
Human Specific Regulation of the Telomerase Reverse Transcriptase Gene

Abstract
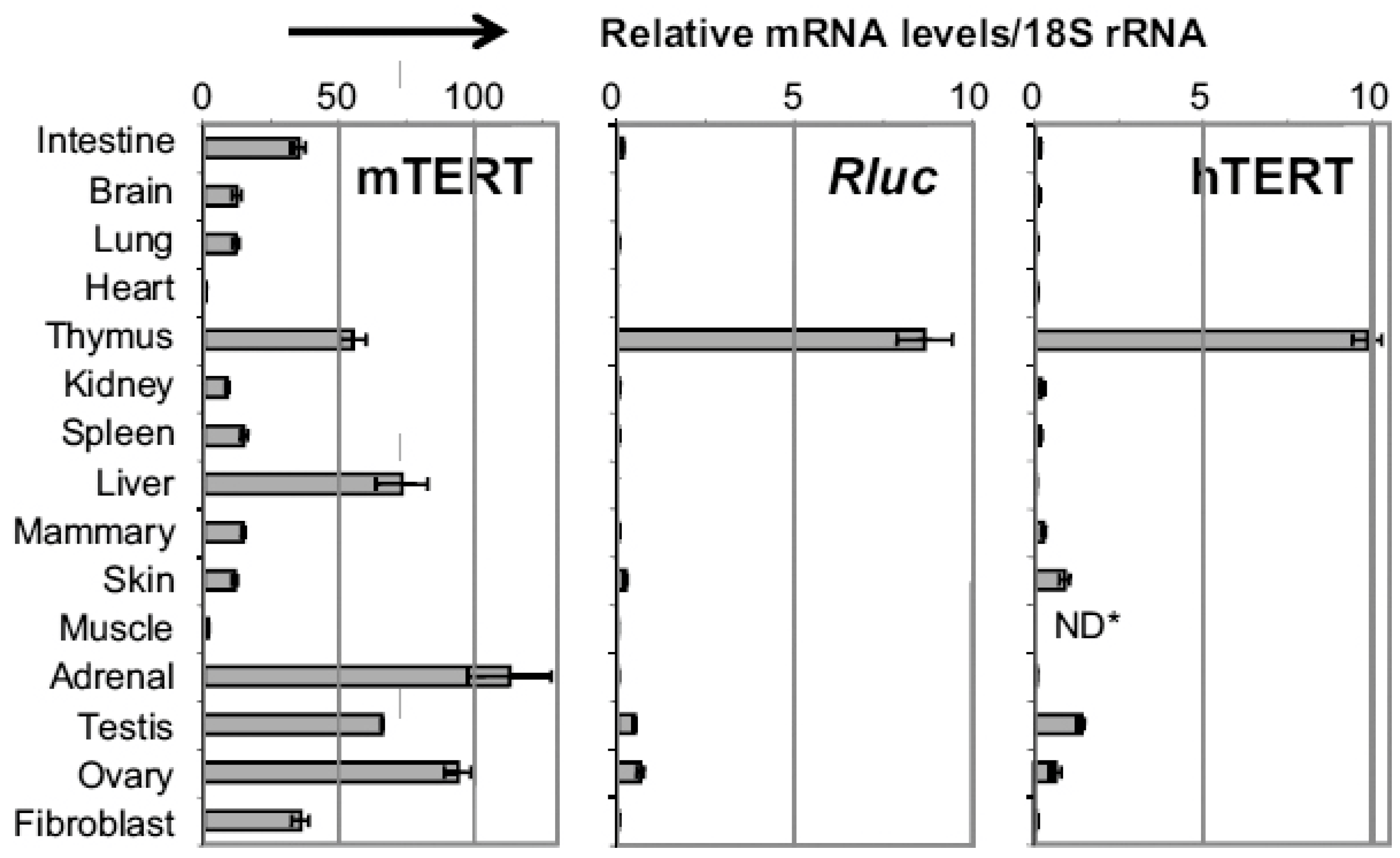
:1. Introduction
2. Regulation of TERT Genes
3. Genomic Sequences Underlining the Differential Regulation of hTERT and mTERT Genes
4. Transcriptional Regulation of hTERT Promoter
4.1. Transcriptional Factors Involved in hTERT Regulation
4.1.1. Myc Family TFs
4.1.2. E2F1
4.1.3. TGF-β signaling
4.2. Histone Acetylation and Deacetylation
4.3. DNA and Histone Methylation
4.4. hTERT Regulation in Cancer Cells
5. Single Nucleotide Polymorphisms (SNPs) at the hTERT Locus
6. Chromosomal Rearrangement at the hTERT Locus
7. Conclusions
Acknowledgments
Conflicts of interest
References
- Venteicher, A.S.; Abreu, E.B.; Meng, Z.; McCann, K.E.; Terns, R.M.; Veenstra, T.D.; Terns, M.P.; Artandi, S.E. A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis. Science 2009, 323, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.S.; Wright, W.E.; Shay, J.W. Human telomerase and its regulation. Microbiol. Mol. Biol. Rev. 2002, 66, 407–425. [Google Scholar] [CrossRef] [PubMed]
- Wright, W.E.; Shay, J.W. Telomere dynamics in cancer progression and prevention: Fundamental differences in human and mouse telomere biology. Nat. Med. 2000, 6, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Akincilar, S.C.; Unal, B.; Tergaonkar, V. Reactivation of telomerase in cancer. Cell Mol. Life Sci. 2016, 73, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.M.; Ryder, O.A.; Houck, M.L.; Charter, S.J.; Walker, W.; Forsyth, N.R.; Austad, S.N.; Venditti, C.; Pagel, M.; Shay, J.W.; et al. Comparative biology of mammalian telomeres: Hypotheses on ancestral states and the roles of telomeres in longevity determination. Aging Cell 2011, 10, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, I.; Chiang, Y.J.; Patterson, T.; Feigenbaum, L.; Leem, S.H.; Michishita, E.; Larionov, V.; Hodes, R.J.; Barrett, J.C. Differential cis-regulation of human versus mouse TERT gene expression in vivo: Identification of a human-specific repressive element. Proc. Natl. Acad. Sci. USA 2005, 102, 18437–18442. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wang, S.; Horner, J.W.; Wang, N.; Wang, H.; Gunther, E.J.; DePinho, R.A.; Zhu, J. A BAC transgenic reporter recapitulates in vivo regulation of human telomerase reverse transcriptase in development and tumorigenesis. FASEB J. 2011, 25, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Marti’N-Rivera, L.; Herrera, E.S.; Albar, J.P.; Blasco, M.A.A. Expression of mouse telomerase catalytic subunit in embryos and adult tissues. Proc. Natl. Acad. Sci. USA 1998, 95, 10471–10477. [Google Scholar] [CrossRef]
- Prowse, K.R.; Greider, C.W. Developmental and tissue-specific regulation of mouse telomerase and telomere length. Proc. Natl. Acad. Sci. USA 1995, 92, 4818–4822. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, R.A.; Allsopp, R.C.; Chin, L.; Morin, G.B.; DePinho, R.A. Expression of mouse telomerase reverse transcriptase during development, differentiation and proliferation. Oncogene 1998, 16, 1723–1730. [Google Scholar] [PubMed]
