
Et tu, Brute? Not Even Intracellular Mutualistic Symbionts Escape Horizontal Gene Transfer


1
Biologie Fonctionnelle Insectes et Interactions, UMR203 BF2I, INRA, INSA-Lyon, Université de Lyon, 69100 Villeurbanne, France
2
Institute for Integrative Systems Biology (I2SysBio), Universitat de València/CSIC, 46980 Paterna (València), Spain
3
Departament de Genètica, Universitat de València, Dr. Moliner, 50, 46100 Burjassot (València), Spain
*
Author to whom correspondence should be addressed.
Genes 2017, 8(10), 247; https://0-doi-org.brum.beds.ac.uk/10.3390/genes8100247
Submission received: 21 August 2017
/
Revised: 14 September 2017
/
Accepted: 22 September 2017
/
Published: 29 September 2017
(This article belongs to the Special Issue Horizontal Gene Transfer)
Abstract
:Many insect species maintain mutualistic relationships with endosymbiotic bacteria. In contrast to their free-living relatives, horizontal gene transfer (HGT) has traditionally been considered rare in long-term endosymbionts. Nevertheless, meta-omics exploration of certain symbiotic models has unveiled an increasing number of bacteria-bacteria and bacteria-host genetic transfers. The abundance and function of transferred loci suggest that HGT might play a major role in the evolution of the corresponding consortia, enhancing their adaptive value or buffering detrimental effects derived from the reductive evolution of endosymbionts’ genomes. Here, we comprehensively review the HGT cases recorded to date in insect-bacteria mutualistic consortia, and discuss their impact on the evolutionary success of these associations.
1. Introduction
1.1. Horizontal Gene Transfer: Molecular Signatures and Mechanisms
Vertical inheritance between generations, via sexual or asexual reproduction, represents the main mechanism for the transmission of genetic material in nature [1]. Nevertheless, genetic information can also be transmitted between reproductively isolated species via Horizontal Gene Transfer (HGT). This phenomenon is governed by three major mechanisms in prokaryotes: transformation (i.e., direct uptake of exogenous DNA), conjugation (i.e., plasmid-mediated uptake of exogenous DNA), and transduction (i.e., virus-mediated uptake of exogenous DNA). More recently, genetic transfer through prophages-derived gene transfer agents (GTAs) and cell fusion have been described [2]. On the other hand, the mechanisms allowing for HGT in eukaryotes remain unclear. Evidences suggest the key involvement of transposable elements [3,4,5,6,7], bacteriophages [8], giant viruses [9], and extracellular vesicles such as exosomes [10] in bacteria-to-animals transfer of genetic material.
Mutational change and subsequent selection might lead to the appearance of novel genes after the duplication of pre-existing loci. Alternatively, genes that have already undergone selective pressures can be directly transferred between different species via HGT [11]. Despite the evident evolutionary advantages of HGT (see Section 1.2), newly acquired loci often function inefficiently within their new genomic background [12,13] and/or generate detrimental side effects [14]. HGT-associated costs are related to several non-mutually exclusive phenomena. These include genetic/genomic features disruption [15,16]; sequence-specific signatures of the horizontally acquired loci [17]; sequestration of cell limited resources due to transcription and translation of gene products encoded by the foreign DNA [18]; cytotoxic effects due to misfolded proteins [19,20]; disruption of fine-tuning of cellular networks caused by changes in protein dosage [13,21] or inefficient interaction with local proteins due to the lack of molecular co-evolution [22,23]; and system-level effects derived from direct or indirect impact of acquired loci on the regulation of transcriptional patterns or the concentration of signaling metabolites [24,25].
Due to the HGT-associated costs, the kind of genes and pathways maintained over extended timescales after being transferred are highly biased. When present, horizontally acquired sequences are detected by using probabilistic methods, including molecular phylogenetics, codon usage, and oligonucleotide composition analyses [2]. In addition, synteny-based evidences or shared ecological niches between donor and recipient species can be used to further support the HGT-hypothesis.
1.2. Horizontal Gene Transfer as an Evolutionary Force
Horizontal transfer of genetic material plays a major evolutionary role among prokaryotes [26,27], explaining their extensive ecological diversification [28,29,30], and being relevant for bacterial evolution at least since the origins of the bacterial divisions [31]. Eukaryotes, however, present several barriers to HGT, including the selective double membrane of the cellular nucleus, the required adjustment of acquired genetic elements to the eukaryotic transcription machinery, and the need to affect the germ cell line in order to ensure intergenerational transmission in multicellular organisms [11,32]. In spite of this, prokaryotes-to-eukaryotes HGT events have drastically influenced eukaryotes early evolution. According to the Serial Endosymbiosis Theory (SET), free-living alphaproteobacteria and cyanobacteria were the ancestors of mitochondria and chloroplast, respectively. Their functional integration with early eukaryotes led to the organelles drastic genome reduction and horizontal transfer of both RNA- and protein-coding genes to the eukaryotic nuclear genome [1,33,34]. This phenomenon apparently keeps playing an important role in eukaryotes adaptive evolution [35]. Thus, many HGT events detected in fungi [36,37,38], plants [9,39], and animal genomes (see next sections for further details) involve bacteria as donor species. In general, bacteria possessing the ability to transfer DNA to eukaryotes closely interact with eukaryotic hosts (i.e., they maintain symbiotic associations of parasitic or mutualistic nature), and show high levels of genome plasticity and gene motility by means of a relevant mobilome. In addition, certain bacterial structures such as Type IV Secretion System (T4SS), the only natural bacterium-to-eukaryote DNA transfer system known so far [40], might facilitate this kind of transfer.
Next-generation sequencing technologies are yielding a growing body of evidence on HGT signatures in animal genomes [41]. Most HGT events affect invertebrates that display close associations with a broad range of microorganisms [42,43], and whose simpler structural organization is expected to increase the accessibility of their germline to exogenous DNA. In some cases, HGT drastically impacts animals’ biology. For instance, the human parasitic nematode Brugia malayi encodes an essential ferrochelatase gene of prokaryotic origin [44], and the transfer of a nearly complete Wolbachia pipientis genome has triggered the evolution of a new sex chromosome in pill bugs [45]. Furthermore, HGT have apparently allowed for the colonization of novel ecological niches. Thus, a number of genes encoding plant cell-wall degrading enzymes such as cellulases, xylanases, pectate lyases, and polygalacturonases have been found in plant–parasitic nematodes [4,46]. In the same line, the adaptation to the herbivorous lifestyle of the coffee berry borer beetle Hypothenemus hampei and the mustard leaf beetle Phaedon cochleariae has been facilitated by horizontally acquired genes (i.e., mannanase and xylanase, respectively), likely coming from the gut microbiota [3,6]. In some cases, selective advantages supplied by horizontally transferred loci might not be so obvious. For example, the genetic diversity of bdelloid rotifers provided by massive HGT from bacteria, fungi and plants might both compensate for their unisexual reproductive strategy and enable tolerance against desiccation [47].
