
Identification of the CKM Gene as a Potential Muscle-Specific Safe Harbor Locus in Pig Genome

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Quantitative RT-PCR (qPCR)
2.3. Cell Culture and Transfection
2.4. T7EN1 Detection Assay and Sequencing
2.5. Screening of Monoclonal Cells
2.6. Off-Target Analysis of sgRNA
3. Results
3.1. Identification of the Expression of the CKM Gene in Various Tissues
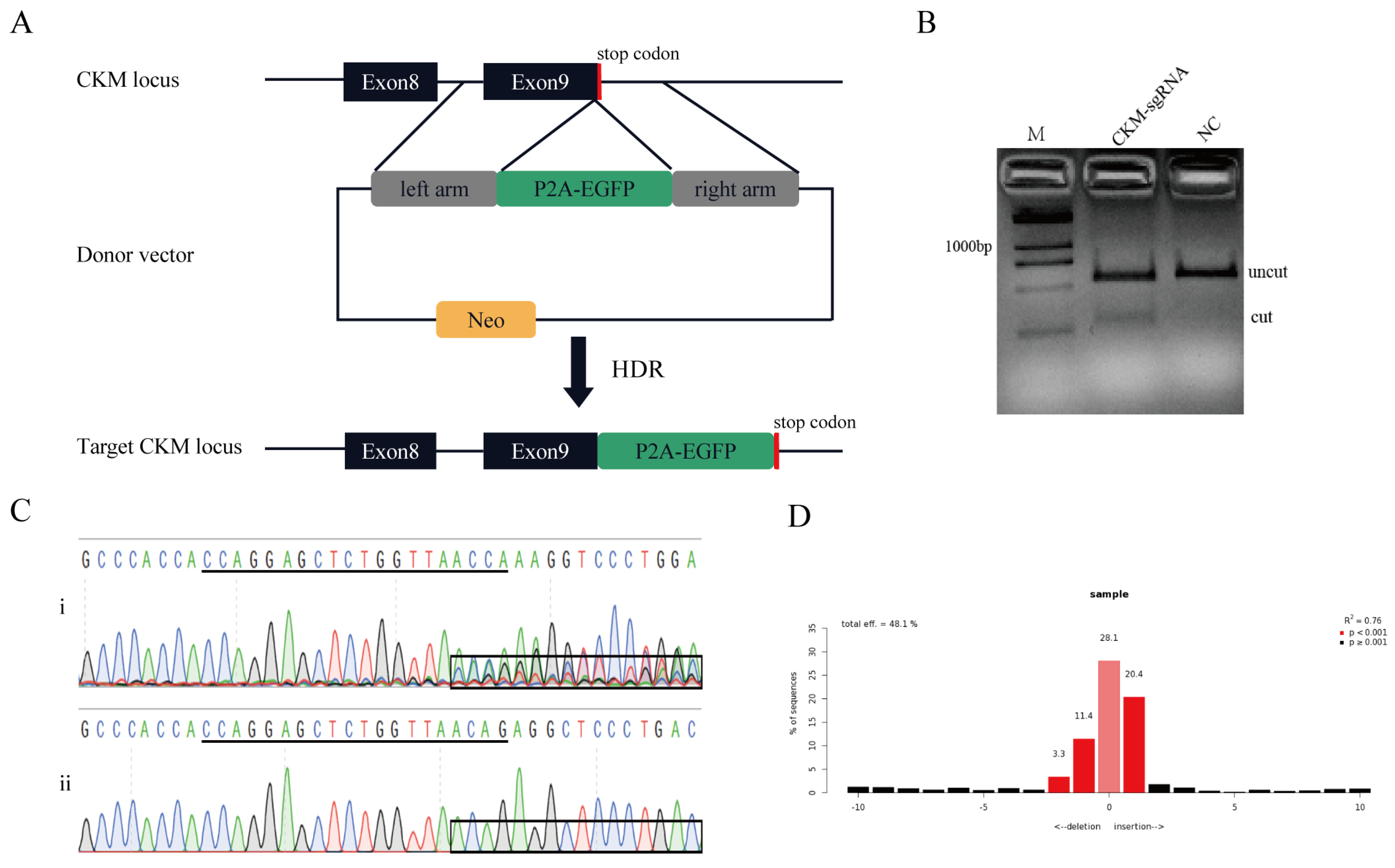
3.2. Establishment of the Knock-In Reporter System in Pig Genome
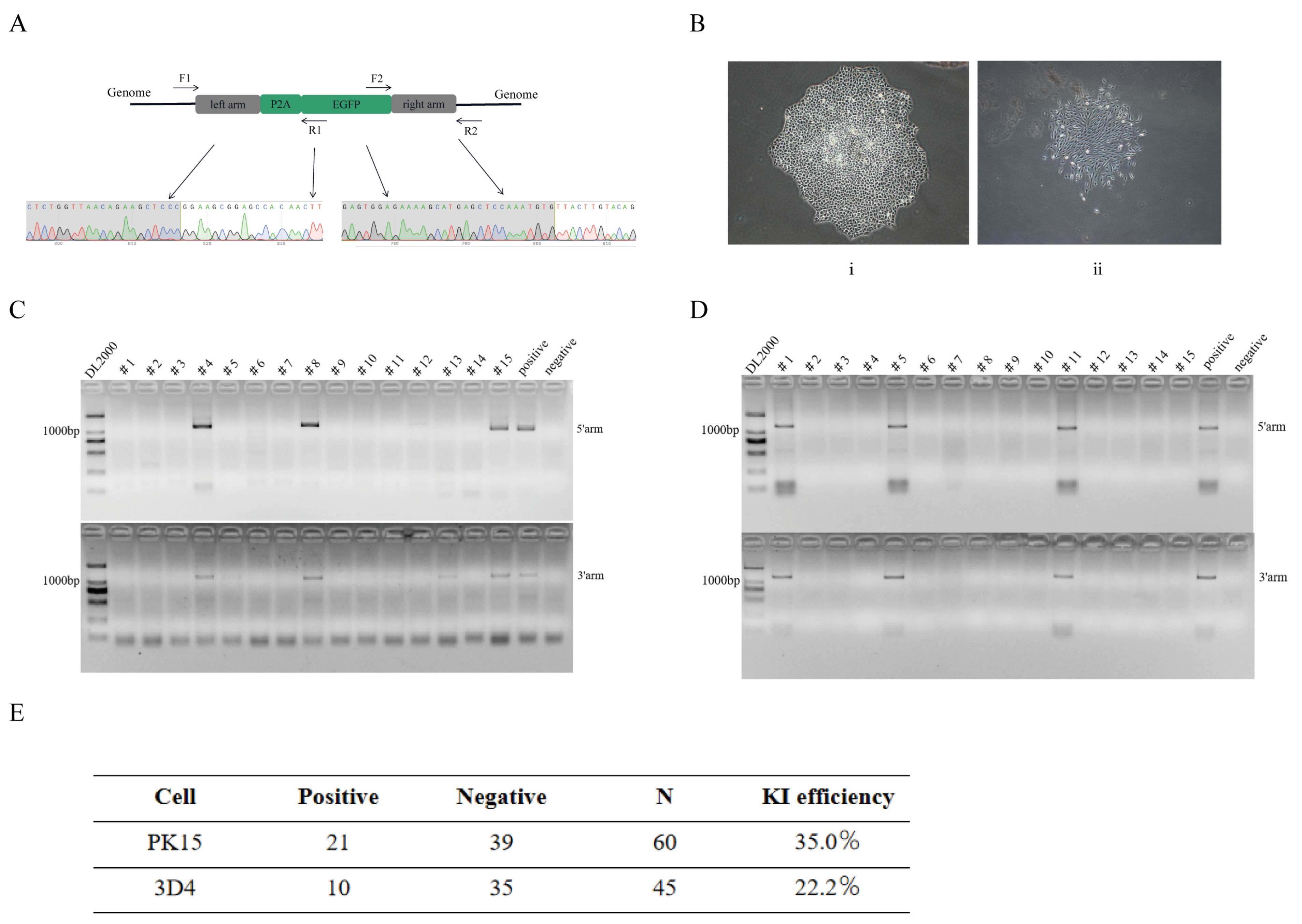
3.3. Assay of the Knock-In Efficiency in Pig Genome
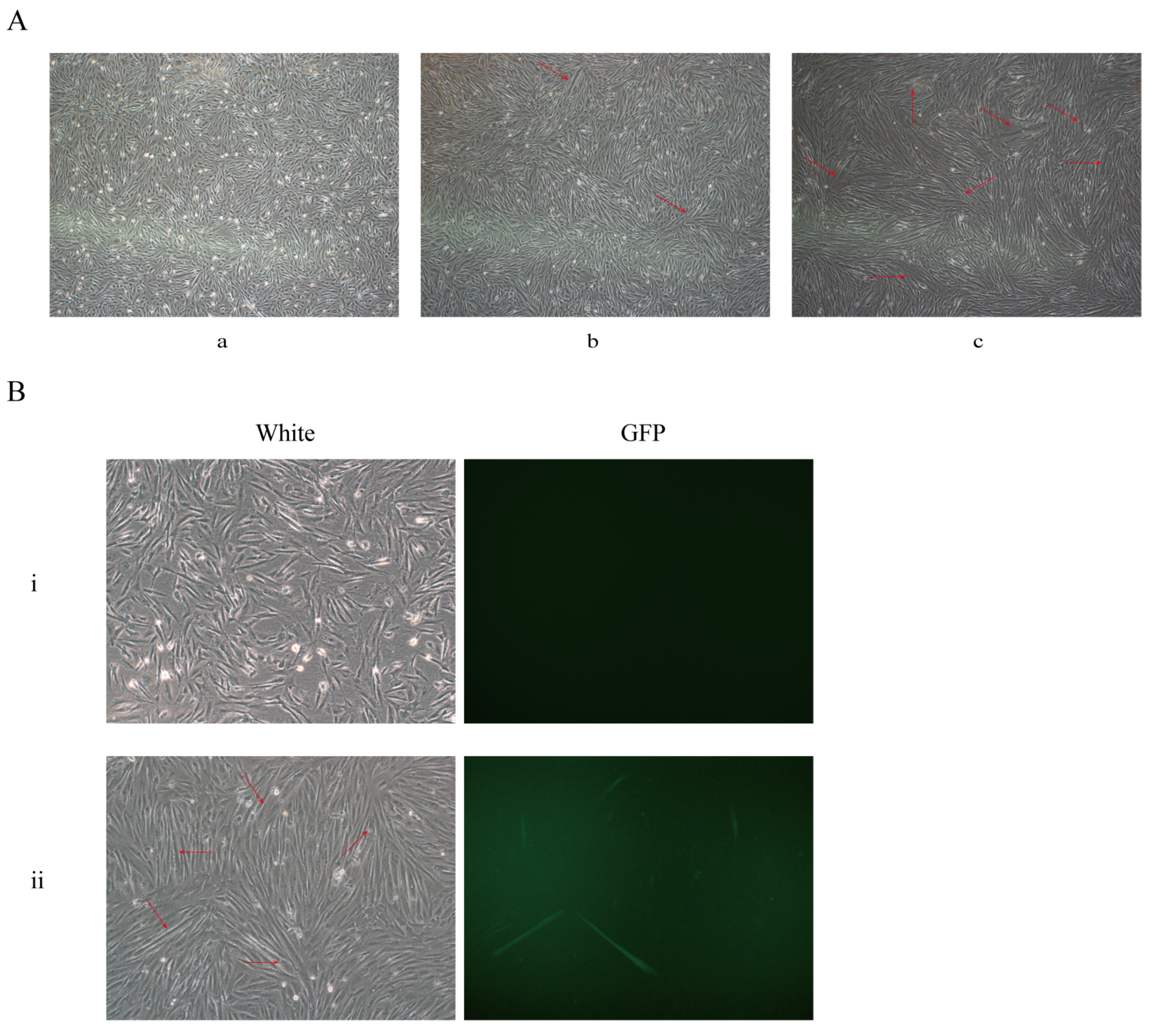
3.4. Identification of the CKM Gene Knock-In System in Muscle Satellite Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dai, Y.; Vaught, T.D.; Boone, J.; Chen, S.H.; Phelps, C.J.; Ball, S.; Monahan, J.A.; Jobst, P.M.; McCreath, K.J.; Lamborn, A.E.; et al. Targeted disruption of the alpha1,3-galactosyltransferase gene in cloned pigs. Nat. Biotechnol. 2002, 20, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Angelis, R.H.D.; Wurst, R.; Kühn, R. Gene targeting by homologous recombination in mouse zygotes mediated by zinc-finger nucleases. Proc. Natl. Acad. Sci. USA 2010, 107, 15022–15026. [Google Scholar] [CrossRef] [Green Version]
- Mahfouz, M.M.; Li, L.X.; Shamimuzzaman, M.; Wibowo, A.; Fang, X.Y.; Zhu, J.K. De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks. Proc. Natl. Acad. Sci. USA 2011, 108, 2623–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, B.; Frenzel, A.; Lucas-Hahn, A.; Herrmann, D.; Niemann, H. Efficient production of biallelic GGTA1 knockout pigs by cytoplasmic microinjection of CRISPR/Cas9 