- Kipling, D.; Cooke, H.J. Hypervariable ultra-long telomeres in mice. Nature 1990, 347, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Murriel, C.L.; Johnson, L. Genetically engineered mouse models: Closing the gap between preclinical data and trial outcomes. Cancer Res. 2012, 72, 2695–2700. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, A.; Weinberg, R.A. Opinion: Comparative biology of mouse versus human cells: Modelling human cancer in mice. Nat. Rev. Cancer 2003, 3, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Aisner, D.L.; Wright, W.E.; Shay, J.W. Telomerase regulation: Not just flipping the switch. Curr. Opin. Genet. Dev. 2002, 12, 80–85. [Google Scholar] [CrossRef]
- Takakura, M.; Kyo, S.; Kanaya, T.; Hirano, H.; Takeda, J.; Yutsudo, M.; Inoue, M. Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res. 1999, 59, 551–557. [Google Scholar] [PubMed]
- Wu, K.J.; Grandori, C.; Amacker, M.; Simon-Vermot, N.; Polack, A.; Lingner, J.; Dalla-Favera, R. Direct activation of TERT transcription by c-MYC. Nat. Genet. 1999, 21, 220–224. [Google Scholar] [PubMed]
- Zhu, J.; Zhao, Y.; Wang, S. Chromatin and epigenetic regulation of the telomerase reverse transcriptase gene. Protein Cell 2010, 1, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, J. The hTERT gene is embedded in a nuclease-resistant chromatin domain. J. Biol. Chem. 2004, 279, 55401–55410. [Google Scholar] [CrossRef] [PubMed]
- Ducrest, A.L.; Amacker, M.; Mathieu, Y.D.; Cuthbert, A.P.; Trott, D.A.; Newbold, R.F.; Nabholz, M.; Lingner, J. Regulation of human telomerase activity: Repression by normal chromosome 3 abolishes nuclear telomerase reverse transcriptase transcripts but does not affect c-Myc activity. Cancer Res. 2001, 61, 7594–7602. [Google Scholar] [PubMed]
- Wang, S.; Zhu, J. Evidence for a relief of repression mechanism for activation of the human telomerase reverse transcriptase promoter. J. Biol. Chem. 2003, 278, 18842–18850. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, C.; Zhu, J. Transcriptional silencing of a novel hTERT reporter locus during in vitro differentiation of mouse embryonic stem cells. Mol. Biol. Cell. 2007, 18, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of life-span by introduction of telomerase into normal human cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, Y.; Hu, C.; Zhu, J. Differential repression of human and mouse TERT genes during cell differentiation. Nucleic Acids Res. 2009, 37, 2618–2629. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.D.; Barbosa-Morais, N.L.; Schmidt, D.; Conboy, C.M.; Vanes, L.; Tybulewicz, V.L.; Fisher, E.M.; Tavare, S.; Odom, D.T. Species-specific transcription in mice carrying human chromosome 21. Science 2008, 322, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, I.; Barrett, J.C. Transcriptional regulation of the telomerase hTERT gene as a target for cellular and viral oncogenic mechanisms. Carcinogenesis 2003, 24, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Popov, N.; Hou, M.; Wang, Q.; Bjorkholm, M.; Gruber, A.; Menkel, A.R.; Henriksson, M. Switch from Myc/Max to Mad1/Max binding and decrease in histone acetylation at the telomerase reverse transcriptase promoter during differentiation of HL60 cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3826–3831. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cheng, D.; Wang, S.; Zhu, J. Dual roles of c-Myc in the regulation of hTERT gene. Nucleic Acids Res. 2014, 42, 10385–10398. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Elledge, S.J. Multiple tumor suppressor pathways negatively regulate telomerase. Cell 2003, 113, 881–889. [Google Scholar] [CrossRef]