2. Insects-Bacteria Endosymbioses
Nutritional symbioses between insects and intracellular bacteria are among the closest inter-domain associations found in nature. This kind of symbiosis mostly affects insects from orders Blattaria, Curculionidae and Hemiptera [48], being considered a key factor for their evolutionary success. Obligate mutualistic bacteria, also known as primary endosymbionts (P-endosymbionts), inhabit insect polyploid cells (bacteriocytes) which usually aggregate into a specialized bacteria-bearing organ (bacteriome; Figure 1). Many of them complement their host’s unbalanced diets (e.g., plant sap, vertebrates blood), allowing insects to colonize nutritionally poor niches. In coherence, a link between the diet composition and the nutritional role of the bacterial endosymbiont is usually observed, no matter the lifestyle or genetic repertoire of its free-living ancestor [49,50]. For example, phloem sap is enriched in carbohydrates but contains very small amounts of lipids, proteins or vitamins [51,52,53]. While most lipids can be synthesized from carbohydrates, proteins and vitamins cannot, due to the scarcity of nitrogenous precursors. Thus, provision of essential amino acids and vitamins is usually the nutritional role of P-endosymbionts in sap-feeding insects [54].
P-endosymbionts are essential for their host survival and reproduction, being fixed (i.e., 100% prevalence) in the host population [56]. Moreover, they are vertically transmitted from mothers to offspring [57], so that hosts and long-term P-endosymbionts show congruent phylogenies due to coevolution [58,59,60,61]. Both the intracellular environment they inhabit and the population dynamics associated to vertical inheritance drive the dramatic genome shrinkage, also known as genomic reduction syndrome, undergone by these bacteria during the transition to the intracellular lifestyle [43]. Bacteriocyte cytoplasm represents a rich, stable niche, rendering unnecessary or redundant many of the genes encoded by free-living bacteria genomes. Furthermore, periodical bottlenecking associated to the vertical transmission of P-endosymbionts between host generations increases the evolutionary impact of genetic drift [62,63], so that slightly deleterious mutations accumulate in loci under relaxed purifying selection, which then get inactivated and subsequently eliminated. Genes involved in DNA uptake, repair, and recombination are early affected by this genome-reduction process, usually leading to small, AT-enriched, structurally stable genomes showing few (if any) pseudogenes or mobile elements [57]. This is a progressive phenomenon yielding highly simplified genomes (i.e., the tiny genomes exhibited by symbionelles [64]). Highly reduced genomes lack essential functions either for the fulfillment of symbiosis requirements or for the maintenance of P-endosymbionts themselves [65,66,67,68]. In these cases, it has been hypothesized that the role of missing genes is taken over by the cooptation/functional plasticity of remaining loci [69], the recruitment of newly acquired symbionts (i.e., metabolic complementation and/or replacement by a co-primary endosymbiont; [70,71]), or the participation of the host via eukaryotic or horizontally-acquired prokaryotic genes [72].
In addition to nutritional symbioses, transient associations between insects and facultative/secondary symbionts (S-symbionts) have been described. In contrast to obligatory endosymbionts, S-symbionts display only partial infection of host populations. Moreover, they can be laterally transferred between host species [73,74], and are found within cell types other than bacteriocytes, as well as free in the host haemolymph [75,76,77]. Some of them, collectively known as reproductive parasites, are able to distort their host’s sex ratio by inducing reproductive alterations such as cytoplasmic incompatibility, parthenogenesis induction in haplo-diploid species, feminization of genetic males, or male-killing [78,79]. Other S-symbionts are known to enrich the genotype of insects by providing them with adaptive ecological traits. For instance, some S-symbionts broad the food plant range of their host or enhance their resistance to both biological and/or physical environmental stress [80].
3. Horizontal Gene Transfer in Insects-Bacteria Endosymbiotic Systems
3.1. Bacteria to Insects and Vice Versa
Wolbachia is among the most widespread intracellular bacteria described [79]. A variety of HGT events between Wolbachia and many of its insect hosts have been noticed, in coherence with its high estimated prevalence (from 20% to 66% insect species; [81,82]), and its close association with insects’ germ line [79]. These include both the Wolbachia-to-host transfer of genetic material [8,83,84] and vice versa [85]. Nevertheless, the evolutionary significance of these HGT events is hard to demonstrate [86], since most horizontally acquired genes are transcriptionally inactive and/or exhibit premature stop codons, frameshifts, or retroelement insertions [87].
In contrast to this general view, HGT is considered one of the signatures of genome coevolution in insect-bacteria nutritional endosymbioses [72]. As demonstrated by the recent accumulation of hologenomes (i.e., coupling of both endosymbiont and host genomic information), the horizontal acquisition of prokaryotic genes by insects maintaining nutritional symbiosis with intracellular bacteria has allowed for further integration of the corresponding consortia (Figure 2). Examples have been recently described in the pea aphid Acyrthosiphon pisum, the citrus mealybug Planococcus citri, the hackberry petiole gall psyllid Pachypsylla venusta, and the cotton whitefly Bemisia tabaci, which are representatives of all insect families within the suborder Sternorrhyncha [88,89,90,91,92]. Despite considerable overlap in the involved functions, most of the homologous loci acquired by mealybugs, psyllids and whiteflies clearly derive from independent transfer events, since they are not phylogenetically related (Table 1). Strikingly, most of the acquired loci appear to be transferred from transient S-symbionts and/or former, already replaced, P-endosymbionts. Taking into account the fixation of P-endosymbionts in the host populations, as well as their vertical transmission via eggs colonization, this probably indicates that (1) bacteria-to-insect HGT are rare molecular events; and/or (2) P-endosymbionts are bad candidates for successful DNA emission, likely because of the mutational bias and the low protein stability derived from their genomic reductive syndrome [93,94].
Horizontally acquired genes mediating insect–bacteria nutritional symbiosis were first described in the pea aphid A. pisum (Hemiptera: Aphididae), which maintains a long-term mutualistic association with the gammaproteobacterial endosymbiont Buchnera aphidicola [95]. A total of 12 prokaryotic genes or gene fragments have been found in the A. pisum genome, only two of them (the pseudogenes ψdnaE and ψatpH) being likely acquired from its P-endosymbiont B. aphidicola [88,89]. The other 10 genes of prokaryotic origin present in the host’s nuclear genome encode enzymes involved in peptidoglycan metabolism: several LD-carboxypeptidases (LdcA, LdcA1 and LdcA2; EC:3.4.17.13) and an N-acetylmuramoyl-l-alanine amidase (AmiD, EC:3.5.1.28), required for murein recycling [96], as well as lysozyme (bLys, EC:3.2.1.17), which hydrolyzes it [97]. In contrast to LdcA1 and AmiD, the expression of bLys was more abundant in other cell types than in the bacteriocyte, suggesting that it might be involved both in the regulation of symbiosis and the protection of A. pisum against the infection by exogenous bacteria [97,98]. Phylogenetic analyses showed that LdcAs, AmiD, and bLys have been horizontally acquired from rickettsial bacteria, which include many lineages (e.g., Rickettsia, Wolbachia) commonly found as S-symbionts in aphids [99,100]. In addition, five copies of the gene encoding RlpA (rare lipoprotein A) were detected. The bacterial source and function of this gene is still unclear. Nevertheless, protein localization of RlpA4 showed that it is targeted to B. aphidicola cells, which demonstrate that RlpA is both functional and tightly involved in maintenance/control of the nutritional symbiosis between A. pisum and its P-endosymbiont [101].
