
Gut Microbiota as a Potential Predictive Biomarker in Relapsing-Remitting Multiple Sclerosis

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Method
2.1. Study Population
2.2. High-Throughput Sequencing of Stool Samples of Patients with MS
2.3. Bioinformatics
2.4. Statistical Analysis
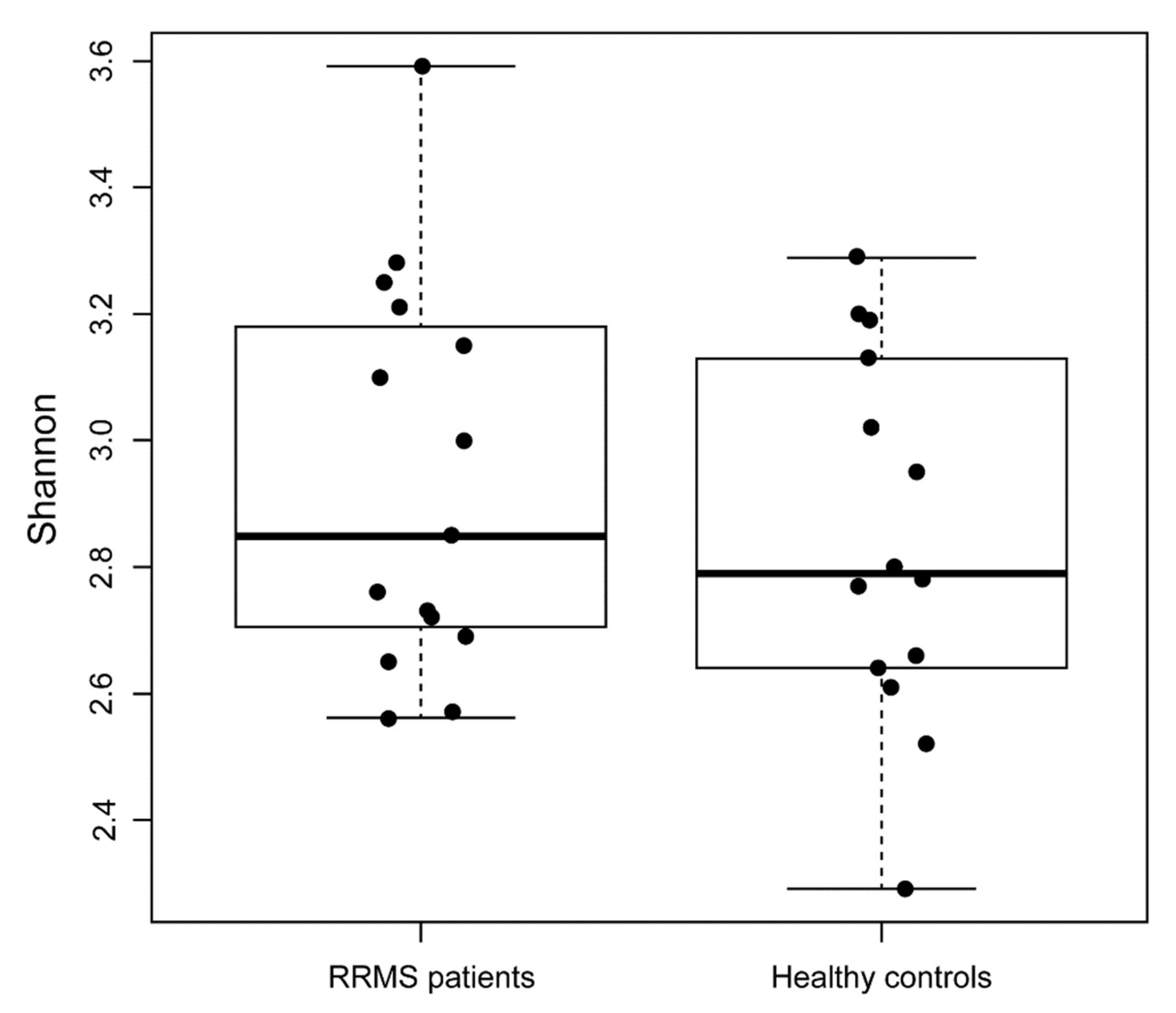
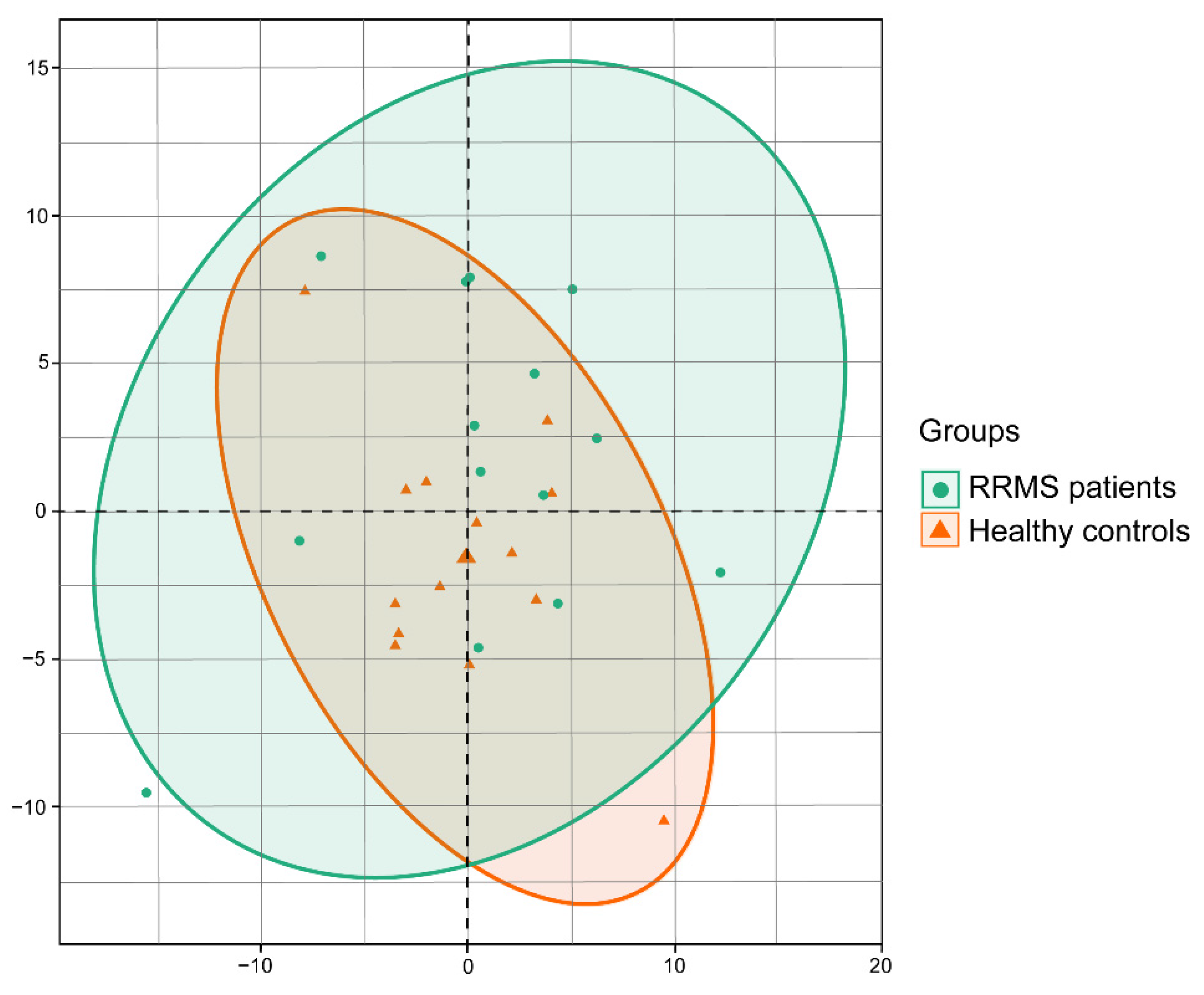
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calvo-Barreiro, L.; Eixarch, H.; Montalban, X.; Espejo, C. Combined therapies to treat complex diseases: The role of the gut microbiota in multiple sclerosis. Autoimmun. Rev. 2018, 17, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Sawcer, S.; Franklin, R.J.M.; Ban, M. Multiple sclerosis genetics. Lancet Neurol. 2014, 13, 700–709. [Google Scholar] [CrossRef]
- Simpson, S.; Blizzard, L.; Otahal, P.; Van der Mei, I.; Taylor, B. Latitude is significantly associated with the prevalence of multiple sclerosis: A meta-analysis. J. Neurol. Neurosurg. Psychiatry 2011, 82, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Fleming, J.; Fabry, Z. The hygiene hypothesis and multiple sclerosis. Ann. Neurol. 2007, 61, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Mendibe, M.M.; Boyero, S.; Rodrigo, R.M.; Zarranz, J.J. Esclerosis múltiple y encefalitis autoinmunes. Leucodistrofias y otras enfermedades desmielinizantes. In Neurología, 5th ed.; Zarranz, J.J., Ed.; Elsevier: Madrid, Spain, 2013; pp. 451–475. [Google Scholar]