into zygotes. Xenotransplantation 2016, 23, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Scienece 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, H.I.; Ahmad, M.J.; Asif, A.R.; Adnan, M.; Iqbal, M.K.; Mehmood, K.; Muhammad, S.A.; Bhuiyan, A.A.; Elokil, A.; Du, X.; et al. A review of CRISPR-based genome editing: Survival, evolution and challenges. Curr. Issues Mol. Biol. 2018, 28, 47–68. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Yang, D.S.; Xu, J.; Zhu, T.Q.; Chen, Y.E.; Zhang, J.F. RS-1 enhances CRISPR/Cas9- and TALEN-mediated knock-in efficiency. Nat. Commun. 2016, 7, 381–396. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Becerra, G.; Kadonaga, J.T. Enhancement of homology-directed repair with chromatin donor templates in cells. eLife 2020, 9, e55780. [Google Scholar] [CrossRef]
- Yu, Y.; Guo, Y.; Tian, Q.; Lan, Y.; Yeh, H.; Zhang, M.; Tasan, I.; Jain, S.; Zhao, H. An efficient gene knock-in strategy using 5′-modified double-stranded DNA donors with short homology arms. Nat. Chem. Biol 2020, 16, 387–390. [Google Scholar] [CrossRef]
- Han, X.; Gao, Y.; Li, G.; Xiong, Y.; Zhao, C.; Ruan, J.; Ma, Y.; Li, X.; Li, C.; Zhao, S.; et al. Enhancing the antibacterial activities of sow milk via site-specifc knock-in of a lactoferrin gene in pigs using CRISPR/Cas9 technology. Cell Biosci. 2020, 10, 133. [Google Scholar] [CrossRef]
- Wen, L.; Zhao, C.; Song, J.; Ma, L.; Ruan, J.; Xia, X.; Chen, Y.E.; Zhang, J.; Ma, P.X.; Xu, J. CRISPR/Cas9-mediated TERT disruption in cancer cells. Int. J. Mol. Sci. 2020, 21, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Wang, C.E.; Zhao, B.; Li, W.; Ouyang, Z.; Liu, Z.; Yang, H.; Fan, P.; O’Neill, A.; Gu, W.; et al. Expression of Huntington’s disease protein results in apoptotic neurons in the brains of cloned transgenic pigs. Hum. Mol. Genet. 