Mealybugs (Hemiptera: Pseudococcidae) maintain a variety of nutritional symbioses involving a betaproteobacterium of the genus “Candidatus Tremblaya” (except if replaced by a newly acquired endosymbiont [102]). While “Candidatus Tremblaya phenacola” remains alone in the bacteriocytes of phenacoccinae mealybugs, “Candidatus Tremblaya princeps” have independently engulfed several lineages of gammaproteobacteria that inhabit “Ca. Tremblaya princeps” cytoplasm as co-primary endosymbionts [103,104,105]. These nutritional symbioses are based on the biosynthesis of essential amino acids and vitamins, as first demonstrated by the genomic characterization of the tripartite consortium involving the citrus mealybug P. citri and its two co-primary endosymbionts “Ca. Tremblaya princeps” and “Candidatus Moranella endobia” [66,67]. Further exploration of the P. citri’s nuclear genome revealed the presence of 22 functional prokaryotic genes coming from Alphaproteobacteria, Gammaproteobacteria and Bacteroidetes, closely related to many of the most frequent insects S-symbionts (Arsenophonus, Cardinium, Serratia, Sodalis, Rickettsia and Wolbachia) [90]. Three of them (dapF, lysA and cysK) apparently fulfill missing gaps in the bacterial lysine and methionine biosynthetic pathways. Another five appear to complement “Ca. Moranella endobia” incomplete pathways for the biosynthesis of riboflavin (ribA and ribD) and biotin (bioA, bioB and bioD). As previously noticed in A. pisum, bacterial genes involved in peptidoglycan metabolism have also been acquired by P. citri. These include loci devoted to peptidoglycan biosynthesis (murA, murB, murC, murD, murE, murF and ddlB) and recycling (mltB and amiD). It has been proposed that these genes might mediate the molecular crosstalk between “Ca. Tremblaya princeps” and its nested endosymbiont “Ca. Moranella endobia“ [90,106].
Ten bacterial genes have been identified in the nuclear genome of P. venusta, most of them being also present in the nuclear genome of the Asian citrus psyllid Diaphorina citri and the potato psyllid Bactericera cockerelli [91]. As expected for psyllids (Hemiptera: Sternorrhyncha), P. venusta maintains a long-term nutritional symbiosis with the gammaproteobacterium “Candidatus Carsonella ruddii” [107]. In line with the findings in P. citri, some of the horizontally acquired genes are involved in the biosynthesis of arginine, phenylalanine and riboflavin. While the gene coding for chorismate mutase (CM, EC:5.4.99.5) appears to compensate for “Ca. Carsonella ruddi” functional simplification during genome shrinkage, the nuclear gene of prokaryotic origin ribC (encoding riboflavin synthase, EC:2.5.1.9) is the single locus devoted to riboflavin biosynthesis in the holobiont genome. Authors suggest that ribC might represent a vestige of an ancient complementarity for riboflavin biosynthesis with a former, already lost endosymbiont in the Pachyspylla lineage. In coherence, this gene is present in the genome of D. citri, which still requires the bacterial supply of riboflavin, according to the functional analysis of its endosymbiotic consortium [108]. On the other hand, “Ca. Carsonella ruddii” still retains a functional copy of argH (encoding argininosuccinate lyase, EC:4.3.2.1) that is closely related to the two copies of such gene found in the host genome, suggesting that this P-endosymbiont was the bacterial donor. For the rest of horizontally acquired loci, frequent S-symbiotic bacteria such as Rickettsia and Wolbachia might be the source [91].
Finally, horizontally acquired genes were also found in the nuclear genome of the whitefly Bemisia tabaci (Hemiptera: Aleyrodoidea), which maintains a long-term obligatory symbiosis with the gammaproteobacterium “Ca. Portiera aleyrodidarum” [60]. In Portiera-BT genomes, many genes involved in the biosynthesis of essential amino acids have been inactivated (argH, dapB) or lost (argA, argB, argC, argE, dapF, lysA, hisD), affecting its capacity to supply arginine, lysine and histidine to its host [109,110,111]. Furthermore, “Ca. Portiera aleyrodidarum” is unable to synthesize a set of cofactors (i.e., thiamine, nicotinamide, pyridoxal-5-phosphate, folic acid, FMN/FADH, ubiquinone, heme) predicted to be required for its metabolic functions. Part of these deficiencies can be compensated by a second endosymbiont, the gammaproteobacterium “Candidatus Hamiltonella defensa”, which retains dapB, dapF and lysA (lysine biosynthesis) and encodes the complete biosynthetic pathways for six of the eight required cofactors. Nevertheless, it still lacks the complete thiamine biosynthetic pathway as well as phoAB, involved in folate biosynthesis. Their host, B. tabaci, encodes ten metabolic genes of bacterial origin [92]. Six of them are involved in the biosynthesis of the essential amino acids arginine (argG and argH), lysine (dapB, dapF and lysA), and phenylalanine (CM). The rest of the horizontally acquired genes code for proteins involved in the biosynthesis of thiamine (bioA and bioB) and the degradation of urea (DUR1,2 and AH, encoding urea carboxylase/allophanate hydrolase, EC:6.3.4.6/EC:3.5.1.54, and allophanate hydrolase, EC:3.5.1.54, respectively). The acquisition of CM, bioA and bioB, at least, is relatively ancient, since the corresponding transcripts were detected also in the greenhouse whitefly Trialeurodes vaporariorum. Even if “Ca. Portiera aleyrodidarum” and/or “Ca. Hamiltonella defensa” still retain homologs for all horizontally acquired loci but AH and DUR1,2, phylogenetic evidences rule out the possibility that they were the bacterial sources. In contrast, transferred genes cluster with different Gammaproteobacteria (including Pantoea and stinkbugs gut symbionts), Alphaproteobacteria (Rickettsiales), Bacteroidetes (Niastella) and Planctomycetes (Isosphaera).
3.2. Bacteria to Bacteria
As mentioned above, prokaryotes are highly prone to DNA exchange. The genomes of many insect endosymbiotic bacteria have been characterized in the last 17 years, but very few evidences for bacteria-to-bacteria gene transfer have been noticed, even if the transient infection by S-symbionts and/or the stable presence of multiple co-primary endosymbiotic species are frequent.
3.2.1. Reasons behind Horizontal Gene Transfer Scarcity among Endosymbionts
The intracellular lifestyle, the vertical transmission and genomic features typically associated with their genome reduction syndrome (i.e., AT-enrichment and gene repertoire simplification) might explain the refractoriness of endosymbionts to HGT.
• Intracellular lifestyle: HGT-associated costs limit the divergence between receptor genomes and successfully transferred loci (see Section 1.1). Therefore, HGT incidence between closely related bacteria is significantly more frequent [112]. Closely related free-living bacteria get often in contact because they are likely to share the same habitat [113]. In contrast, endosymbionts are isolated from their ancestral habitat and close relatives when adapting to the intracellular lifestyle. Moreover, bacteriocytes isolates them from any environmental source of prokaryotic DNA. In fact, even if insects can frequently be co-infected with multiple bacterial lineages, these bacteria do not necessarily co-exist within the same bacteriocyte [99]. On the other hand, because of their intracellular distribution, endosymbiotic populations effective sizes are dramatically smaller than those of free-living bacteria, which is expected to hamper genome fluidity [114].
