siRNA Treatment: “A Sword-in-the-Stone” for Acute Brain Injuries

Abstract
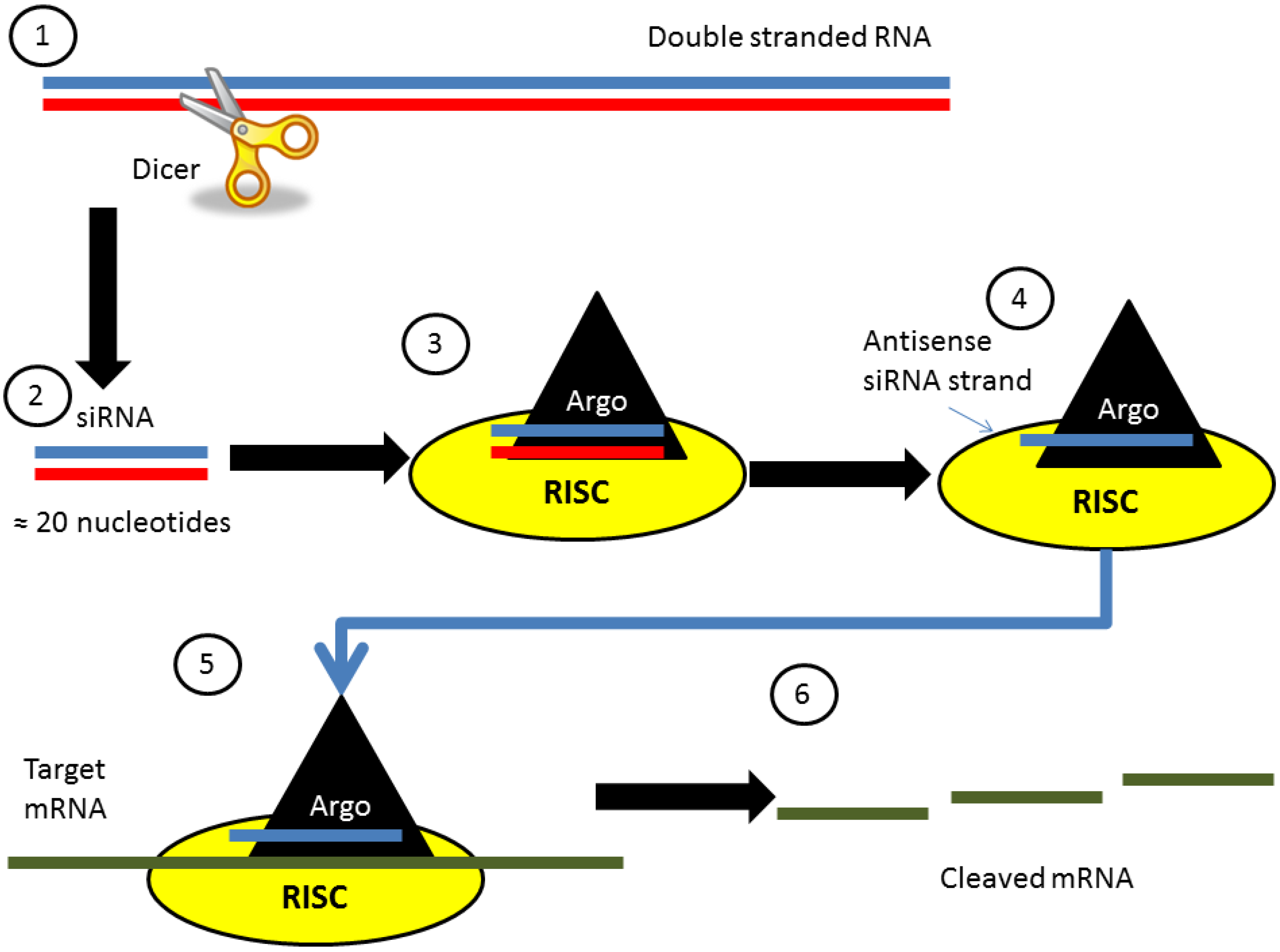
:1. What is siRNA?