2010, 19, 3983–3994. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Tang, M.; Yang, J.; Wang, Q.; Cai, C.; Jiang, S.; Li, H.; Jiang, K.; Gao, P.; Ma, D.; et al. Targeted mutations in myostatin by zinc-finger nucleases result in double-muscled phenotype in Meishan pigs. Sci. Rep. 2015, 5, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Tu, Z.; Liu, Z.; Fan, N.; Yang, H.; Yang, S.; Yang, W.; Zhao, Y.; Ouyang, Z.; Lai, C.; et al. A Huntingtin knockin pig model recapitulates features of selective neurodegeneration in Huntington’s disease. Cell 2018, 4, 989–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, Y.; Bu, L.; Guo, X.; Tang, C.; Song, J.; Fan, N.; Zhao, B.; Ouyang, Z.; Liu, Z.; et al. Rosa26-targeted swine models for stable gene over-expression and Cre-mediated lineage tracing. Cell Res. 2014, 24, 501–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Li, H.; Xu, K.; Wu, T.; Wei, J.; Zhou, R.; Liu, Z.; Mu, Y.; Yang, S.; Ouyang, H.; et al. Highly efficient CRISPR/Cas9-mediated transgene knockin at the H11 locus in pigs. Sci. Rep. 2015, 5, 14253. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Wang, Y.; Wang, H.; Hu, Y.; Chen, J.; Tan, T.; Hu, M.; Liu, X.; Zhang, R.; Xing, Y.; et al. Screen and verification for transgene integration sites in pigs. Sci. Rep. 2018, 8, 7433. [Google Scholar] [CrossRef]