• Vertical transmission: Endosymbionts effective population size is greatly affected by serial bottlenecking during maternal transmission to the host’s next generation, which is expected to enhance genetic drift effects (i.e., elimination of at least part of the genomic polymorphism accumulating in the population [115]). This might hinder the inheritance of horizontally acquired functions, as well as the natural selection-driven amelioration of harmful pleiotropic effects associated with HGT [116,117].
• Genome reductive evolution: The dramatic gene repertoire simplification typically observed among long-term endosymbionts (e.g., scarcity of bacteriophages and plasmids; loss of mobile elements, T4SS, and homologous recombination genes) is expected to hamper the gene flow among endosymbionts and/or the insertion of exogenous DNA into their own replicons. In the same line, the AT-enrichment of their genomes might hinder the incidence of successful HGT events in several ways. First, gene transfer between bacterial lineages exhibiting highly different nucleotide content is unlikely, since integration of exogenous DNA via homologous recombination requires a ‘minimum efficient processing segment’ (MEPS) consisting of near-identical sequences of at least 25 bp at one or both ends of a donor segment [118,119,120]. Moreover, skewed nucleotide content leads to changes in codon usage (or even in the genetic code) [68,121] and protein amino acids composition [122]. Codon content is linked to gene expression by modulating translation efficiency and mRNA stability [123], while improper codon usage might favor a resources sequestration effect by stalling ribosomes during translation [17]. In addition, proteins containing rare amino acids are known to entail higher fitness costs than those using abundant ones [124].
3.2.2. From Genetic Transfer to Genomic Fusion
In spite of evident barriers for exogenous DNA acquisition, HGT events have been noticed in several bacterial endosymbionts. For instance, genes for nitrogen fixation were horizontally acquired by “Candidatus Thiodiazotropha endoloripes”, endosymbiont of the lucinid bivalve Loripes lucinalis [125]. In the same line, “Candidatus Endomicrobium trichonymphae” strain Rs-D17, endosymbiont of the termite gut flagellate Trichonympha agilis, might have acquired loci encoding a bifunctional aldehyde dehydrogenase/ethanol dehydrogenase, amino acids transporters, and proteins involved in the biosynthesis of thiamine pyrophosphate [126]. Because of their distribution within the host (“Ca. Thiodiazotropha endoloripes”-containing bacteriocytes are located along the L. lucinalis gill lamellae, being highly exposed to the seawater flow), the lower structural complexity of hosts and/or the abundance of DNA sources in the niche they colonize (T. agilis inhabits the microbial-enriched gut of termites [127]), these bacteria seem relatively more exposed to exogenous DNA than insect-associated nutritional endosymbionts. In any case, HGT events accounting for a variable number of loci have been also noticed in bacteriome-associated endosymbionts of insects, confirming that HGT is a key source of evolutionary novelties throughout the prokaryotes. The examples described so far include Wolbachia pipientis str. wCle, “Candidatus Profftella armature” and “Ca. Tremblaya phenacola” (Table 2).
Wobachia wCle is associated with the bedbug Cimex lectularius, being essential for insect growth and reproduction via provisioning of biotin and riboflavin [128,130]. Genomic characterization of this Wolbachia strain revealed that, unlike all available insect-associated facultative Wolbachia, it encodes a complete operon for the biosynthesis of biotin (i.e., genes bioA, bioB, bioC, bioD, bioF and bioH), as well as a partial operon for the biosynthesis of thiamine (genes tenA1 and thiD, and the pseudogene ψthiM). A similar operon structure for biotin biosynthesis has been identified in Cardinium hertigii cEper1, S-symbiont of the parasitoid wasp Encarsia pergandiella [131], the swine pathogen Lawsonia intracellularis [132], and a Rickettsia strain isolated from the tick Ixodes scapularis [133]. On the other hand, similar operon configuration for thiamine biosynthesis was identified in the fish pathogen Francisella noatunensis [134]. Based on this information, as well as on phylogenetic analyses, authors suggest that biotin-biosynthetic genes were probably horizontally acquired as a whole operon by an ancestor of Wolbachia wCle, either from Cardinium or Rickettsia, which frequently coinfect the same insect host [78]. Similarly, phylogenetic analyses showed that thiamine-biosynthetic loci were closely related to the homologous genes found in F. noatunensis, Brachyspira hyodysenteriae, Legionella drancourtii, and additional bacterial lineages, including representatives of the Gammaproteobacteria, Bacteroidetes, Spirochaetes, etc., supporting the HGT hypothesis. Moreover, nutritional and physiological experiments allowed them to postulate the horizontal acquisition of genes for biotin biosynthesis as a key step towards the transition between the facultative and the obligatory association of Wolbachia with its host [128].
Up to 20 genes have been horizontally acquired by “Ca. Profftella armature”, the co-primary endosymbiont of the psyllid D. citri, along with “Ca. Carsonella ruddi” str. DC [108]. Acquired genes, organized into two polyketide synthase biosynthetic gene clusters, synthesize a polyketide toxin named diaphorin. It is an analog of pederin, a defensive polyketide produced by a Pseudomonas symbiont of Paederus rove beetles, allowing the insect host to deter predators [135,136]. Authors suggest that prey-predator relationship might be involved in this HGT event, since Paederus rove beetles often feed on hemipteran insects [137].
In contrast to the above described examples, the case of the betaproteobacterium “Ca. Tremblaya phenacola” PPER, single P-endosymbiont of the bougainvillea mealybug P. peruvianus, goes far beyond conventional HGT, rather suggesting the formation of a new chimeric organism after the fusion of two complete genomes. Recent genome sequencing and analysis of “Ca. Tremblaya phenacola” PPER (Figure 1, [55]) revealed the presence of at least 80 gammaproteobacterial genes still showing a characteristic molecular signature in terms of GC-content and codon usage [129]. The functional distribution of genes is not random. According to the complexity hypothesis, genes coding for functions burdened with many complex interactions might display low HGT rates, since partial transfer of co-adapted structures is likely to end up in loss of function due to incompatibility [138,139]. Coherently, the components of complex molecular machineries share a common evolutionary origin in “Ca. Tremblaya phenacola” PPER [129]. Similarly to previously described symbiotic systems in mealybugs [66,67,90,105,140], bacterial supply of essential amino acids appears to be the basis of the nutritional association between “Ca. Tremblaya phenacola” PPER and P. peruvianus. Regarding this role, betaproteobacterial genes carry out the biosynthesis of methionine, threonine, isoleucine, leucine and valine, as well as the production of phenylalanine from chorismate. Only gammmaproteobacterial genes have been retained for the biosynthesis of histidine and cysteine, while both beta- and gammaproteobacterial genes collaborate in the biosynthesis of tryptophan. Overall, gammaproteobacterial genes represent 46% of PPER’s genome and place it into the Sodalis-allied clade. In contrast, its betaproteobacterial genes (including a single ribosomal operon) clearly place it within the “Ca. Tremblaya phenacola” clade. Thus, similarly to free-living bacteria undergoing extended HGT events, PPER is a nightmare for molecular taxonomists [141]. Taking into account the diverse organization of nutritional symbioses among mealybugs [102,103], a gammaproteobacterium apparently entered the symbiotic consortium in the lineage leading to “Ca. Tremblaya phenacola” PPER. Then, instead of replacing “Ca. Tremblaya” [102], genomic fusion and subsequent gene shuffling took place, likely involving homologous recombination genes encoded by the gammaproteobacterial donor [142,143]. Both the scarcity of functional redundancies and the taxonomic assignation of genes involved in the tryptophan biosynthetic pathway suggest that “Ca. Tremblaya phenacola” PPER coevolved with the gammaproteobacterial donor within P. peruvianus bacteriocytes for some time [129]. Whether a pseudococcinae-like nested consortium preceded the genomic fusion remains unclear.