1.1. Discovery and Endogenous Function

1.2. Clinical Treatment Tool
2. siRNA and Acute Brain Injuries
2.1. Non-Traumatic Brain Hemorrhage Models
2.1.1. Background

{kind=link}
| Reference | Targeted Protein | Pathophysiological Pathway Targeted | Injury Model | Delivery Method | Delivery Timepoint | Result |
|---|---|---|---|---|---|---|
| [12] | CHOP | apoptosis | Endovascular perforation of MCA | ICV | 24 h prior | ↓ edema ↓ BBB disruption ↓ behavior deficit |
| [13] | CHOP | apoptosis | Endovascular perforation of MCA | ICV | 24 h prior | ↓ cell death Reversed detrimental phenotypic changes of the blood vessels |
| [14] | PUMA | apoptosis | Endovascular perforation | ICV | Immediately after | ↓ cell death ↓ edema ↓ behavior deficit ↓ BBB disruption ↓ mortality rate |
| [15] | VAP-1 | neuroinflammation | Collagenase Injection | ICV | 48 h prior | ↓ edema ↑ behavior ↓ microglial activation ↓ proinflammatory molecule secretion |
2.1.2. In Vivo siRNA Studies
2.2. Cerebral Ischemic Stroke
2.2.1. Background
2.2.2. In Vivo Studies
| Reference | Targeted Protein | Pathophysiological Pathway Targeted | Injury Model | Delivery Method | Delivery Timepoint | Result |
|---|---|---|---|---|---|---|
| [27] | Caspase-3 | Apoptosis | ET-1 Injection Rats | Intracerebral cortex Injection | 24 h Prior | ↓ TUNEL ↑ behavior outcome (significant only at 24 h PreInjury) |
| 24 h Post | ||||||
| [26] | Beclin1 | Apoptosis | Transient MCAO Rats | Lateral Ventricle Injection | 7 d Prior | ↓ infarct volume ↑ neurological outcome |
| [28] | Ask1 | Apoptosis | Transient MCAO Mice | Osmotic Minipump Ventricle | Continuously 3 d Prior | ↓ infarct volume ↓ TUNEL |
| [29] | PAR1 | Coagulation Cascade | Transient MCAO Rats | Lateral Ventricle Injection | 7 d Prior | ↓ infarct volume ↑ neurological outcome |
| [32] | HIF1α | Hypoxia Induced Cascade | Transient MCAO Rats | Intraparenchyal Injection | <1 h Post | ↓ mortality ↑ neurological outcome ↓ Infarct Volume ↓ BBB disruption ↓ cell death |
| [33] | GPR17 | Microgliosis (neuroinflammation) | Transient MCAO Rats | ICV Injection | Once daily from 2 d prior to 7 d post and then every other day from day 8–14 | 24 h post injury ↓ neurological dysfunction ↓ infarction ↓ neuron loss |
| 14 d Post injury ↓ brain atrophy ↓ neuron loss ↓ microglial activation | ||||||
| [41] | HMGB1 | Neuroinflammation | Transient MCAO | Intranasal | 1 h Prior | ↓ infarct volume ↑ behavior |
2.3. Traumatic Brain Injury
2.3.1. Background
2.3.2. In Vivo Studies
| Reference | Targeted Protein | Pathophysiological Pathway Targeted | Injury Model | Delivery Method | Delivery Timepoint | Result |
|---|---|---|---|---|---|---|
| [51] | Claudin-5 | Edema | Cold Induced Mice | Tail Vein Injection | <1 h Post | ↓ lesion size ↑ cognitive outcome |
| [49] | Int6 | Angiogenesis | Cold Induced Rats | Internal carotid Artery | 1 h Post | ↓ lesion |
| [55] | Fzd2 | Ca2+ Accumulation | Weight Drop Rats | Direct Hippocampal Injection | 48 h Prior | ↓ Fzd2, Wnt5a, p-CaMKII ↓ intracellular Ca2+ |
| [54] | AQP4 | Edema Formation | Controlled Cortical Injury | Intracortical Injection | Immediately after injury and 2 d after injury (2 injections) | Acutely, ↑ motor function ↑ neuronal survival ↓ BBB disruption ↓ edema Chronically, ↑ memory ↑ neuronal survival |
2.4. Overall Summary of the In Vivo Acute Brain Injury Studies
| Method of delivery | Advantages | Disadvantages |
|---|---|---|