- Weiner, H.L. The challenge of multiple sclerosis: How do we cure a chronic heterogeneous disease? Ann. Neurol. 2009, 65, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Krumbholz, M.; Derfuss, T.; Hohlfeld, R. B cells and antibodies in multiple sclerosis pathogenesis and therapy. Nat. Rev. Neurol. 2012, 8, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Mindur, J.E.; Ito, K. Advances in the immunopathogenesis of multiple sclerosis. Curr. Opin. Neurol. 2015, 28, 206–219. [Google Scholar] [CrossRef]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Paz, M.M.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar] [CrossRef] [Green Version]
- Hanne, F.H.; Gold, R.; Tintoré, M. Sex and gender issues in multiple sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar]
- Hauser, S.L.; Cree, B.A.C. Treatment of multiple sclerosis: A review. Am. J. Med. 2020, 133, 1380–1390. [Google Scholar] [CrossRef]
- Lünemann, J.D.; Ruck, T.; Muraro, P.A.; Bar-Or, A.; Wiendl, H. Immune reconstitution therapies: Concepts for durable remission in multiple sclerosis. Nat. Rev. Neurol. 2019, 16, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Río, J.; Nos, C.; Tintoré, M.; Borrás, C.; Galán, I.; Comabella, M.; Montalban, X. Assessment of different treatment failure criteria in a cohort of relapsing-remitting multiple sclerosis patients treated with interferon beta: Implications for clinical trials. Ann. Neurol. 2002, 52, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Ziemssen, T.; Akgün, K.; Brück, W. Molecular biomarkers in multiple sclerosis. J. Neuroinflamm. 2019, 16, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, M.; Higgins, P.D.; Middha, S.; Rioux, K.P. The human gut microbiome: Current knowledge, challenges, and future directions. Transl. Res. 2012, 160, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Torrazza, R.M.; Neu, J. The developing intestinal microbiome and its relationship to health and disease in the neonate. J. Perinatol. 