4. Conclusions
Close association with microorganisms allowed animals to colonize highly specialized niches. This is the case of insects, whose facultative/obligatory association with mutualistic intracellular bacteria is considered essential for their evolutionary success. The ever-increasing availability of genomic data has highlighted the high impact of inter-domain associations on the horizontal acquisition of exogenous DNA by insects. Although most of the cases appear to represent transient transfers of genes, lacking an effective integration in the biology of the recipient species, recent findings strongly suggest that HGT might play a key role in the fine-tuning of mechanisms allowing for the maintenance and regulation of insect-bacteria nutritional symbioses. In the holobiont era, further analyses of available genomic/transcriptomic data, exploration of additional symbiotic models, and empirical assessment of the adaptive value of transferred loci are expected to enhance this new paradigm.
Acknowledgments
Financial support was provided by grants BFU2015-64322-C2-1-R (co-financed by FEDER funds and Ministerio de Economía y Competitividad, Spain) and PrometeoII/2014/065 (Conselleria d’Educació, Generalitat Valenciana, Spain).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ku, C.; Nelson-Sathi, S.; Roettger, M.; Sousa, F.L.; Lockhart, P.J.; Bryant, D.; Hazkani-Covo, E.; McInerney, J.O.; Landan, G.; Martin, W.F. Endosymbiotic origin and differential loss of eukaryotic genes. Nature 2015, 524, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Acuña, R.; Padilla, B.E.; Flórez-Ramos, C.P.; Rubio, J.D.; Herrera, J.C.; Benavides, P.; Lee, S.J.; Yeats, T.H.; Egan, A.N.; Doyle, J.J.; et al. Adaptive horizontal transfer of a bacterial gene to an invasive insect pest of coffee. Proc. Natl. Acad. Sci. USA 2012, 109, 4197–4202. [Google Scholar] [CrossRef] [PubMed]
- Paganini, J.; Campan-Fournier, A.; Da Rocha, M.; Gouret, P.; Pontarotti, P.; Wajnberg, E.; Abad, P.; Danchin, E.G. Contribution of lateral gene transfers to the genome composition and parasitic ability of root-knot nematodes. PLoS ONE 2012, 7, e50875. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.-F.; Hespeels, B.; Li, X.; Noel, B.; Arkhipova, I.; Danchin, E.G.J.; Hejnol, A.; Henrissat, B.; Koszul, R.; Aury, J.M.; et al. Genomic evidence for ameiotic evolution in the bdelloid rotifer Adineta vaga. Nature 2013, 500, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Pauchet, Y.; Heckel, D.G. The genome of the mustard leaf beetle encodes two active xylanases originally acquired from bacteria through horizontal gene transfer. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Boulain, H.; Gauthier, J.; Hua-Van, A.; Musset, K.; Jakubowska, A.K.; Aury, J.M.; Volkoff, A.N.; Huguet, E.; Herrero, S.; et al. Recurrent domestication by Lepidoptera of genes from their parasites mediated by bracoviruses. PLoS Genet. 2015, 11, e1005470. [Google Scholar] [CrossRef] [PubMed]
- Klasson, L.; Kambris, Z.; Cook, P.E.; Walker, T.; Sinkins, S.P. Horizontal gene transfer between Wolbachia and the mosquito Aedes aegypti. BMC Genom. 2009, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Schönknecht, G.; Chen, W.-H.; Ternes, C.M.; Barbier, G.G.; Shrestha, R.P.; Stanke, M.; Bräutigam, A.; Baker, B.J.; Banfield, J.F.; Garavito, R.M.; et al. Gene transfer from bacteria and archaea facilitated evolution of an extremophilic eukaryote. Science 2013, 339, 1207–1210. [Google Scholar] [CrossRef]
- Mashburn-Warren, L.M.; Whiteley, M. Special delivery: Vesicle trafficking in prokaryotes. Mol. Microbiol. 2006, 61, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, B.; Citovsky, V. Transfer of DNA from bacteria to eukaryotes. Am. Soc. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.-H.; Chiu, H.-C.; Delaney, N.F.; Segre, D.; Marx, C.J. Diminishing returns epistasis among beneficial mutations decelerates adaptation. Science 2011, 332, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Zhang, J. High expression hampers horizontal gene transfer. Genome Biol. Evol. 2012, 4, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Engelberg-Kulka, H.; Glaser, G. Addiction modules and programmed cell death and antideath in bacterial cultures. Ann. Rev. Microbiol. 1999, 53, 43–70. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F. Transposon-based strategies for microbial functional genomics and proteomics. Ann. Rev. Genet. 2003, 37, 3–29. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, L.; Sigal, N.; Borovok, I.; Nir-Paz, R.; Herskovits, A.A. Prophage excision activates Listeria competence genes that promote phagosomal escape and virulence. Cell 2012, 150, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Gingold, H.; Pilpel, Y. Determinants of translation efficiency and accuracy. Mol. Syst. Biol. 2011, 7, 481. [Google Scholar] [CrossRef] [PubMed]
- Bragg, J.G.; Wagner, A. Protein material costs: Single atoms can make an evolutionary difference. Trends Genet. 2009, 25, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Narra, H.P.; Cordes, M.H.J.; Ochman, H. Structural features and the persistence of acquired proteins. Proteomics 2008, 8, 4772–4781. [Google Scholar] [CrossRef] [PubMed]
- Geiler-Samerotte, K.A.; Dion, M.F.; Budnik, B.A.; Wang, S.M.; Hartl, D.L.; Drummond, D.A. Misfolded proteins impose a dosage-dependent fitness cost and trigger a cytosolic unfolded protein response in yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Gout, J.-F.; Kahn, D.; Duret, L. Paramecium post-genomics consortium. The relationship among gene expression, the evolution of gene dosage, and the rate of protein evolution. PLoS Genet. 2010, 6, e1000944. [Google Scholar] [CrossRef]
- Pál, C.; Papp, B.; Lercher, M.J. Horizontal gene transfer depends on gene content of the host. Bioinformatics 2005, 21. [Google Scholar] [CrossRef] [PubMed]
- Wellner, A.; Gophna, U. Neutrality of foreign complex subunits in an experimental model of lateral gene transfer. Mol. Biol. Evol. 2008, 25, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Golding, I.; Sawai, S.; Guo, L.; Cox, E.C. Population fitness and the regulation of Escherichia coli genes by bacterial viruses. PLoS Biol. 2005, 3, e229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, G.A.L.; Bower, D.M.; Prazeres, D.M.F.; Monteiro, G.A.; Prather, K.L.J. Rational engineering of Escherichia coli strains for plasmid biopharmaceutical manufacturing. Biotechnol. J. 2012, 7, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal gene transfer in prokaryotes: Quantification and classification. Ann. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef] [PubMed]