| Intra-cerebroventricular | Directly in the brain; Distribute in all brain | Difficult to apply in clinic; Dilution of the siRNA and potential degradation; Off target effects on other brain regions |
| Intra-vascular, tail vein and carotid | Possible in clinic | Difficulties to cross BBB; Off target effects on other organs (i.e., Liver); Dilution of the siRNA; Increase chance of siRNA degradation |
| In the brain structures: Cortex and hippocampus | In the targeted brain region; Small amount of siRNA | Difficult to apply in clinic; Mechanical lesion by the injection; Neuroinflammation |
| Intra-nasal | Target the brain; Diffusion along the perivascular space; Possible in clinic; Accommodate large molecules | Potential dilution of siRNA and degradation; Off target effects on lungs and other brain regions |
3. Future Direction
3.1. Intranasal Drug Delivery
3.2. Exosomes, Microvesicles and Nanoparticles
4. Conclusions
Abbreviations
| small interfering RNA | (siRNA) |
| blood-brain barrier | (BBB) |
| traumatic brain injury | (TBI) |
| subarachnoid hemorrhage | (SAH) |
| intracerebral hemorrhage | (ICH) |
| middle cerebral artery | (MCA) |
| middle cerebral artery occlusion | (MCAo) |
| intracereboventricular | (ICV) |
| neurovascular unit | (NVU) |
Acknowledgements
Conflicts of Interest
References
- Burnett, J.C.; Rossi, J.J.; Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnol. J. 2011, 6, 1130–1146. [Google Scholar]
- Burnett, J.C.; Rossi, J.J. RNA-based therapeutics: Current progress and future prospects. Chem. Biol. 2012, 19, 60–71. [Google Scholar] [CrossRef]
- Brower, V. RNA interference advances to early-stage clinical trials. J. Nat. Cancer Inst. 2010, 102, 1459–1461. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef]
- Ketting, R.F.; Fischer, S.E.; Bernstein, E.; Sijen, T.; Hannon, G.J.; Plasterk, R.H. Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Gene. Dev. 2001, 15, 2654–2659. [Google Scholar] [CrossRef]
- Montgomery, M.K.; Xu, S.; Fire, A. RNA as a target of double-stranded RNA-mediated genetic interference in Caenorhabditis elegans. Proc. Nat. Acad. Sci. USA 1998, 95, 15502–15507. [Google Scholar] [CrossRef]
- Tabernero, J.; Shapiro, G.I.; Lorusso, P.M.; Cervantes, A.; Schwartz, G.K.; Weiss, G.J.; Paz-Ares, L.; Cho, D.C.; Infante, J.R.; Alsina, M.; et al. First-in-humans trial of an RNA interference therapeutic targeting VEGF and KSP in cancer patients with liver involvement. Cancer Discov. 2013, 3, 406–417. [Google Scholar] [CrossRef]
- Thanvi, B.R.; Sprigg, N.; Munshi, S.K. Advances in spontaneous intracerebral haemorrhage. Int. J. Clin. Pract. 2012, 66, 556–564. [Google Scholar] [CrossRef]
- Caner, B.; Hou, J.; Altay, O.; Fuj, M., 2nd.; Zhang, J.H. Transition of research focus from vasospasm to early brain injury after subarachnoid hemorrhage. J. Neurochem. 2012, 123, 12–21. [Google Scholar] [CrossRef]
- He, Z.; Ostrowski, R.P.; Sun, X.; Ma, Q.; Huang, B.; Zhan, Y.; Zhang, J.H. CHOP silencing reduces acute brain injury in the rat model of subarachnoid hemorrhage. Stroke 2012, 43, 484–490. [Google Scholar] [CrossRef]
- He, Z.; Ostrowski, R.P.; Sun, X.; Ma, Q.; Tang, J.; Zhang, J.H. Targeting C/EBP homologous protein with siRNA attenuates cerebral vasospasm after experimental subarachnoid hemorrhage. Exp. Neurol. 2012, 238, 218–224. [Google Scholar] [CrossRef]
