
Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma

Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. DNA Isolation and Mutational Analysis
3. Results
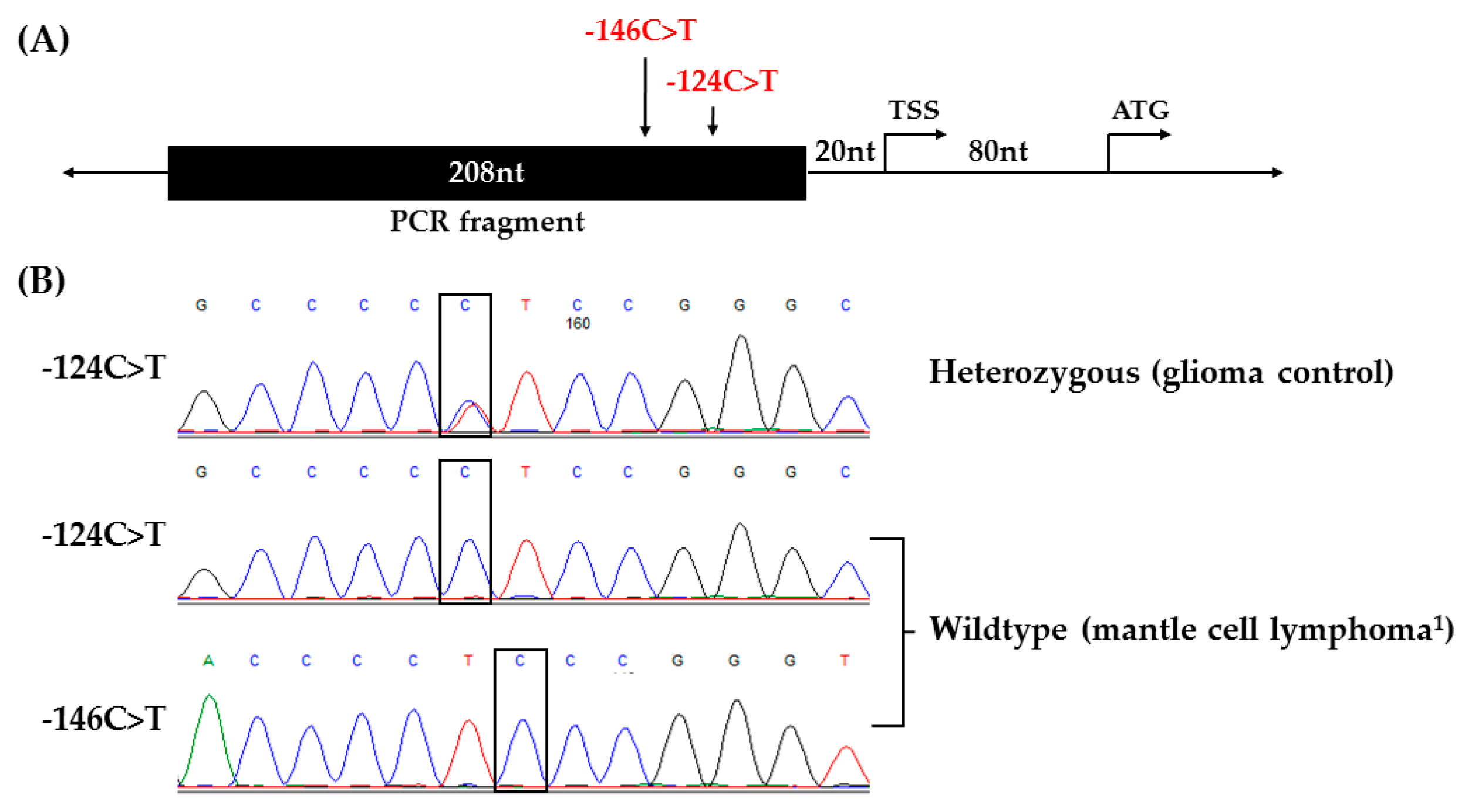
Absence of TERT Promoter Mutations in NHLs
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; IARC Press: Lyon, France, 2008. [Google Scholar]
- Biagi, J.; Seymour, J. Insights into the molecular pathogenesis of follicular lymphoma arising from analysis of geographic variation. Blood 2002, 99, 4265–4275. [Google Scholar] [CrossRef] [PubMed]
- Barrans, S.L.; Evans, P.A.; O’Connor, S.J.; Kendall, S.J.; Owen, R.G.; Haynes, A.P.; Morgan, G.J.; Jack, A.S. The t(14;18) is associated with germinal center-derived diffuse large B-cell lymphoma and is a strong predictor of outcome. Clin. Cancer Res. 2003, 9, 2133–2139. [Google Scholar] [PubMed]
- Bertoni, F.; Rinaldi, A.; Zucca, E.; Cavalli, F. Update on the molecular biology of mantle cell lymphoma. Hematol. Oncol. 2006, 24, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.G.; Stojanov, P.; Lawrence, M.S.; Auclair, D.; Chapuy, B.; Sougnez, C.; Cruz-Gordillo, P.; Knoechel, B.; Asmann, Y.W.; Slager, S.L.; et al. Discovery and prioritization of somatic mutations in diffuse large B-cell lymphoma (DLBCL) by whole-exome sequencing. Proc. Natl. Acad. Sci. USA 2011, 109, 3879–3884. [Google Scholar] [CrossRef] [PubMed]
- Okosun, J.; Bödör, C.; Wang, J.; Araf, S.; Yang, C.Y.; Pan, C.; Boller, S.; Cittaro, D.; Bozek, M.; Iqbal, S.; et al. Integrated genomic analysis identifies recurrent mutations and evolution patterns driving the initiation and progression of follicular lymphoma. Nat. Genet. 2014, 46, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Ansell, S.M. Non-Hodgkin lymphoma: Diagnosis and treatment. Mayo Clin. Proc. 2015, 90, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, S.; Bartsch, L.; Berkmann, J.C.; Fröse, K.; Heitmann, J.; Hoppe, C.; Iggena, D.; Jazmati, D.; Karschnia, P.; Linsenmeier, M.; et al. The dark matter of the cancer genome: Aberrations in regulatory elements, untranslated regions, splice sites, non-coding RNA and synonymous mutations. EMBO Mol. Med. 2016, 8, 442–457. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.J.; Rube, H.T.; Kreig, A.; Mancini, A.; Fouse, S.D.; Nagarajan, R.P.; Choi, S.; Hong, C.; He, D.; Pekmezci, M.; et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science 2015, 348, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, H.; Deb, S.; Liu, J. TERT regulates cell survival independent of telomerase enzymatic activity. Oncogene 2002, 21, 3130–3138. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hemminki, K.; Kumar, R. TERT promoter mutations in cancer development. Curr. Opin. Genet. Dev. 2014, 24, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef] [PubMed]