- Beiko, R.G.; Harlow, T.J.; Ragan, M.A. Highways of gene sharing in prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 14332–14337. [Google Scholar] [CrossRef] [PubMed]
- Martiny, A.C.; Huang, Y.; Li, W. Occurrence of phosphate acquisition genes in Prochlorococcus cells from different ocean regions. Environ. Microbiol. 2009, 11, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Monchy, S.; Taghavi, S.; Zhu, W.; Ramos, J.; van der Lelie, D. Comparative genomics and functional analysis of niche-specific adaptation in Pseudomonas putida. FEMS Microbiol. 2011, 35, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Walk, S.T.; Gordon, D.M.; Feldgarden, M.; Tiedje, J.M.; Konstantinidis, K.T. Genome sequencing of environmental Escherichia coli expands understanding of the ecology and speciation of the model bacterial species. Proc. Natl. Acad. Sci. USA 2011, 108, 7200–7205. [Google Scholar] [CrossRef] [PubMed]
- Gogarten, J.P.; Doolittle, W.F.; Lawrence, J.G. Prokaryotic evolution in light of gene transfer. Mol. Biol. Evol. 2002, 19, 2226–2238. [Google Scholar] [CrossRef] [PubMed]
- Wybouw, N.; Pauchet, Y.; Heckel, D.G.; Van Leeuwen, T. Horizontal gene transfer contributes to the evolution of arthropod herbivory. Genome Biol. Evol. 2016, 8, 1785–1801. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Archibald, J.M. Endosymbiosis and eukaryotic cell evolution. Curr. Biol. 2015, 25, R911–921. [Google Scholar] [CrossRef] [PubMed]
- Schönknecht, G.; Weber, A.P.M.; Lercher, M.J. Horizontal gene acquisitions by eukaryotes as drivers of adaptive evolution. BioEssays 2014, 36, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.; Brachat, S.; Dietrich, F.S. Contribution of horizontal gene transfer to the evolution of Saccharomyces cerevisiae. Eukaryot. Cell 2005, 4, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Klosterman, S.J.; Subbarao, K.V.; Kang, S.; Veronese, P.; Gold, S.E.; Thomma, B.P.H.J.; Chen, Z.; Henrissat, B.; Lee, Y.H.; Park, J.; et al. Comparative genomics yields insights into niche adaptation of plant vascular wilt pathogens. PLoS Pathog. 2011, 7, e1002137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo, V.D.A.; Sukno, S.A.; Thon, M.R. Identification of horizontally transferred genes in the genus Colletotrichum reveals a steady tempo of bacterial to fungal gene transfer. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Hu, X.; Sun, H.; Yang, Y.; Huang, J. Widespread impact of horizontal gene transfer on plant colonization of land. Nat. Commun. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Christie, P.J.; Whitaker, N.; González-Rivera, C. Mechanism and structure of the bacterial type IV secretion systems. Biochim. Biophys. Acta 2014, 1843, 1578–1591. [Google Scholar] [CrossRef] [PubMed]
- Boto, L. Horizontal gene transfer in the acquisition of novel traits by metazoans. Proc. Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Dubilier, N.; Bergin, C.; Lott, C. Symbiotic diversity in marine animals: The art of harnessing chemosynthesis. Nat. Rev. Microbiol. 2008, 6, 725–740. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Peretó, J.; Gil, R.; Latorre, A. Learning how to live together: Genomic insights into prokaryote-animal symbioses. Nat. Rev. Genet. 2008, 9, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Novelli, J.; Jiang, D.; Dailey, H.A.; Landmann, F.; Ford, L.; Taylor, M.J.; Carlow, C.K.; Kumar, S.; Foster, J.M.; et al. Interdomain lateral gene transfer of an essential ferrochelatase gene in human parasitic nematodes. Proc. Natl. Acad. Sci. USA 2013, 110, 7748–7753. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, S.; Thézé, J.; Chebbi, M.A.; Giraud, I.; Moumen, B.; Ernenwein, L.; Grève, P.; Gilbert, C.; Cordaux, R. Birth of a W sex chromosome by horizontal transfer of Wolbachia bacterial symbiont genome. Proc. Natl. Acad. Sci. USA 2016, 113, 15036–15041. [Google Scholar] [CrossRef] [PubMed]
- Danchin, E.G.J.; Rosso, M.-N.; Vieira, P.; de Almeida-Engler, J.; Coutinho, P.M.; Henrissat, B.; Abad, P. Multiple lateral gene transfers and duplications have promoted plant parasitism ability in nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 17651–17656. [Google Scholar] [CrossRef] [PubMed]
- Boschetti, C.; Pouchkina-Stantcheva, N.; Hoffmann, P.; Tunnacliffe, A. Foreign genes and novel hydrophilic protein genes participate in the desiccation response of the bdelloid rotifer Adineta ricciae. J. Exp. Biol. 2011, 214, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H. Insect symbiosis: An Introduction. In Insect Symbiosis, 1st ed.; Bourtzis, K., Miller, T.A., Eds.; CRC Press: Boca Raton, FL, USA, 2003; Volume 1, pp. 1–16. ISBN 9780849312861. [Google Scholar]
- McCutcheon, J.P.; McDonald, B.R.; Moran, N.A. Convergent evolution of metabolic roles in bacterial co-symbionts of insects. Proc. Natl. Acad. Sci. USA 2009, 106, 15394–15399. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.P.; Moran, N.A. Functional convergence in reduced genomes of bacterial symbionts spanning 200 My of evolution. Genome Biol. Evol. 2010, 2, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. The nutritional quality of phloem sap utilized by natural aphid populations. Ecol. Entomol. 1993, 18, 31–38. [Google Scholar] [CrossRef]
- Sandström, J.; Pettersson, J. Amino acid composition of phloem sap and the relation to intraspecific variation in pea aphid (Acyrthosiphon pisum) performance. J. Insect Physiol. 1994, 40, 947–955. [Google Scholar] [CrossRef]
- Dinant, S.; Bonnemain, J.-L.; Girousse, C.; Kehr, J. Phloem sap intricacy and interplay with aphid feeding. C. R. Biol. 2010, 333, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosser, W.A.; Douglas, A.E. A test of the hypotheses that nitrogen is upgraded and recycled in an aphid (Acyrthosiphon pisum) symbiosis. J. Insect Physiol. 1992, 38, 93–99. [Google Scholar] [CrossRef]
- López-Madrigal, S.; Beltrà, A.; Resurrección, S.; Soto, A.; Latorre, A.; Moya, A.; Gil, R. Molecular evidence for ongoing complementarity and horizontal gene transfer in endosymbiotic systems of mealybugs. Front. Microbiol. 2014, 5, 449. [Google Scholar] [CrossRef] [PubMed]
- Houk, E.J.; Griffiths, G.W. Intracellular symbiotes of the Homoptera. Ann. Rev. Entomol. 1980, 25, 161–187. [Google Scholar] [CrossRef]