- Yan, J.; Li, L.; Khatibi, N.H.; Yang, L.; Wang, K.; Zhang, W.; Martin, R.D.; Han, J.; Zhang, J.; Zhou, C. Blood-brain barrier disruption following subarchnoid hemorrhage may be faciliated through PUMA induction of endothelial cell apoptosis from the endoplasmic reticulum. Exp. Neurol. 2011, 230, 240–247. [Google Scholar] [CrossRef]
- Ma, Q.; Manaenko, A.; Khatibi, N.H.; Chen, W.; Zhang, J.H.; Tang, J. Vascular adhesion protein-1 inhibition provides antiinflammatory protection after an intracerebral hemorrhagic stroke in mice. J. Cerebr. Blood Flow Metabol. 2011, 31, 881–893. [Google Scholar] [CrossRef]
- Zhang, J.H.; Badaut, J.; Tang, J.; Obenaus, A.; Hartman, R.; Pearce, W.J. The vascular neural network—A new paradigm in stroke pathophysiology. Nat. Rev. Neurol. 2012, 8, 711–716. [Google Scholar] [CrossRef]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Adams, R.J.; Berry, J.D.; Brown, T.M.; Carnethon, M.R.; Dai, S.; de Simone, G.; Ford, E.S.; et al. Heart disease and stroke statistics—2011 update: A report from the American Heart Association. Circulation 2011, 123, e18–e209. [Google Scholar] [CrossRef]
- Rosamond, W.; Flegal, K.; Friday, G.; Furie, K.; Go, A.; Greenlund, K.; Haase, N.; Ho, M.; Howard, V.; Kissela, B.; et al. Heart disease and stroke statistics—2007 update: A report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation 2007, 115, e69–e171. [Google Scholar] [CrossRef]
- Saver, J.L. Time is brain—Quantified. Stroke 2006, 37, 263–266. [Google Scholar] [CrossRef]
- Hacke, W.; Kaste, M.; Bluhmki, E.; Brozman, M.; Davalos, A.; Guidetti, D.; Larrue, V.; Lees, K.R.; Medeghri, Z.; Machnig, T.; et al. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. N. Engl. J. Med. 2008, 359, 1317–1329. [Google Scholar] [CrossRef]
- Saver, J.L.; Gornbein, J.; Grotta, J.; Liebeskind, D.; Lutsep, H.; Schwamm, L.; Scott, P.; Starkman, S. Number needed to treat to benefit and to harm for intravenous tissue plasminogen activator therapy in the 3- to 4.5-hour window: Joint outcome table analysis of the ECASS 3 trial. Stroke 2009, 40, 2433–2437. [Google Scholar] [CrossRef]
- The National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group. Tissue plasminogen activator for acute ischemic stroke. N. Engl. J. Med. 1995, 333, 1581–1587. [CrossRef]
- The National Institute of Neurological Disorders and Stroke t-PA Stroke Study Group. Intracerebral hemorrhage after intravenous t-PA therapy for ischemic stroke. Stroke 1997, 28, 2109–2118. [CrossRef]
- Dirnagl, U.; Simon, R.P.; Hallenbeck, J.M. Ischemic tolerance and endogenous neuroprotection. Trends Neurosci. 2003, 26, 248–254. [Google Scholar] [CrossRef]
- Dirnagl, U.; Iadecola, C.; Moskowitz, M.A. Pathobiology of ischaemic stroke: An integrated view. Trends Neurosci. 1999, 22, 391–397. [Google Scholar] [CrossRef]
- Zheng, Y.Q.; Liu, J.X.; Li, X.Z.; Xu, L.; Xu, Y.G. RNA interference-mediated downregulation of Beclin1 attenuates cerebral ischemic injury in rats. Acta Pharmacol. Sin. 2009, 30, 919–927. [Google Scholar] [CrossRef]
- Al-Jamal, K.T.; Gherardini, L.; Bardi, G.; Nunes, A.; Guo, C.; Bussy, C.; Herrero, M.A.; Bianco, A.; Prato, M.; Kostarelos, K.; et al. Functional motor recovery from brain ischemic insult by carbon nanotube-mediated siRNA silencing. Proc. Nat. Acad. Sci. USA 2011, 108, 10952–10957. [Google Scholar] [CrossRef]
- Kim, H.W.; Cho, K.J.; Lee, S.K.; Kim, G.W. Apoptosis signal-regulating kinase 1 (Ask1) targeted small interfering RNA on ischemic neuronal cell death. Brain Res. 2011, 1412, 73–78. [Google Scholar]