2011, 31, S29–S34. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.J.; Musfeldt, M.; Wenderoth, D.F.; Hampe, J.; Brant, O.; Fölsch, U.R.; Timmis, K.N.; Schreiber, S. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut 2004, 53, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Stecher, B.; Chaffron, S.; Käppeli, R.; Hapfelmeier, S.; Freedrich, S.; Weber, T.C.; Kirundi, J.; Suar, M.; McCoy, K.D.; von Mering, C.; et al. Like will to like: Abundances of closely related species can predict susceptibility to intestinal colonization by pathogenic and commensal bacteria. PLoS Pathog. 2010, 6, e1000711. [Google Scholar] [CrossRef]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat. Med. 2010, 16, 228–231. [Google Scholar] [CrossRef] [Green Version]
- Berer, K.; Krishnamoorthy, G. Commensal gut flora and brain autoimmunity: A love or hate affair? Acta Neuropathol. 2012, 123, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Gelfand, J.M.; Lewis, J.D. Increased risk for demyelinating diseases in patients with inflammatory bowel disease. Gastroenterology 2005, 129, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Reparaz, J.; Mielcarz, D.W.; Ditrio, L.E.; Burroughs, A.R.; Begum-Haque, S.; Dasgupta, S.; Kasper, D.L.; Kasper, L.H. Central Nervous System Demyelinating Disease Protection by the Human Commensal Bacteroides fragilis Depends on Polysaccharide A Expression. J. Immunol. 