, 2210–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.N.; Yang, Q.Q.; Kou, E.F.; Song, S.W.; Chen, R.Y.; Hao, Y.W. Effects of salicylic acid treatment on growth and quality of flower Chinese cabbage. North Hortic. 2020, 22, 53–59. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef] [PubMed]
- Welsch, R.; Arango, J.; Bar, C.; Salazar, B.; Al-Babili, S.; Beltran, J.; Chavarriaga, P.; Ceballos, H.; Tohme, J.; Beyer, P. Provitamin A accumulation in cassava (Manihot esculenta) roots driven by a single nucleotide polymorphism in a phytoene synthase gene. Plant Cell 2010, 22, 3348–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, D.; Arango, J.; Wust, F.; Beyer, P.; Welsch, R. Carotenoid crystal formation in Arabidopsis and carrot roots caused by increased phytoene synthase protein levels. PLoS ONE 2009, 4, e6373. [Google Scholar] [CrossRef] [Green Version]
- Shewmaker, C.; Sheehy, J.; Daley, M.; Colburn, S.; Ke, D. Seed-specifific overexpression of phytoene synthase: Increase in carotenoids and other metabolic effects. Plant J. 1999, 20, 401–412. [Google Scholar] [CrossRef]
- Li, P.; Zhang, S.; Zhang, S.; Li, F.; Zhang, H.; Cheng, F.; Wu, J.; Wang, X.; Sun, R. Carotenoid biosynthetic genes in Brassica rapa: Comparative genomic analysis, phylogenetic analysis, and expression profiling. BMC Genom. 2015, 16, 492. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Chai, Y.; Zhou, Y.; Yang, X.; Warburton, M.L.; Xu, S.; Cai, Y.; Zhang, D.; Li, J.; Yan, J. Natural variation in the sequence of PSY1 and frequency of favorable polymorphisms among tropical and temperate maize germplasm. Theor. Appl. Genet. 2013, 126, 923–935. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Arabidopsis thaliana | B. olerecea | |||
|---|---|---|---|---|---|
| Syntenic Orthologs | Non-Syntenic Orthologs | ||||
| LF | MF1 | MF2 | |||
| DXS | AT4G15560 | BoDXS.1 (Bol020307) | BoDXS.2 (Bol005061) | - | BoDXS.3 (Bol005062) |
| DXR | AT5G62790 | BoDXR.1 (Bol019430) | - | BoDXR.2 (Bol019181) | - |
| MCT | AT2G02500 | - | BoMCT (Bol015155) | - | - |
| CMK | AT2G26930 | BoCMK (Bol027780) | - | - | - |
| MDS | AT1G63970 | BoMDS.1 (Bol021045) | - | BoMDS.2 (Bol022634) | - |
| HDS | AT5G60600 | BoHDS (Bol003103) | - | - | - |
| HDR | AT4G34350 | BoHDR.1 (Bol013657) | - | BoHDR.2 (Bol001484) | - |
| IPPI1 | AT5G16440 | - | BoIPPI1 (Bol034402) | - | - |
| IPPI2 | AT3G02780 | BoIPPI2.1 (Bol000950) | BoIPPI2.2 (Bol001738) | BoIPPI2.3 (Bol002232) | - |
| GGPS1 | AT4G36810 | BoGGPS1 (Bol028967) | - | - | - |
| GGPS2 | AT2G23800 | BoGGPS2 (Bol045796) | - | - | - |
| GGPS3 | AT3G14550 | BoGGPS3.1 (Bol005099) | - | BoGGPS3.2 (Bol025714) | - |
| GGPS7 | AT2G18620 | - | - | - | - |
| GGPS8 | AT3G14510 | - | - | - | - |
| GGPS11 | AT3G29430 | - | - | - | - |
| GGPS4 | AT2G18640 | - | - | - | - |
| GGPS6 | AT1G49530 | - | - | - | - |