- Zhang, J.; Wang, Y.; Zhu, P.; Wang, X.; Lv, M.; Feng, H. siRNA-mediated silence of protease-activated receptor-1 minimizes ischemic injury of cerebral cortex through HSP70 and MAP2. J. Neurol. Sci. 2012, 320, 6–11. [Google Scholar] [CrossRef]
- Price, M.; Badaut, J.; Thevenet, J.; Hirt, L. Activation of c-Jun in the nuclei of neurons of the CA-1 in thrombin preconditioning occurs via PAR-1. J. Neurosci. Res. 2010, 88, 1338–1347. [Google Scholar]
- Thevenet, J.; Angelillo-Scherrer, A.; Price, M.; Hirt, L. Coagulation factor Xa activates thrombin in ischemic neural tissue. J. Neurochem. 2009, 111, 828–836. [Google Scholar] [CrossRef]
- Chen, C.; Hu, Q.; Yan, J.; Yang, X.; Shi, X.; Lei, J.; Chen, L.; Huang, H.; Han, J.; Zhang, J.H.; et al. Early inhibition of HIF-1alpha with small interfering RNA reduces ischemic-reperfused brain injury in rats. Neurobiol. Dis. 2009, 33, 509–517. [Google Scholar] [CrossRef]
- Zhao, B.; Zhao, C.Z.; Zhang, X.Y.; Huang, X.Q.; Shi, W.Z.; Fang, S.H.; Lu, Y.B.; Zhang, W.P.; Xia, Q.; Wei, E.Q. The new P2Y-like receptor G protein-coupled receptor 17 mediates acute neuronal injury and late microgliosis after focal cerebral ischemia in rats. Neuroscience 2012, 202, 42–57. [Google Scholar] [CrossRef]
- Ciana, P.; Fumagalli, M.; Trincavelli, M.L.; Verderio, C.; Rosa, P.; Lecca, D.; Ferrario, S.; Parravicini, C.; Capra, V.; Gelosa, P.; et al. The orphan receptor GPR17 identified as a new dual uracil nucleotides/cysteinyl-leukotrienes receptor. EMBO J. 2006, 25, 4615–4627. [Google Scholar] [CrossRef]
- Lecca, D.; Trincavelli, M.L.; Gelosa, P.; Sironi, L.; Ciana, P.; Fumagalli, M.; Villa, G.; Verderio, C.; Grumelli, C.; Guerrini, U.; et al. The recently identified P2Y-like receptor GPR17 is a sensor of brain damage and a new target for brain repair. PLoS One 2008, 3, e3579. [Google Scholar]
- Fukuda, A.M.; Badaut, J. Aquaporin 4: A player in cerebral edema and neuroinflammation. J. Neuroinflammation 2012, 9, 279. [Google Scholar] [CrossRef]
- Loane, D.J.; Byrnes, K.R. Role of microglia in neurotrauma. Neurotherapeutics 2010, 7, 366–377. [Google Scholar] [CrossRef]
- Bianchi, M.E.; Manfredi, A.A. High-mobility group box 1 (HMGB1) protein at the crossroads between innate and adaptive immunity. Immunol. Rev. 2007, 220, 35–46. [Google Scholar] [CrossRef]
- Kim, I.D.; Lim, C.M.; Kim, J.B.; Nam, H.Y.; Nam, K.; Kim, S.W.; Park, J.S.; Lee, J.K. Neuroprotection by biodegradable PAMAM ester (e-PAM-R)-mediated HMGB1 siRNA delivery in primary cortical cultures and in the postischemic brain. J. Control. Release 2010, 142, 422–430. [Google Scholar] [CrossRef]
- Kim, J.B.; Sig Choi, J.; Yu, Y.M.; Nam, K.; Piao, C.S.; Kim, S.W.; Lee, M.H.; Han, P.L.; Park, J.S.; Lee, J.K. HMGB1, a novel cytokine-like mediator linking acute neuronal death and delayed neuroinflammation in the postischemic brain. J. Neurosci. 2006, 26, 6413–6421. [Google Scholar] [CrossRef]
- Kim, I.D.; Shin, J.H.; Kim, S.W.; Choi, S.; Ahn, J.; Han, P.L.; Park, J.S.; Lee, J.K. Intranasal delivery of HMGB1 siRNA confers target gene knockdown and robust neuroprotection in the postischemic brain. Mol. Ther. 2012, 20, 829–839. [Google Scholar] [CrossRef]
- Thorne, R.G.; Emory, C.R.; Ala, T.A.; Frey, W.H., 2nd. Quantitative analysis of the olfactory pathway for drug delivery to the brain. Brain Res. 1995, 692, 278–282. [Google Scholar]
- Thorne, R.G.; Hanson, L.R.; Ross, T.M.; Tung, D.; Frey, W.H., 2nd. Delivery of interferon-beta to the monkey nervous system following intranasal administration. Neuroscience 2008, 152, 785–797. [Google Scholar] [CrossRef]