2010, 185, 4101–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokote, H.; Miyake, S.; Croxford, J.L.; Oki, S.; Mizusawa, H.; Yamamura, T. NKT cell-dependent amelioration of a mouse model of multiple sclerosis by altering gut flora. Am. J. Pathol. 2008, 173, 1714–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 158, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and treatment of multiple sclerosis: A review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef]
- Yuan, S.; Cohen, D.B.; Ravel, J.; Abdo, Z.; Forney, L.J. Evaluation of methods for the extraction and purification of DNA from the human microbiome. PLoS ONE 2012, 7, e33865. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Horton, M.K.; McCauley, K.; Fadrosh, D.; Fujimura, K.; Graves, J.; Ness, J.; Wheeler, Y.; Gorman, M.P.; Benson, L.A.; Weinstock-Guttman, B.; et al. Gut microbiome is associated with multiple sclerosis activity in children. Ann. Clin. Transl. Neurol. 2021, 8, 1867–1883. [Google Scholar] [CrossRef]
- Tremlett, H.; Fadrosh, D.W.; Faruqi, A.A.; Hart, J.; Roalstad, S.; Graves, J.; Lynch, S.; Waubant, E.; Aaen, G.; Belman, A.; et al. Gut microbiota composition and relapse risk in pediatric MS: A pilot study. J. Neurol. Sci. 2016, 363, 153–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahi, S.K.; Freedman, S.N.; Mangalam, A.K. Gut microbiome in multiple sclerosis: The players involved and the roles they play. Gut Microbes 2017, 8, 607–615. [Google Scholar] [CrossRef]