- Han, X.; Xiong, Y.; Zhao, C.; Xie, S.; Li, C.; Li, X.; Liu, X.; Li, K.; Zhao, S.; Ruan, J. Identification of glyceraldehyde-3-phosphate dehydrogenase gene as an alternative safe harbor locus in pig genome. Genes 2019, 10, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Han, X.; Zhang, J.; Zhao, G.; Wang, Z.; Zhuang, R.; Nie, X.; Xie, S.; Li, C.; Li, X.; et al. Identifcation of ACTB gene as a potential safe harbor locus in pig genome. Mol. Biotechnol. 2020, 62, 589–597. [Google Scholar] [CrossRef]
- Jaynes, J.B.; Johnson, J.E.; Buskin, J.N.; Gartside, C.L.; Hauschka, S.D. The muscle creatine kinase gene is regulated by multiple upstream elements, including a muscle-specific enhancer. Mol. Cell Biol. 1988, 8, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, E.K.; Chen, T.; Amendola, M.; Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 2014, 22, e168. [Google Scholar] [CrossRef]
- Trask, R.V.; Strauss, A.W.; Billadello, J.J. Developmental regulation and tissue-specific expression of the human muscle creatine kinase gene. J. Biol. Chem. 1988, 263, 17142–17149. [Google Scholar] [CrossRef]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Hai, T.; Teng, F.; Guo, R.F.; Li, W.; Zhou, Q. One-step generation of knockout pigs by zygote injection of CRISPR/Cas system. Cell Res. 2014, 24, 372–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saric, A.; Andreau, K.; Armand, A.S.; Mølle, I.M.; Petit, P.X. Barth syndrome: From mitochondrial dysfunctions associated with aberrant production of reactive oxygen species to pluripotent stem cell studies. Front. Genet. 2015, 6, 359. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.H.; Kim, Y.J.; Kim, E.Y.; Kim, S.E.; Kim, J.; Park, M.J.; Lee, H.G.; Park, S.P.; Kang, M.J. Knock-in fibroblasts and transgenic blastocysts for expression of human FGF2 in the bovine β-casein gene locus