- Gunes, C.; Lichtsteiner, S.; Vasserot, A.P.; Englert, C. Expression of the hTERT gene is regulated at the level of transcriptional initiation and repressed by Mad1. Cancer Res. 2000, 60, 2116–2121. [Google Scholar] [PubMed]
- Han, S.; Park, K.; Kim, H.Y.; Lee, M.S.; Kim, H.J.; Kim, Y.D. Expression of Mad1 protein inhibits proliferation of cancer cells and inversely correlated with Myc protein expression in primary gastric cancer. Oncol. Rep. 1999, 6, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.K.; Lee, D.F.; Sun, H.L.; Li, L.Y.; Lin, C.Y.; Huang, W.C.; Hsu, J.M.; Kuo, H.P.; Yamaguchi, H.; Wang, Y.N.; et al. The suppression of MAD1 by AKT-mediated phosphorylation activates MAD1 target genes transcription. Mol. Carcinog. 2009, 48, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Crowe, D.L.; Nguyen, D.C.; Tsang, K.J.; Kyo, S. E2F-1 represses transcription of the human telomerase reverse transcriptase gene. Nucleic Acids Res. 2001, 29, 2789–2794. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Seo, H.W.; Jung, E.S.; Kim, B.H.; Jung, G. The TERT promoter SNP rs2853669 decreases E2F1 transcription factor binding and increases mortality and recurrence risks in liver cancer. Oncotarget 2016, 7, 684–699. [Google Scholar] [CrossRef] [PubMed]
- Coller, H.A.; Forman, J.J.; Legesse-Miller, A. “Myc’ed Messages”: Myc Induces Transcription of E2F1 while Inhibiting Its Translation via a microRNA Polycistron. PLoS Genet. 2007, 3, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, L.; Yang, S.; Fang, D. E2F1: A potential negative regulator of hTERT transcription in normal cells upon activation of oncogenic c-Myc. Med. Sci. Monit. 2012, 18, RA12–RA15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, A.; Shen, C.; Zhang, B.; Rao, Z.; Wang, R.; Yang, S.; Ning, S.; Mao, G.; Fang, D. E2F1 acts as a negative feedback regulator of c-Mycinduced hTERT transcription during tumorigenesis. Oncol. Rep. 2014, 32, 1273–1280. [Google Scholar] [PubMed]
- Alonso, M.M.; Fueyo, J.; Yung, W.K.; Gomez-Manzano, C. E2F1 and telomerase: alliance in the dark side. Cell Cycle 2006, 5, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.M.; Fueyo, J.; Shay, J.W.; Aldape, K.D.; Jiang, H.; Lee, O.H.; Johnson, D.G.; Xu, J.; Kondo, Y.; Kanzawa, T.; et al. Expression of transcription factor E2F1 and telomerase in glioblastomas: Mechanistic linkage and prognostic significance. J. Natl. Cancer Inst. 2005, 97, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.M.; Fueyo, J.; Gomez-Manzano, C. Malignant Gliomas: Role of E2F1 Transcription Factor. Tumors Cent. Nervous Syst. 2011, 1, 89–97. [Google Scholar]
- Yoo, Y.S.; Park, S.; Gwak, J.; Ju, B.G.; Oh, S. Involvement of transcription repressor Snail in the regulation of human telomerase reverse transcriptase (hTERT) by transforming growth factor-beta. Biochem. Biophys. Res. Commun. 2015, 465, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Fujiwara, T.; Inoue, M. Understanding and exploiting hTERT promoter regulation for diagnosis and treatment of human cancers. Cancer Sci. 2008, 99, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, C.; Zhu, J. Distinct and temporal roles of nucleosomal remodeling and histone deacetylation in the repression of the hTERT gene. Mol. Biol. Cell. 2010, 21, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.; Peek, G.W.; Tollefsbol, T.O. Regulation of the human catalytic subunit of telomerase (hTERT). Gene 2012, 498, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.S.; Bacchetti, S. Histone deacetylation is involved in the transcriptional repression of hTERT in normal human cells. J. Biol. Chem. 2000, 275, 35665–35668. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Zhao, Y.; Wang, S.; Jia, W.; Kang, J.; Zhu, J. Human Telomerase Reverse Transcriptase (hTERT) Transcription Requires Sp1/Sp3 Binding to the Promoter and a Permissive Chromatin Environment. J. Biol. Chem. 2015, 290, 30193–30203. [Google Scholar] [CrossRef] [PubMed]