- Forbes, J.D.; Van Domselaar, G.; Bernstein, C.N. The gut microbiota in immune-mediated inflammatory diseases. Front. Microbiol. 2016, 7, 1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tankou, S.K.; Regev, K.; Healy, B.C.; Tjon, E.; Laghi, L.; Cox, L.M.; Kivisäkk, L.; Pierre, I.V.; Hrishikesh, L.; Gandhi, R.; et al. A probiotic modulates the microbiome and immunity in multiple sclerosis. Ann. Neurol. 2018, 83, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Waubant, E.; Chehoud, C.; Kuczynski, J.; DeSantis, T.Z.; Warrington, J.; Venkatesan, A.; Fraser, C.M.; Mowry, E.M. Gut microbiota in multiple sclerosis: Possible influence of immunomodulators. J. Investig. Med. 2015, 63, 729–734. [Google Scholar] [CrossRef]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar] [CrossRef] [Green Version]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; De Vos, M.; Boon, B.; Van de Wiele, T. Butyrate-producing bacteria supplemented in vitro to Crohn’s disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci. Rep. 2017, 7, 11450. [Google Scholar] [CrossRef] [Green Version]
- Tremlett, H.; Fadrosh, D.W.; Faruqi, A.A.; Zhu, F.; Hart, J.; Roalstad, S.; Graves, J.; Lynch, S.; Waubant, E.; US Network of Pediatric MS Centers. Gut microbiota in early pediatric multiple sclerosis: A case-control study. Eur. J. Neurol. 2016, 23, 1308–1321. [Google Scholar] [CrossRef] [Green Version]
- Issa, M.; Ananthakrishnan, A.N.; Binion, D.G. Clostridium difficile and inflammatory bowel disease. Inflamm. Bowel. Dis. 2008, 14, 1432–1442. [Google Scholar] [CrossRef]
- Sansonetti, P.J. Rupture, Invasion and Inflammatory Destruction of the Intestinal Barrier by Shigella: The Yin and Yang of Innate Immunity. Can. J. Infect. Dis. Med. Microbiol. 2006, 17, 117–119. [Google Scholar] [CrossRef] [Green Version]