using CRISPR/Cas9 nuclease-mediated homologous recombination. Zygote 2016, 24, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J.; et al. Reconstitution of UCP1 using CRISPR/Cas9 in the white adipose tissue of pigs decreases fat deposition and improves thermogenic capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474–E9482. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Tsunekawa, Y.; Hernandez-Benitez, R.; Wu, J.; Zhu, J.; Kim, E.J.; Hatanaka, F.; Yamamoto, M.; Araoka, T.; Li, Z.; et al. In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration. Nature 2016, 540, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Wang, X.; Hu, X.; Liu, Z.; Liu, J.; Zhou, H.; Shen, X.; Wei, Y.; Huang, Z.; Ying, W.; et al. Homology-mediated end joining-based targeted integration using CRISPR/Cas9. Cell Res. 2017, 27, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wu, S.; Capecchi, M.R.; Jaenisch, R. A brief review of genome editing technology for generating animal models. Front. Agric. Sci. Eng. 2020, 7, 123–128. [Google Scholar] [CrossRef]
- Ma, S.; Wang, X.; Hu, Y.; Lv, J.; Liu, C.; Liao, K.; Guo, X.; Wang, D.; Lin, Y.; Rong, Z. Enhancing site-specific DNA integration by a Cas9 nuclease fused with a DNA donor-binding domain. Nucleic Acids Res. 2020, 48, 10590–10601. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Martins, M.; Cavalheiro, G.R.; Matos-Rodrigues, G.E.; Martins, R.A. From gene targeting to genome editing: Transgenic animals applications and beyond. Anais da Academia Brasileira de Ciências 2015, 87, 1323–1348. [Google Scholar] [CrossRef] [PubMed]