- Bechter, O.E.; Eisterer, W.; Dlaska, M.; Kühr, T.; Thaler, J. CpG island methylation of the hTERT promoter is associated with lower telomerase activity in B-cell lymphocytic leukemia. Exp. Hematol. 2002, 30, 26–33. [Google Scholar] [CrossRef]
- Fan, Y.; Lee, S.; Wu, G.; Easton, J.; Yergeau, D.; Dummer, R.; Vogel, P.; Kirkwood, J.M.; Barnhill, R.L.; Pappo, A.; et al. Telomerase Expression by Aberrant Methylation of the TERT Promoter in Melanoma Arising in Giant Congenital Nevi. J. Investig. Dermatol. 2016, 136, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhou, D.; Zhang, J.; Chen, Y.; Wang, C.; Liu, Y.; Zhao, K. Aberrant methylation of the TERT promoter in esophageal squamous cell carcinoma. Cancer Genet. 2015, 208, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Kochling, M.; Ewelt, C.; Furtjes, G.; Peetz-Dienhart, S.; Koos, B.; Hasselblatt, M.; Paulus, W.; Stummer, W.; Brokinkel, B. hTERT promoter methylation in pituitary adenomas. Brain Tumor Pathol. 2016, 33, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.; Jia, W.; Sharma, A.; Zhao, Y.; Clarke, L.E.; Cheng, X.; Wang, H.; Salli, U.; Vrana, K.E.; Robertson, G.P.; et al. Robust activation of the human but not mouse telomerase gene during the induction of pluripotency. FASEB J. 2010, 24, 2702–2715. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.P.; Hoare, S.F.; Glasspool, R.M.; Keith, W.N. Lack of telomerase gene expression in alternative lengthening of telomere cells is associated with chromatin remodeling of the hTR and hTERT gene promoters. Cancer Res. 2005, 65, 7585–7590. [Google Scholar] [PubMed]
- Zinn, R.L.; Pruitt, K.; Eguchi, S.; Baylin, S.B.; Herman, J.G. hTERT is expressed in cancer cell lines despite promoter DNA methylation by preservation of unmethylated DNA and active chromatin around the transcription start site. Cancer Res. 2007, 67, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Taira, T.; Kanaya, T.; Itoh, H.; Yutsudo, M.; Ariga, H.; Inoue, M. Sp1 cooperates with c-Myc to activate transcription of the human telomerase reverse transcriptase gene (hTERT). Nucleic Acids Res. 2000, 28, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Kanaya, T.; Zhuo, W.; Fujimoto, K.; Nishio, Y.; Orimo, A.; Inoue, M. Estrogen activates telomerase. Cancer Res. 1999, 59, 5917–5921. [Google Scholar] [PubMed]
- Kondoh, K.; Tsuji, N.; Asanuma, K.; Kobayashi, D.; Watanabe, N. Inhibition of estrogen receptor beta-mediated human telomerase reverse transcriptase gene transcription via the suppression of mitogen-activated protein kinase signaling plays an important role in 15-deoxy-Delta(12,14)-prostaglandin J(2)-induced apoptosis in cancer cells. Exp. Cell. Res. 2007, 313, 3486–3496. [Google Scholar] [PubMed]