- Planas, R.; Santos, R.; Tomas-Ojer, P.; Cruciani, C.; Lutterotti, A.; Faigle, W.; Schaeren-Wiemers, N.; Espejo, C.; Eixarch, H.; Pinilla, C.; et al. GDP-l-fucose synthase is a CD4T+ T cell-specific autoantigen in DRB3*02:02 patients with multiple sclerosis. Sci. Transl. Med. 2018, 10, eaat4301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangalam, A.K.; Yadav, M.; Yadav, R. The emerging world of microbiome in autoimmune disorders: Opportunities and challenges. Indian J. Rheumatol. 2021, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, J.J.; Zamvil, S.S. T cells take aim at a ubiquitous autoantigen in multiple sclerosis. Sci. Transl. Med. 2018, 10, eaau8826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Hoogen, W.J.; Laman, J.D.; Hart, B.A. Modulation of Multiple Sclerosis and Its Animal Model Experimental Autoimmune Encephalomyelitis by Food and Gut Microbiota. Front. Immunol. 2017, 8, 1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas-Sierra, A.; Romo-Hualde, A.; Aranaz, P.; Goni, L.; Cuervo, M.; Martínez, J.A.; Milagro, F.I.; Riezu-Boj, J.I. Diet-and sex-related changes of gut microbiota composition and functional profiles after 4 months of weight loss intervention. Eur. J. Nutr. 2021, 60, 3279–3301. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
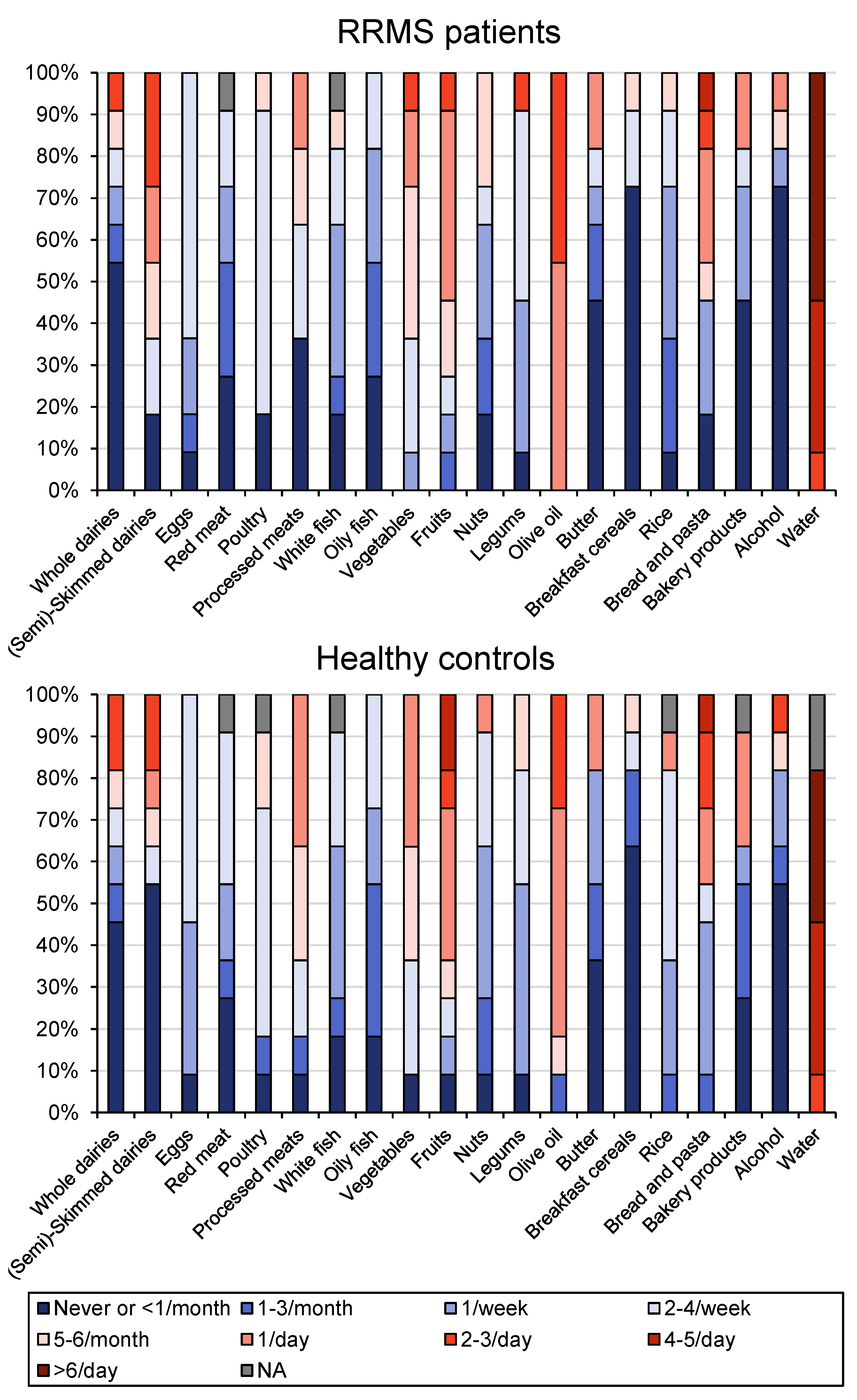{kind=link}
| Patient | Gender | Age (Years) | Weight (kg) | Height (m) | BMI (kg/m2) | DMT | Initial Clinical Presentation | EDSS | Relapses | MRI Lesions |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Female | 38 | 90 | 1.72 | 30.42 | beta-interferon | Brainstem | 2.5 | 0 | 0 |