- Baumann, P. Biology bacteriocyte-associated endosymbionts of plant sap-sucking insects. Ann. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef] [PubMed]
- Munson, M.A.; Baumann, P.; Moran, N.A. Phylogenetic relationships of the endosymbionts of mealybugs (Homoptera: Pseudococcidae) based on 16S rDNA sequences. Mol. Phylogenet. Evol. 1992, 1, 26–30. [Google Scholar] [CrossRef]
- Sauer, C.; Stackebrandt, E.; Gadau, J.; Hölldobler, B.; Gross, R. Systematic relationships and cospeciation of bacterial endosymbionts and their carpenter ant host species: Proposal of the new taxon Candidatus Blochmannia gen. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Thao, M.L.; Baumann, P. Evolutionary relationships of primary prokaryotic endosymbionts of whiteflies and their hosts. Appl. Environ. Microbiol. 2004, 70, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Rosenblueth, M.; Sayavedra, L.; Sámano-Sánchez, H.; Roth, A.; Martínez-Romero, E. Evolutionary relationships of flavobacterial and enterobacterial endosymbionts with their scale insect hosts (Hemiptera: Coccoidea). J. Evol. Biol. 2012, 25, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A. Accelerated evolution and Muller’s rachet in endosymbiotic bacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 2873–2878. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict host-symbiont cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biol. 2006, 4, e337. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Prieto, M.; Latorre, A.; Moya, A. Scanty microbes, the “symbionelle” concept. Environ. Microbiol. 2014, 16, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Nakabachi, A.; Yamashita, A.; Toh, H.; Ishikawa, H.; Dunbar, H.E.; Moran, N.A.; Hattori, M. The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 2006, 314, 267. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.P.; von Dohlen, C.D. An interdependent metabolic patchwork in the nested symbiosis of mealybugs. Curr. Biol. 2011, 21, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- López-Madrigal, S.; Latorre, A.; Porcar, M.; Moya, A.; Gil, R. Complete genome sequence of “Candidatus Tremblaya princeps” strain PCVAL, an intriguing translational machine below the living-cell status. J. Bacteriol. 2011, 193, 5587–5588. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.M.; Moran, N.A. Small, smaller, smallest: The origins and evolution of ancient dual symbioses in a phloem-feeding insect. Genome Biol. Evol. 2013, 5, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.G.; Wilson, A.C.C. A substrate ambiguous enzyme facilitates genome reduction in an intracellular symbiont. BMC Biol. 2014, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Gil, R.; Latorre, A. The evolutionary history of symbiotic associations among bacteria and their animal hosts: A model. Clin. Microbiol. Infect. 2009, 15, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Sudakaran, S.; Kost, C.; Kaltenpoth, M. Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol. 2017, 25, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.C.; Duncan, R.P. Signatures of host/symbiont genome coevolution in insect nutritional endosymbioses. Proc. Natl. Acad. Sci. USA 2015, 112, 201423305. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Latorre, A.; Sabater-Muñoz, B.; Moya, A.; Moran, N.A. Side-stepping secondary symbionts: Widespread horizontal transfer across and beyond the Aphidoidea. Mol. Ecol. 2003, 12, 1061–1075. [Google Scholar] [CrossRef] [PubMed]
- Nováková, E.; Hypsa, V.; Moran, N.A. Arsenophonus, an emerging clade of intracellular symbionts with a broad host distribution. BMC Microbiol. 2009, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Dale, C.; Welburn, S.C. The endosymbionts of tsetse flies: Manipulating host-parasite interactions. Int. J. Parasitol. 2001, 31, 628–631. [Google Scholar] [CrossRef]
- Moran, N.A.; Russell, J.A.; Koga, R.; Fukatsu, T. Evolutionary relationships of three new species of Enterobacteriaceae living as symbionts of aphids and other insects. Appl. Environ. Microbiol. 2005, 71, 3302–3310. [Google Scholar] [CrossRef] [PubMed]
- Brumin, M.; Kontsedalov, S.; Ghanim, M. Rickettsia influences thermotolerance in the whitefly Bemisia tabaci B biotype. Insect Sci. 2011, 18, 57–66. [Google Scholar] [CrossRef]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Nikoh, N.; Ijichi, N.; Shimada, M.; Fukatsu, T. Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect. Proc. Natl. Acad. Sci. USA 2002, 99, 14280–14285. [Google Scholar] [CrossRef] [PubMed]
- Dunning-Hotopp, J.C.; Clark, M.E.; Oliveira, D.C.S.G.; Foster, J.M.; Fischer, P.; Muñoz-Torres, M.C.; Giebel, J.D.; Kumar, N.; Ishmael, N.; Wang, S.; et al. Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes. Science 2007, 317, 1753–1756. [Google Scholar] [CrossRef] [PubMed]
- Woolfit, M.; Iturbe-Ormaetxe, I.; McGraw, E.A.; O’Neill, S.L. An ancient horizontal gene transfer between mosquito and the endosymbiotic bacterium Wolbachia pipientis. Mol. Biol. Evol. 2009, 26, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Blaxter, M. Symbiont genes in host genomes: Fragments with a future? Cell Host Microbe 2007, 2, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Tanaka, K.; Shibata, F.; Kondo, N.; Hizume, M.; Shimada, M.; Fukatsu, T. Wolbachia genome integrated in an insect chromosome: Evolution and fate of laterally transferred endosymbiont genes. Genome Res. 2008, 18, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Nakabachi, A. Aphids acquired symbiotic genes via lateral gene transfer. BMC Biol. 2009, 7. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; McCutcheon, J.P.; Kudo, T.; Miyagishima, S.; Moran, N.A.; Nakabachi, A. Bacterial genes in the aphid genome: Absence of functional gene transfer from Buchnera to its host. PLoS Genet. 2010, 6, e1000827. [Google Scholar] [CrossRef] [PubMed]
- Husnik, F.; Nikoh, N.; Koga, R.; Ross, L.; Duncan, R.P.; Fujie, M.; Tanaka, M.; Satoh, N.; Bachtrog, D.; Wilson, A.C.; et al. Horizontal gene transfer from diverse bacteria to an insect genome enables a tripartite nested mealybug symbiosis. Cell 2013, 153, 1567–1578. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.B.; Nakabachi, A.; Richards, S.; Qu, J.; Murali, S.C.; Gibbs, R.A.; Moran, N.A. Parallel histories of horizontal gene transfer facilitated extreme reduction of endosymbiont genomes in sap-feeding insects. Mol. Biol. Evol. 2014, 31, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.-B.; Chen, W.; Hasegawa, D.K.; Simmons, A.M.; Wintermantel, W.M.; Ling, K.-S.; Fei, Z.; Liu, S.S.; Douglas, A.E. Metabolic coevolution in the bacterial symbiosis of whiteflies and related plant sap-feeding insects. Genome Biol. Evol. 2015, 7, 2635–2647. [Google Scholar] [CrossRef] [PubMed]