- Grasselli, A.; Nanni, S.; Colussi, C.; Aiello, A.; Benvenuti, V.; Ragone, G.; Moretti, F.; Sacchi, A.; Bacchetti, S.; Gaetano, C.; et al. Estrogen receptor-alpha and endothelial nitric oxide synthase nuclear complex regulates transcription of human telomerase. Circ. Res. 2008, 103, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Veldman, T.; Liu, X.; Yuan, H.; Schlegel, R. Human papillomavirus E6 and Myc proteins associate in vivo and bind to and cooperatively activate the telomerase reverse transcriptase promoter. Proc. Natl. Acad. Sci. USA 2003, 100, 8211–8216. [Google Scholar] [CrossRef] [PubMed]
- Katzenellenbogen, R.A.; Vliet-Gregg, P.; Xu, M.; Galloway, D.A. NFX1-123 increases hTERT expression and telomerase activity posttranscriptionally in human papillomavirus type 16 E6 keratinocytes. J. Virol. 2009, 83, 6446–6456. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Xi, L.; Zaug, A.J.; Powell, N.M.; Dancik, G.; Cohen, S.B.; Costello, J.C.; Theodorescu, D.; Cech, T.R. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science 2015, 347, 1006–1010. [Google Scholar] [CrossRef] [PubMed]
- Muzza, M.; Colombo, C.; Rossi, S.; Tosi, D.; Cirello, V.; Perrino, M.; de Leo, S.; Magnani, E.; Pignatti, E.; Vigo, B.; et al. Telomerase in differentiated thyroid cancer: Promoter mutations, expression and localization. Mol. Cell Endocrinol. 2015, 399, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Zagni, C.; Chiacchi, U.; Rescifina, A. Histone Methyltransferase Inhibitors: Novel Epigenetic Agents for Cancer Treatment. Curr. Med. Chem. 2013, 20, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Schmidt, J.C.; Zaug, A.J.; Ascarrunz, D.R.; Cech, T.R. A novel two-step genome editing strategy with CRISPR-Cas9 provides new insights into telomerase action and TERT gene expression. Genome Biol. 2015, 16, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.J.; Rube, H.T.; Kreig, A.; Mancini, A.; Fouse, S.D.; Nagarajan, R.P.; Choi, S.; Hong, C.; He, D.; Pekmezci, M.; et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science 2015, 348, 1036–1039. [Google Scholar]
- Stern, J.L.; Theodorescu, D.; Vogelstein, B.; Papadopoulos, N.; Cech, T.R. Mutation of the TERT promoter, switch to active chromatin, and monoallelic TERT expression in multiple cancers. Genes Dev. 2015, 29, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.Y.; et al. Non-canonical NF-kappaB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat. Cell Biol. 2015, 17, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Bielski, C.M.; Rinne, M.L.; Hahn, W.C.; Sellers, W.R.; Stegmeier, F.; Garraway, L.A.; Kryukov, G.V. TERT promoter mutations and monoallelic activation of TERT in cancer. Oncogenesis 2015. [Google Scholar] [CrossRef] [PubMed]
- Bojesen, S.E.; Pooley, K.A.; Johnatty, S.E.; Beesley, J.; Michailidou, K.; Tyrer, J.P.; Edwards, S.L.; Pickett, H.A.; Shen, H.C.; Smart, C.E.; et al. Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat. Genet. 2013, 45, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Qiao, N.; Wang, Y.; Jiang, M.; Wang, S.; Wang, C.; Hu, L. Association between the telomerase reverse transcriptase (TERT) rs2736098 polymorphism and cancer risk: evidence from a case-control study of non-small-cell lung cancer and a meta-analysis. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tian, Y.P.; Wang, Y.; Guo, F.H.; Qin, J.F.; Ni, H. hTERT rs2736098 genetic variants and susceptibility of hepatocellular carcinoma in the Chinese population: A case-control study. Hepatobiliary Pancreat Dis. Int. 2013, 12, 74–79. [Google Scholar] [CrossRef]