| 2 | Female | 18 | 57 | 1.71 | 19.49 | beta-interferon | Myelitis | 1 | 1 | 1 |
| 3 | Female | 38 | 57 | 1.54 | 22.77 | natalizumab | Myelitis | 2.5 | 0 | 0 |
| 4 | Female | 35 | 59 | 1.51 | 25.88 | fingolimod | Myelitis | 1 | 0 | 0 |
| 5 | Female | 35 | 107 | 1.78 | 33.77 | dimethyl fumarate | Myelitis, brainstem | 3.5 | 1 | 0 |
| 6 | Female | 34 | 75,5 | 1.60 | 29.49 | beta-interferon | Optic neuritis | 3 | 3 | 6 |
| 7 | Female | 31 | 100 | 1.80 | 30.86 | fingolimod | Hemiparesis | 3.5 | 2 | 5 |
| 8 | Male | 33 | 85.5 | 1.70 | 29.58 | natalizumab | Myelitis | 6.5 | 0 | 0 |
| 9 | Female | 45 | 59 | 1.65 | 21.67 | fingolimod | Myelitis | 2.5 | 2 | 8 |
| 10 | Female | 53 | 92.7 | 1.55 | 38.58 | beta-interferon | Brainstem | 3.5 | 0 | 2 |
| 11 | Female | 37 | 63 | 1.69 | 22.05 | beta-interferon | Brainstem | 1.5 | 0 | 0 |
| 12 | Male | 44 | 92 | 1.73 | 30.77 | teriflunomide | Hemiparesis | 3. 5 | 2 | 3 |
| 13 | Female | 29 | 54 | 1.60 | 21.09 | dimethyl fumarate | Hemiparesis | 1 | 0 | 3 |
| 14 | Female | 51 | 63 | 1.66 | 22.86 | beta-interferon | Brainstem, optic neuritis | 2.5 | 1 | 4 |
| 15 | Female | 37 | 59 | 1.51 | 25.88 | glatiramer acetate | Brainstem | 2 | 0 | 0 |
| Genera | Multiplier 95% CI | p-Value | AUC (95% CI) |
|---|---|---|---|
| Lachnospiraceae | 0.06 (0.01 to 0.35) | 0.001 | 67.1 (46.7 to 87.6) |
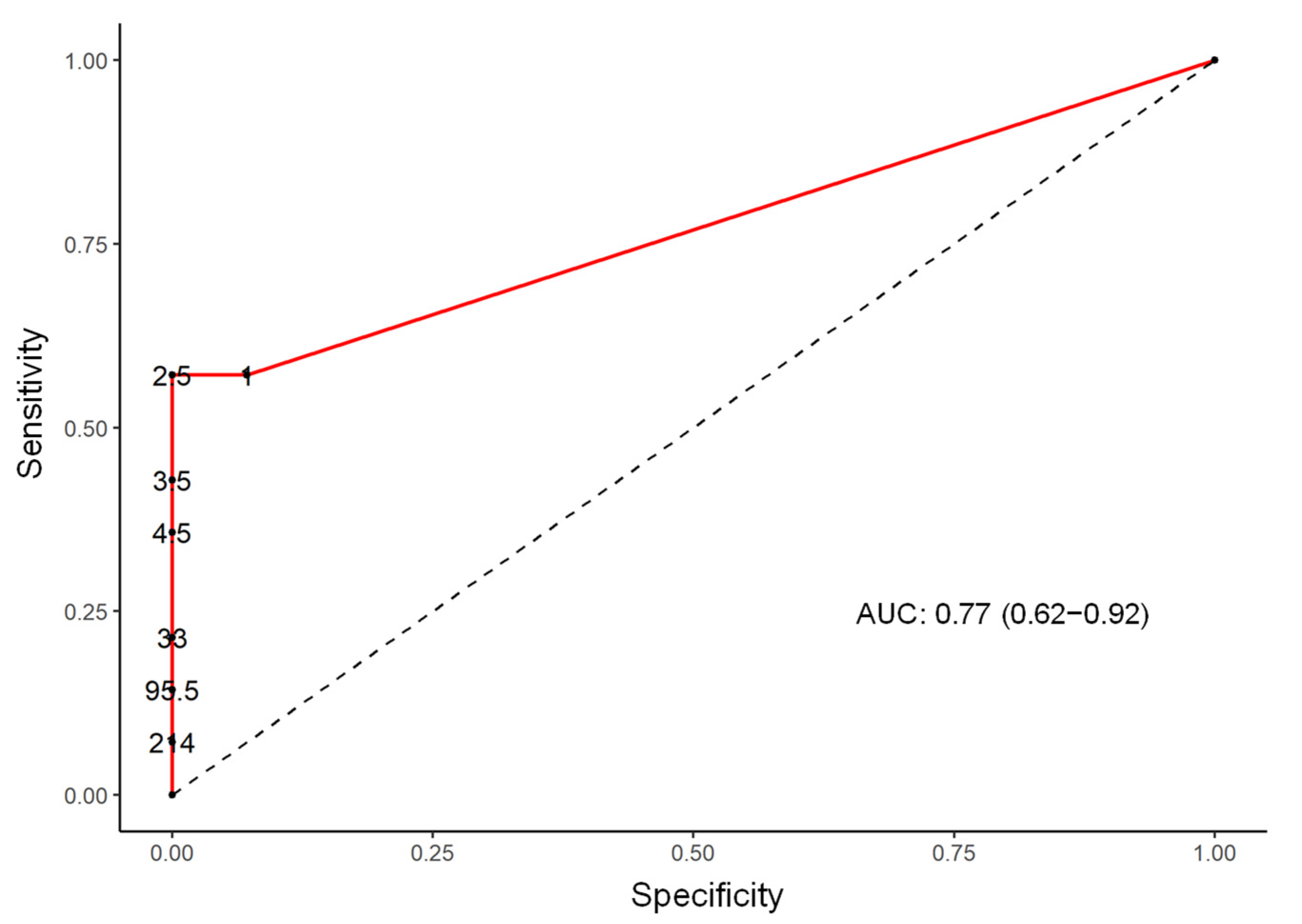
| Ezakiella | 13.08 (2.16 to 79.24) | 0.005 | 77.0 (62.0 to 92.0) |
| Ruminococcaceae | 12.65 (2.09 to 76.62) | 0.006 | 68.1 (47.9 to 88.3) |
| Hungatella | 8.57 (1.41 to 51.89) | 0.02 | 68.3 (51.3 to 85.4) |
| Roseburia | 0.12 (0.02 to 0.73) | 0.02 | 68.1 (50.2 to 86.0) |