- Weinhold, N.; Jacobsen, A.; Schultz, N.; Sander, C.; Lee, W. Genome-wide analysis of noncoding regulatory mutations in cancer. Nat. Genet. 2014, 46, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Almeida, A.; Pópulo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wu, G.; Shan, Y.; Hartmann, C.; von Deimling, A.; Xing, M. Highly prevalent TERT promoter mutations in bladder cancer and glioblastoma. Cell Cycle 2013, 12, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, P.S.; Hosen, I.; de Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.J.; Rube, H.T.; Xavier-Magalhães, A.; Costa, B.M.; Mancini, A.; Song, J.S.; Costello, J.F. Understanding TERT promoter mutations: A common path to immortality. Mol. Cancer Res. 2016, 14, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xian, R.R.; Li, Y.; Polony, T.S.; Beemon, K.L. Telomerase reverse transcriptase expression elevated by avian leucosis virus integration in B cell lymphomas. Proc. Natl. Acad. Sci. USA 2007, 104, 18952–18957. [Google Scholar] [CrossRef] [PubMed]
- Justice, J.F., 4th; Morgan, R.W.; Beemon, K.L. Common viral integration sites identified in avian leukosis virus-induced B-cell lymphomas. Am. Soc. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.M. Telomeres and telomerase in leukaemia and lymphoma. Transfus. Apher. Sci. 2007, 37, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Lobetti-Bodoni, C.; Bernocco, E.; Genuardi, E.; Boccadoro, M.; Ladetto, M. Telomeres and telomerase in normal and malignant B-cells. Hematol. Oncol. 2010, 29, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Alentorn, A.; Daniau, M.; Labussière, M.; Rahimian, A.; Tabouret, E.; Polivka, M.; Jouvet, A.; Adam, C.; Figarella-Branger, D.; et al. TERT promoter mutations in primary central nervous system lymphoma are associated with spatial distribution in the splenium. Acta Neuropathol. 2015, 130, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Lan, Q.; Slager, S.L.; Vermeulen, R.C.; Teras, L.R.; Camp, N.J.; Cerhan, J.R.; Spinelli, J.J.; Wang, S.S.; Nieters, A.; et al. Genetically predicted longer telomere length is associated with increased risk of B-cell lymphoma subtypes. Hum. Mol. Genet. 2016, 25, 1663–1676. [Google Scholar] [CrossRef] [PubMed]