- Zhang, X.J.; Xu, Z.; Gong, Y.L.; Tang, C.J.; Chen, J.F. Association of TERT rs2736098 polymorphism with cancer risk: A meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 4943–4946. [Google Scholar] [CrossRef]
- Zhou, P.; Wei, L.; Xia, X.; Shao, N.; Qian, X.; Yang, Y. Association between telomerase reverse transcriptase rs2736100 polymorphism and risk of glioma. J. Surg. Res. 2014, 191, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Cao, L.; Pu, H.; Wang, H.; Zheng, Y.; Niu, X.; Weng, X.; Zhang, H.; Favus, M.J.; Zhang, L.; et al. TERT Polymorphism rs2736100-C Is Associated with EGFR Mutation-Positive Non-Small Cell Lung Cancer. Clin. Cancer Res. 2015, 21, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Kinnersley, B.; Migliorini, G.; Broderick, P.; Whiffin, N.; Dobbins, S.E.; Casey, G.; Hopper, J.; Sieber, O.; Lipton, L.; Kerr, D.J.; et al. The TERT variant rs2736100 is associated with colorectal cancer risk. Br. J. Cancer 2012, 107, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Cawthon, R.; Gao, Y.; Hu, W.; Hosgood, H.D., 3rd; Barone-Adesi, F.; Ji, B.T.; Bassig, B.; Chow, W.H.; Shu, X.; et al. Longer telomere length in peripheral white blood cells is associated with risk of lung cancer and the rs2736100 (CLPTM1L-TERT) polymorphism in a prospective cohort study among women in China. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Kote-Jarai, Z.; Saunders, E.J.; Leongamornlert, D.A.; Tymrakiewicz, M.; Dadaev, T.; Jugurnauth-Little, S.; Ross-Adams, H.; Al Olama, A.A.; Benlloch, S.; Halim, S.; et al. Fine-mapping identifies multiple prostate cancer risk loci at 5p15, one of which associates with TERT expression. Hum. Mol. Genet. 2013. [Google Scholar] [CrossRef]
- Batista, R.; Cruvinel-Carloni, A.; Vinagre, J.; Peixoto, J.; Catarino, T.A.; Campanella, N.C.; Menezes, W.; Becker, A.P.; de Almeida, G.C.; Matsushita, M.M.; et al. The prognostic impact of TERT promoter mutations in glioblastomas is modified by the rs2853669 single nucleotide polymorphism. Int. J. Cancer 2016, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, P.S.; Hosen, I.; de Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly Recurrent TERT Promoter Mutations in Human Melanoma. Science 2013, 339, 957–958. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Hosen, I.; Gousias, K.; Rachakonda, S.; Heidenreich, B.; Gessi, M.; Schramm, J.; Hemminki, K.; Waha, A.; Kumar, R. TERT promoter mutations: A novel independent prognostic factor in primary glioblastomas. Neuro-Oncol. 2015, 17, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Almeida, A.; Populo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosrati, M.A.; Malmström, A.; Lysiak, M.; Krysztofiak, A.; Hallbeck, M.; Milos, P.; Hallbeck, A.-L. TERT promoter mutations and polymorphisms as prognostic factors in primary glioblastoma. Oncotarget 2015, 6, 16663–16673. [Google Scholar] [CrossRef] [PubMed]
- Nencha, U.; Rahimian, A.; Giry, M.; Sechi, A.; Mokhtari, K.; Polivka, M.; Schmitt, Y.; di Stefano, A.L.; Alentorn, A.; Labussiere, M.; et al. TERT promoter mutations and rs2853669 polymorphism: Prognostic impact and interactions with common alterations in glioblastomas. J. Neurooncol. 2016, 126, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; Sitaram, R.T.; Ljungberg, B.; Roos, G.; Hemminki, K.; Kumar, R. TERT promoter mutations in clear cell renal cell carcinoma. Int. J. Cancer 2015, 136, 2448–2452. [Google Scholar] [CrossRef] [PubMed]