| Clostridium | 7.26 (1.20 to 43.99) | 0.03 | 67.4 (47.5 to 87.2) |
| Shuttleworthia | 6.32 (1.04 to 38.29) | 0.04 | 62.4 (42.5 to 82.3) |
| Ruminococcaceae | 6.23 (1.03 to 37.71) | 0.04 | 62.4 (42.1 to 82.7) |
| Porphyromonas | 6.00 (0.99 to 36.33) | 0.05 | 64.3 (46.0 to 82.5) |
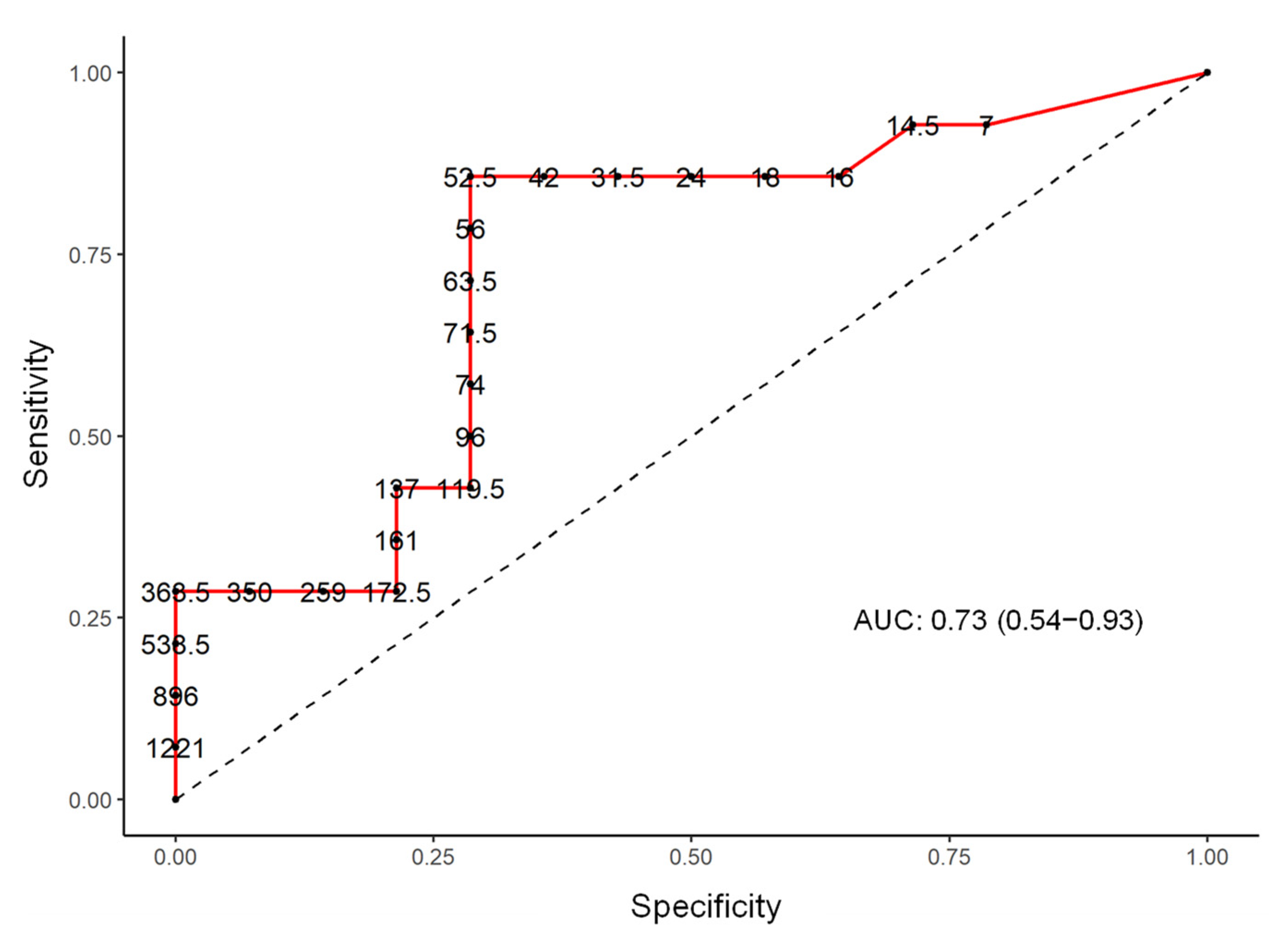
| Bilophila | 5.93 (0.98 to 35.91) | 0.05 | 73.0 (54.0 to 93.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-López, V.; Méndez-Miralles, M.Á.; Vela-Yebra, R.; Fríes-Ramos, A.; Sánchez-Pellicer, P.; Ruzafa-Costas, B.; Núñez-Delegido, E.; Gómez-Gómez, H.; Chumillas-Lidón, S.; Picó-Monllor, J.A.; et al. Gut Microbiota as a Potential Predictive Biomarker in Relapsing-Remitting Multiple Sclerosis. Genes 2022, 13, 930. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050930
Navarro-López V, Méndez-Miralles MÁ, Vela-Yebra R, Fríes-Ramos A, Sánchez-Pellicer P, Ruzafa-Costas B, Núñez-Delegido E, Gómez-Gómez H, Chumillas-Lidón S, Picó-Monllor JA, et al. Gut Microbiota as a Potential Predictive Biomarker in Relapsing-Remitting Multiple Sclerosis. Genes. 2022; 13(5):930. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050930
Chicago/Turabian StyleNavarro-López, Vicente, María Ángeles Méndez-Miralles, Rosa Vela-Yebra, Ana Fríes-Ramos, Pedro Sánchez-Pellicer, Beatriz Ruzafa-Costas, Eva Núñez-Delegido, Humberto Gómez-Gómez, Sara Chumillas-Lidón, Jose A. Picó-Monllor, and et al. 2022. "Gut Microbiota as a Potential Predictive Biomarker in Relapsing-Remitting Multiple Sclerosis" Genes 13, no. 5: 930. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13050930