- Ohyashiki, J.; Sashida, G.; Tauchi, T.; Ohyashiki, K. Telomeres and telomerase in hematologic neoplasia. Oncogene 2002, 21, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Panero, J.; Alves-Paiva, R.M.; Roisman, A.; Santana-Lemos, B.A.; Falcão, R.P.; Oliveira, G.; Martins, D.; Stanganelli, C.; Slavutsky, I.; Calado, R.T. Acquired TERT promoter mutations stimulate TERT transcription in mantle cell lymphoma. Am. J. Hematol. 2016, 91, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Pittaluga, S.; Verhoef, G.; Criel, A.; Maes, A.; Nuyts, J.; Boogaerts, M.; De Wolf Peeters, C. Prognostic significance of bone marrow trephine and peripheral blood smears in 55 patients with mantle cell lymphoma. Leuk. Lymphoma. 1996, 21, 115–125. [Google Scholar] [CrossRef] [PubMed]
- De Unamuno Bustos, B.; Estal, R.M.; Simó, G.P.; Martínez, V.O.; Ros, M.L.; Suela, S.P.; Estrada, R.B. Lack of TERT promoter mutations in melanomas with extensive regression. J. Am. Acad. Dermatol. 2016, 74, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Melton, C.; Reuter, J.; Spacek, D.; Snyder, M. Recurrent somatic mutations in regulatory regions of human cancer genomes. Nat. Genet. 2015, 47, 710–716. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tumor Type | No. of Tumors | No. of Tumors Mutated |
|---|---|---|
| B-cell acute lymphoblastic leukemia | 12 | 0 |
| Burkitt lymphoma | 9 | 0 |
| Chronic lymphocytic leukemia | 11 | 0 |
| Diffuse large B-cell lymphoma | 9 | 0 |
| Follicular lymphoma | 13 | 0 |
| Lymphoplasmacytic lymphoma | 7 | 0 |
| Mantle cell lymphoma | 12 | 0 |
| Marginal zone lymphoma | 16 | 0 |
| Myeloma/plasmacytoma | 9 | 0 |
| Plasmablastic lymphoma | 7 | 0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, G.; Xian, R.R.; Li, Y.; Burns, K.H.; Beemon, K.L. Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma. Genes 2016, 7, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110093
Lam G, Xian RR, Li Y, Burns KH, Beemon KL. Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma. Genes. 2016; 7(11):93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110093
Chicago/Turabian StyleLam, Gary, Rena R. Xian, Yingying Li, Kathleen H. Burns, and Karen L. Beemon. 2016. "Lack of TERT Promoter Mutations in Human B-Cell Non-Hodgkin Lymphoma" Genes 7, no. 11: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7110093