- Thorne, R.G.; Pronk, G.J.; Padmanabhan, V.; Frey, W.H., 2nd. Delivery of insulin-like growth factor-I to the rat brain and spinal cord along olfactory and trigeminal pathways following intranasal administration. Neuroscience 2004, 127, 481–496. [Google Scholar] [CrossRef]
- Liu, X.F.; Fawcett, J.R.; Thorne, R.G.; DeFor, T.A.; Frey, W.H., 2nd. Intranasal administration of insulin-like growth factor-I bypasses the blood-brain barrier and protects against focal cerebral ischemic damage. J. Neurol. Sci. 2001, 187, 91–97. [Google Scholar] [CrossRef]
- Faul, M.; Xu, L.; Wald, M.M.; Coronado, V. Traumatic Brain Injury in the United States: Emergency Department Visits, Hospitalizations, and Deaths,2002–2006; CDC, National Center for Injury Prevention and Control: Atlanta, GA, USA, 2010. [Google Scholar]
- Pop, V.; Badaut, J. A neurovascular perspective for long-term changes after brain trauma. Transl. Stroke Res. 2011, 2, 533–545. [Google Scholar] [CrossRef]
- Morales, D.M.; Marklund, N.; Lebold, D.; Thompson, H.J.; Pitkanen, A.; Maxwell, W.L.; Longhi, L.; Laurer, H.; Maegele, M.; Neugebauer, E.; et al. Experimental models of traumatic brain injury: Do we really need to build a better mousetrap? Neuroscience 2005, 136, 971–989. [Google Scholar] [CrossRef]
- Miyashita, R.; Chen, L.; Oshiro, H.; Uchino, H.; Shibasaki, F. Int6 silencing causes induction of angiogenic factors in neuronal cells via accumulation of hypoxia-inducible factor 2alpha and decreases brain damage in rats. Neurosci. Lett. 2012, 528, 83–88. [Google Scholar] [CrossRef]
- Woodcock, T.; Morganti-Kossmann, M.C. The role of markers of inflammation in traumatic brain injury. Front. Neurol. 2013, 4, 18. [Google Scholar]
- Campbell, M.; Hanrahan, F.; Gobbo, O.L.; Kelly, M.E.; Kiang, A.S.; Humphries, M.M.; Nguyen, A.T.; Ozaki, E.; Keaney, J.; Blau, C.W.; et al. Targeted suppression of claudin-5 decreases cerebral oedema and improves cognitive outcome following traumatic brain injury. Nat. Commun. 2012, 3, 849. [Google Scholar] [CrossRef]
- Badaut, J.; Ashwal, S.; Obenaus, A. Aquaporins in cerebrovascular disease: A target for treatment of brain edema? Cerebrovasc. Dis. 2011, 31, 521–531. [Google Scholar] [CrossRef]
- Fukuda, A.M.; Pop, V.; Spagnoli, D.; Ashwal, S.; Obenaus, A.; Badaut, J. Delayed increase of astrocytic aquaporin 4 after juvenile traumatic brain injury: Possible role in edema resolution? Neuroscience 2012, 222, 366–378. [Google Scholar] [CrossRef]
- Fukuda, A.M.; Adami, A.; Pop, V.; Bellone, J.A.; Coats, J.S.; Hartman, R.E.; Ashwal, S.; Obenaus, A.; Badaut, J. Posttraumatic reduction of edema with aquaporin-4 RNA interference improves acute and chronic functional recovery. J. Cerebr. Blood Flow Metabol. 2013. [Google Scholar] [CrossRef]
- Niu, L.J.; Xu, R.X.; Zhang, P.; Du, M.X.; Jiang, X.D. Suppression of Frizzled-2-mediated Wnt/Ca(2)(+) signaling significantly attenuates intracellular calcium accumulation in vitro and in a rat model of traumatic brain injury. Neuroscience 2012, 213, 19–28. [Google Scholar] [CrossRef]
- Schanne, F.A.; Kane, A.B.; Young, E.E.; Farber, J.L. Calcium dependence of toxic cell death: A final common pathway. Science 1979, 206, 700–702. [Google Scholar]
- Mathupala, S.P. Delivery of small-interfering RNA (siRNA) to the brain. Expert Opin. Ther. Patents 2009, 19, 137–140. [Google Scholar] [CrossRef]