- Catarino, R.; Araujo, A.; Coelho, A.; Gomes, M.; Nogueira, A.; Lopes, C.; Medeiros, R.M. Prognostic significance of telomerase polymorphism in non-small cell lung cancer. Clin. Cancer Res. 2010, 16, 3706–3712. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Tong, N.; Tao, G.; Luo, D.; Wang, M.; Fang, Y.; Li, J.; Xu, M.; Zhang, Z.; Wu, D. TERT polymorphisms modify the risk of acute lymphoblastic leukemia in Chinese children. Carcinogenesis 2013, 34, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Carvajal-Carmona, L.G.; O’Mara, T.A.; Painter, J.N.; Lose, F.A.; Dennis, J.; Michailidou, K.; Tyrer, J.P.; Ahmed, S.; Ferguson, K.; Healey, C.S.; et al. Candidate locus analysis of the TERT-CLPTM1L cancer risk region on chromosome 5p15 identifies multiple independent variants associated with endometrial cancer risk. Hum. Genet. 2015, 134, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Scherf, D.B.; Sarkisyan, N.; Jacobsson, H.; Claus, R.; Bermejo, J.L.; Peil, B.; Gu, L.; Muley, T.; Meister, M.; Dienemann, H.; et al. Epigenetic screen identifies genotype-specific promoter DNA methylation and oncogenic potential of CHRNB4. Oncogene 2013, 32, 3329–3338. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, S.; Popova, E.Y.; Grigoryev, S.A.; Zhu, J. Rearrangement of upstream sequences of the hTERT gene during cellular immortalization. Genes Chromosomes Cancer 2009, 48, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Cobrinik, D.; Ostrovnaya, I.; Hassimi, M.; Tickoo, S.K.; Cheung, I.Y.; Cheung, N.K. Recurrent pre-existing and acquired DNA copy number alterations, including focal TERT gains, in neuroblastoma central nervous system metastases. Genes Chromosomes Cancer 2013, 52, 1150–1166. [Google Scholar] [CrossRef] [PubMed]
- Peifer, M.; Hertwig, F.; Roels, F.; Dreidax, D.; Gartlgruber, M.; Menon, R.; Kramer, A.; Roncaioli, J.L.; Sand, F.; Heuckmann, J.M.; et al. Telomerase activation by genomic rearrangements in high-risk neuroblastoma. Nature 2015, 526, 700–704. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Site | Position (GRCh37) | A1/A2 | Cancer risk | References |
|---|---|---|---|---|
| 124(C/T) | 1,295,228 | C/T | multiple tumors, like melanoma glioblastomas | [59,79,80,81] |
| 146(C/T) | 1,295,250 | C/T(CC/TT) | ||
| rs2853669 | 1,295,349 | T/C | glioblastomas | [77,82,83] |
| thyroid cancer | [61] | |||
| bladder cancer | [78] | |||
| clear cell renal cell carcinoma | [84] | |||
| hepatocellular carcinoma | [33] | |||
| rs2735940 | 1,296,486 | T/C | lung cancer | [85] |
| childhood acute lymphoblastic leukemia | [86] | |||
| rs2736108 | 1,297,488 | C/T | longer telomere | [68] |
| rs13174814 | 1,299,859 | G/C | endometrial cancer | [87] |
| rs421629 | 1,320,136 | C/T | lung cancer | [88] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Cheng, D.; Wang, S.; Zhu, J. Human Specific Regulation of the Telomerase Reverse Transcriptase Gene. Genes 2016, 7, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070030
Zhang F, Cheng D, Wang S, Zhu J. Human Specific Regulation of the Telomerase Reverse Transcriptase Gene. Genes. 2016; 7(7):30. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070030
Chicago/Turabian StyleZhang, Fan, De Cheng, Shuwen Wang, and Jiyue Zhu. 2016. "Human Specific Regulation of the Telomerase Reverse Transcriptase Gene" Genes 7, no. 7: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7070030