- Carey, K.; Ryu, J.; Uh, K.; Lengi, A.J.; Clark-Deener, S.; Corl, B.A.; Lee, K. Frequency of off-targeting in genome edited pigs produced via direct injection of the CRISPR/Cas9 system into developing embryos. BMC Biotechnol. 2019, 19, 25. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Predicted OTS | Sequence (5′ to 3′) | Indel |
|---|---|---|---|
| CKM-sgR1 | CCAGGAGCTCTGGTTAACAG AGG | ||
| 1 | Predicted-OFF-Target1 | CCTGGAGCTCCGGTTAGCAG GGG | NO |
| 2 | Predicted-OFF-Target2 | CGAGGAGGTCTGGCTAACAG GGG | NO |
| 3 | Predicted-OFF-Target3 | GCAGGAGCTCTGGATGACAG TGG | NO |
| 4 | Predicted-OFF-Target4 | GCAGGAGCTCTGTTTATCAG TGG | NO |
| 5 | Predicted-OFF-Target5 | CCTGGAGCTCTGGTTGGCAG TGG | NO |
| 6 | Predicted-OFF-Target6 | CCAGGAGCTCTGGGGCACAG AGG | NO |
| 7 | Predicted-OFF-Target7 | CCAGGAGCTCTGGGTGGCAG TGG | NO |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.; Zhuang, R.; Zhao, G.; Liu, Y.; Su, Y.; Wang, W.; Xi, X.; Yang, Y.; Han, X.; Xie, S.; et al. Identification of the CKM Gene as a Potential Muscle-Specific Safe Harbor Locus in Pig Genome. Genes 2022, 13, 921. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050921
Xiong Y, Zhuang R, Zhao G, Liu Y, Su Y, Wang W, Xi X, Yang Y, Han X, Xie S, et al. Identification of the CKM Gene as a Potential Muscle-Specific Safe Harbor Locus in Pig Genome. Genes. 2022; 13(5):921. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050921
Chicago/Turabian StyleXiong, Youcai, Rongzhi Zhuang, Guangxing Zhao, Yanwen Liu, Yinyu Su, Wei Wang, Xiaoning Xi, Yanyu Yang, Xiaosong Han, Shengsong Xie, and et al. 2022. "Identification of the CKM Gene as a Potential Muscle-Specific Safe Harbor Locus in Pig Genome" Genes 13, no. 5: 921. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050921