- Zhou, J.; Shum, K.T.; Burnett, J.C.; Rossi, J.J. Nanoparticle-based delivery of RNAi therapeutics: Progress and challenges. Pharmaceuticals 2013, 6, 85–107. [Google Scholar] [CrossRef]
- Shim, M.S.; Kwon, Y.J. Efficient and targeted delivery of siRNA in vivo. FEBS J. 2010, 277, 4814–4827. [Google Scholar] [CrossRef]
- Van de Water, F.M.; Boerman, O.C.; Wouterse, A.C.; Peters, J.G.; Russel, F.G.; Masereeuw, R. Intravenously administered short interfering RNA accumulates in the kidney and selectively suppresses gene function in renal proximal tubules. Drug Metab. Dispos. 2006, 34, 1393–1397. [Google Scholar] [CrossRef]
- Walton, S.P.; Wu, M.; Gredell, J.A.; Chan, C. Designing highly active siRNAs for therapeutic applications. FEBSJ. 2010, 277, 4806–4813. [Google Scholar] [CrossRef]
- Danielyan, L.; Schafer, R.; von Ameln-Mayerhofer, A.; Buadze, M.; Geisler, J.; Klopfer, T.; Burkhardt, U.; Proksch, B.; Verleysdonk, S.; Ayturan, M.; et al. Intranasal delivery of cells to the brain. Eur. J. Cell Biol. 2009, 88, 315–324. [Google Scholar] [CrossRef]
- Danielyan, L.; Klein, R.; Hanson, L.R.; Buadze, M.; Schwab, M.; Gleiter, C.H.; Frey, W.H. Protective effects of intranasal losartan in the APP/PS1 transgenic mouse model of Alzheimer disease. Rejuv. Res. 2010, 13, 195–201. [Google Scholar] [CrossRef]
- Gomez, D.; Martinez, J.A.; Hanson, L.R.; Frey, W.H., 2nd.; Toth, C.C. Intranasal treatment of neurodegenerative diseases and stroke. Front. Biosci. 2012, 4, 74–89. [Google Scholar]
- Hanson, L.R.; Frey, W.H., 2nd. Intranasal delivery bypasses the blood-brain barrier to target therapeutic agents to the central nervous system and treat neurodegenerative disease. BMC Neurosci. 2008, 9, S5. [Google Scholar] [CrossRef]
- Hanson, L.R.; Roeytenberg, A.; Martinez, P.M.; Coppes, V.G.; Sweet, D.C.; Rao, R.J.; Marti, D.L.; Hoekman, J.D.; Matthews, R.B.; Frey, W.H., 2nd.; et al. Intranasal deferoxamine provides increased brain exposure and significant protection in rat ischemic stroke. J. Pharmacol. Exp. Ther. 2009, 330, 679–686. [Google Scholar] [CrossRef]
- Liu, X.F.; Fawcett, J.R.; Thorne, R.G.; Frey, W.H., 2nd. Non-invasive intranasal insulin-like growth factor-I reduces infarct volume and improves neurologic function in rats following middle cerebral artery occlusion. Neurosci. Lett. 2001, 308, 91–94. [Google Scholar]
- Wei, G.; Wang, D.; Lu, H.; Parmentier, S.; Wang, Q.; Panter, S.S.; Frey, W.H., 2nd.; Ying, W. Intranasal administration of a PARG inhibitor profoundly decreases ischemic brain injury. Front. Biosci. 2007, 12, 4986–4996. [Google Scholar] [CrossRef]
- Renner, D.B.; Frey, W.H., 2nd.; Hanson, L.R. Intranasal delivery of siRNA to the olfactory bulbs of mice via the olfactory nerve pathway. Neurosci. Lett. 2012, 513, 193–197. [Google Scholar] [CrossRef]
- Bortolozzi, A.; Castane, A.; Semakova, J.; Santana, N.; Alvarado, G.; Cortes, R.; Ferres-Coy, A.; Fernandez, G.; Carmona, M.C.; Toth, M.; et al. Selective siRNA-mediated suppression of 5-HT1A autoreceptors evokes strong anti-depressant-like effects. Mol. Psychiatr. 2012, 17, 612–623. [Google Scholar]
- Lu, X.X.; McCoy, K.S.; Xu, J.L.; Hu, W.K.; Chen, H.B. Small interfering RNA targeting T-cell Ig mucin-3 decreases allergic airway inflammation and hyperresponsiveness. Inflammation 2013, 36, 582–591. [Google Scholar] [CrossRef]
- Massaro, D.; Massaro, G.D.; Clerch, L.B. Noninvasive delivery of small inhibitory RNA and other reagents to pulmonary alveoli in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L1066–L1070. [Google Scholar] [CrossRef]