| GGPS9 | AT3G14530 | - | - | - | - |
| GGPS10 | AT3G20160 | - | - | - | - |
| GGPS12 | AT3G32040 | - | - | - | - |
| GGR | AT4G38460 | BoGGR (Bol000792) | - | - | - |
| PSY | AT5G17230 | BoPSY.1 (Bol019820) | BoPSY.2 (Bol034439) | BoPSY.3 (Bol021326) | - |
| PDS3 | AT4G14210 | - | BoPDS3.1 (Bol009962) | BoPDS3.2 (Bol016089) | - |
| Z-ISO | AT1G10830 | BoZ-ISO (Bol036691) | - | - | - |
| ZDS | AT3G04870 | - | BoZDS (Bol003131) | - | - |
| CRTISO | AT1G06820 | - | - | BoCRTISO (Bol004146) | - |
| CRTISO2 | AT1G57770 | - | - | BoCRTISO2 (Bol016311) | - |
| LYC | AT3G10230 | BoLYC (Bol011368) | - | - | - |
| LUT2 | AT5G57030 | BoLUT2.1 (Bol037991) | BoLUT2.2 (Bol026053) | - | - |
| CHY1 | AT4G25700 | BoCHY1.1 (Bol039555) | BoCHY1.2 (Bol042254) | - | - |
| CHY2 | AT5G52570 | BoCHY2.1 (Bol030235) | - | BoCHY2.2 (Bol045243) | - |
| LUT5 | AT1G31800 | - | BoLUT5.1 (Bol027064) | - | BoLUT5.2 (Bol027080) BoLUT5.3 (Bol014196) |
| CYP97B3 | AT4G15110 | - | - | BoCYP97B3 (Bol028312) | - |
| LUT1 | AT3G53130 | - | - | - | - |
| ZEP | AT5G67030 | BoZEP.1 (Bol027222) | - | BoZEP.2 (Bol019241) | - |
| VDE | AT1G08550 | BoVDE (Bol041221) | - | - | - |
| NSY | AT1G67080 | - | BoNSY (Bol045096) | - | - |
| CCD7 | AT2G44990 | - | BoCCD7 (Bol021707) | - | - |
| CCD8 | AT4G32810 | BoCCD8.1 (Bol017864) | - | - | BoCCD8.2 (Bol000605) |
| NCED2 | AT4G18350 | BoNCED2.1 (Bol009433) | BoNCED2.2 (Bol037062) | - | - |
| NCED3 | AT3G14440 | BoNCED3.1 (Bol005093) | BoNCED3.2 (Bol011830) | BoNCED3.3 (Bol035582) | - |
| NCED4 | AT4G19170 | BoNCED4.1 (Bol009345) | - | BoNCED4.2 (Bol029878) | - |
| NCED5 | AT1G30100 | BoNCED5 (Bol022516) | - | - | - |
| NCED6 | AT3G24220 | BoNCED6 (Bol007451) | - | - | - |
| NCED9 | AT1G78390 | BoNCED9.1 (Bol027485) | BoNCED9.2 (Bol018961) | - | - |
| ABA2 | AT1G52340 | BoABA2.1 (Bol000478) | BoABA2.2 (Bol035060) | - | - |
| AAO3 | AT2G27150 | - | BoAAO3.1 (Bol026459) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, W.; Wang, P.; Yang, L.; Fang, Z.; Zhang, Y.; Zhuang, M.; Lv, H.; Wang, Y.; Ji, J. Carotenoid Biosynthetic Genes in Cabbage: Genome-Wide Identification, Evolution, and Expression Analysis. Genes 2021, 12, 2027. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122027
Cao W, Wang P, Yang L, Fang Z, Zhang Y, Zhuang M, Lv H, Wang Y, Ji J. Carotenoid Biosynthetic Genes in Cabbage: Genome-Wide Identification, Evolution, and Expression Analysis. Genes. 2021; 12(12):2027. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122027
Chicago/Turabian StyleCao, Wenxue, Peng Wang, Limei Yang, Zhiyuan Fang, Yangyong Zhang, Mu Zhuang, Honghao Lv, Yong Wang, and Jialei Ji. 2021. "Carotenoid Biosynthetic Genes in Cabbage: Genome-Wide Identification, Evolution, and Expression Analysis" Genes 12, no. 12: 2027. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12122027