- Thorne, R.G.; Frey, W.H., 2nd. Delivery of neurotrophic factors to the central nervous system: Pharmacokinetic considerations. Clin. Pharmacokinet. 2001, 40, 907–946. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar]
- El Andaloussi, S.; Lakhal, S.; Mager, I.; Wood, M.J. Exosomes for targeted siRNA delivery across biological barriers. Adv. Drug Deliv. Rev. 2013, 65, 391–397. [Google Scholar] [CrossRef]
- Wahlgren, J.; Karlson, T.D.L.; Brisslert, M.; Vaziri Sani, F.; Telemo, E.; Sunnerhagen, P.; Valadi, H. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucl. Acids Res. 2012, 40, e130. [Google Scholar] [CrossRef]
- Cossetti, C.; Smith, J.A.; Iraci, N.; Leonardi, T.; Alfaro-Cervello, C.; Pluchino, S. Extracellular membrane vesicles and immune regulation in the brain. Front. Physiol. 2012, 3, 117. [Google Scholar]
- Lai, C.P.; Breakefield, X.O. Role of exosomes/microvesicles in the nervous system and use in emerging therapies. Front. Physiol. 2012, 3, 228. [Google Scholar]
- Badaut, J.; Ashwal, S.; Adami, A.; Tone, B.; Recker, R.; Spagnoli, D.; Ternon, B.; Obenaus, A. Brain water mobility decreases after astrocytic aquaporin-4 inhibition using RNA interference. J. Cerebr. Blood Flow Metabol. 2011, 31, 819–831. [Google Scholar] [CrossRef]
- Davis, M.E.; Chen, Z.G.; Shin, D.M. Nanoparticle therapeutics: An emerging treatment modality for cancer. Nat. Rev. Drug Discov. 2008, 7, 771–782. [Google Scholar] [CrossRef]
- Petros, R.A.; DeSimone, J.M. Strategies in the design of nanoparticles for therapeutic applications. Nat. Rev. Drug Discov. 2010, 9, 615–627. [Google Scholar] [CrossRef]
- Mishima, T.; Mizuguchi, Y.; Kawahigashi, Y.; Takizawa, T.; Takizawa, T. RT-PCR-based analysis of microRNA (miR-1 and -124) expression in mouse CNS. Brain Res. 2007, 1131, 37–43. [Google Scholar] [CrossRef]
- Saugstad, J.A. MicroRNAs as effectors of brain function with roles in ischemia and injury, neuroprotection, and neurodegeneration. J. Cerebr. Blood Flow Metabol. 2010, 30, 1564–1576. [Google Scholar] [CrossRef]
- Linsen, S.E.; de Wit, E.; de Bruijn, E.; Cuppen, E. Small RNA expression and strain specificity in the rat. BMC Genomics 2010, 11, 249. [Google Scholar] [CrossRef]
- Hu, Z.; Yu, D.; Almeida-Suhett, C.; Tu, K.; Marini, A.M.; Eiden, L.; Braga, M.F.; Zhu, J.; Li, Z. Expression of miRNAs and their cooperative regulation of the pathophysiology in traumatic brain injury. PLoS One 2012, 7, e39357. [Google Scholar]
- Liu, N.K.; Xu, X.M. MicroRNA in central nervous system trauma and degenerative disorders. Physiol. Genomics 2011, 43, 571–580. [Google Scholar] [CrossRef]
- Ziu, M.; Fletcher, L.; Rana, S.; Jimenez, D.F.; Digicaylioglu, M. Temporal differences in microRNA expression patterns in astrocytes and neurons after ischemic injury. PLoS One 2011, 6, e14724. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fukuda, A.M.; Badaut, J. siRNA Treatment: “A Sword-in-the-Stone” for Acute Brain Injuries. Genes 2013, 4, 435-456. https://0-doi-org.brum.beds.ac.uk/10.3390/genes4030435
Fukuda AM, Badaut J. siRNA Treatment: “A Sword-in-the-Stone” for Acute Brain Injuries. Genes. 2013; 4(3):435-456. https://0-doi-org.brum.beds.ac.uk/10.3390/genes4030435
Chicago/Turabian StyleFukuda, Andrew M., and Jerome Badaut. 2013. "siRNA Treatment: “A Sword-in-the-Stone” for Acute Brain Injuries" Genes 4, no. 3: 435-456. https://0-doi-org.brum.beds.ac.uk/10.3390/genes4030435