- Bastolla, U.; Moya, A.; Viguera, E.; van Ham, R.C.H.J. Genomic determinants of protein folding thermodynamics in prokaryotic organisms. J. Mol. Biol. 2004, 343, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Kupper, M.; Gupta, S.K.; Feldhaar, H.; Gross, R. Versatile roles of the chaperonin GroEL in microorganism-insect interactions. FEMS Microbiol. Lett. 2014, 353, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, S.; Watanabe, H.; Hattori, M.; Sakaki, Y.; Ishikawa, H. Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS. Nature 2000, 407, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Park, J.T. An anhydro-N-acetylmuramyl-l-alanine amidase with broad specificity tethered to the outer membrane of Escherichia coli. J. Bacteriol. 2007, 189, 5634–5641. [Google Scholar] [CrossRef] [PubMed]
- Jollès, P. Lysozymes: Model Enzymes in Biochemistry and Biology, 1st ed.; Birkhäuser: Basel, Switzerland, 1996; p. 449. [Google Scholar]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; et al. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Valero, L.; Soriano-Navarro, M.; Pérez-Brocal, V.; Heddi, A.; Moya, A.; García-Verdugo, J.M.; Latorre, A. Coexistence of Wolbachia with Buchnera aphidicola and a secondary symbiont in the aphid Cinara cedri. J. Bacteriol. 2004, 186, 6626–6633. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, M.; Koga, R.; Tsuchida, T.; Meng, X.-Y.; Fukatsu, T. Rickettsia symbiont in the pea aphid Acyrthosiphon pisum: Novel cellular tropism, effect on host fitness, and interaction with the essential symbiont Buchnera. Appl. Environ. Microbiol. 2005, 71, 4069–4075. [Google Scholar] [CrossRef] [PubMed]
- Nakabachi, A.; Ishida, K.; Hongoh, Y.; Ohkuma, M.; Miyagishima, S. Aphid gene of bacterial origin encodes a protein transported to an obligate endosymbiont. Curr. Biol. 2014, 24, R640–641. [Google Scholar] [CrossRef] [PubMed]
- Gruwell, M.E.; Hardy, N.B.; Gullan, P.J.; Dittmar, K. Evolutionary relationships among primary endosymbionts of the mealybug subfamily Phenacoccinae (Hemiptera: Coccoidea: Pseudococcidae). Appl. Environ. Microbiol. 2010, 76, 7521–7525. [Google Scholar] [CrossRef] [PubMed]
- Von Dohlen, C.D.; Kohler, S.; Alsop, S.T.; McManus, W.R. Mealybug beta-proteobacterial endosymbionts contain gamma-proteobacterial symbionts. Nature 2001, 412, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Thao, M.; Gullan, P.; Baumann, P. Secondary (γ-Proteobacteria.) endosymbionts infect the primary (β-Proteobacteria.) endosymbionts of mealybugs multiple times and coevolve with their hosts. Appl. Environ. Microbiol. 2002, 68, 3190–3197. [Google Scholar] [CrossRef] [PubMed]
- Husnik, F.; McCutcheon, J.P. Repeated replacement of an intrabacterial symbiont in the tripartite nested mealybug symbiosis. Proc. Natl. Acad. Sci. USA 2016, 113, E5416–5424. [Google Scholar] [CrossRef] [PubMed]
- López-Madrigal, S.; Balmand, S.; Latorre, A.; Heddi, A.; Moya, A.; Gil, R. How does Tremblaya princeps get essential proteins from its nested partner Moranella endobia in the mealybug Planoccocus citri? PLoS ONE 2013, 8, e77307. [Google Scholar] [CrossRef] [PubMed]
- Thao, M.L.; Moran, N.A.; Abbot, P.; Brennan, E.B.; Burckhardt, D.H.; Baumann, P. Cospeciation of psyllids and their primary prokaryotic endosymbionts. Appl. Environ. Microbiol. 2000, 66, 2898–2905. [Google Scholar] [CrossRef] [PubMed]
- Nakabachi, A.; Ueoka, R.; Oshima, K.; Teta, R.; Mangoni, A.; Gurgui, M.; Oldham, N.J.; van Echten-Deckert, G.; Okamura, K.; Yamamoto, K.; et al. Defensive bacteriome symbiont with a drastically reduced genome. Curr. Biol. 2013, 23, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Santos-Garcia, D.; Farnier, P.-A.; Beitia, F.; Zchori-Fein, E.; Vavre, F.; Mouton, L.; Moya, A.; Latorre, A.; Silva, F.J. Complete genome sequence of “Candidatus Portiera aleyrodidarum” BT-QVLC, an obligate symbiont that supplies amino acids and carotenoids to Bemisia tabaci. J. Bacteriol. 2012, 194, 6654–6655. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.B.; Moran, N.A. Endosymbiotic bacteria as a source of carotenoids in whiteflies. Biol. Lett. 2012, 8, 986–989. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-F.; Xia, F.; Johnson, K.W.; Brown, C.D.; Bartom, E.; Tuteja, J.H.; Stevens, R.; Grossman, R.L.; Brumin, M.; White, K.P.; et al. Comparison of the genome sequences of “Candidatus Portiera aleyrodidarum” primary endosymbionts of the whitefly Bemisia tabaci B and Q biotypes. Appl. Environ. Microbiol. 2013, 79, 1757–1759. [Google Scholar] [CrossRef] [PubMed]
- Popa, O.; Hazkani-Covo, E.; Landan, G.; Martin, W.; Dagan, T. Directed networks reveal genomic barriers and DNA repair bypasses to lateral gene transfer among prokaryotes. Genome Res. 2011, 21, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Andersson, S.G.E.; Battin, T.J.; Prosser, J.I.; Schimel, J.P.; Whitman, W.B.; Hallin, S. The ecological coherence of high bacterial taxonomic ranks. Nat. Rev. Microbiol. 2010, 8, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Andreani, N.A.; Hesse, E.; Vos, M. Prokaryote genome fluidity is dependent on effective population size. ISME J. 2017, 11, 1719–1721. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.R.; Bergstrom, C.T. Bacteria are different: Observations, interpretations, speculations, and opinions about the mechanisms of adaptive evolution in prokaryotes. Proc. Natl. Acad. Sci. USA 2000, 97, 6981–6985. [Google Scholar] [CrossRef] [PubMed]
- Maisnier-Patin, S.; Paulander, W.; Pennhag, A.; Andersson, D.I. Compensatory evolution reveals functional interactions between ribosomal proteins S12, L14 and L19. J. Mol. Biol. 2007, 366, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Davids, W.; Zhang, Z. The impact of horizontal gene transfer in shaping operons and protein interaction networks—Direct evidence of preferential attachment. BMC Evol. Biol. 2008, 8. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Huang, H.V. Homologous recombination in Escherichia coli: Dependence on substrate length and homology. Genetics 1986, 112, 441–457. [Google Scholar] [PubMed]
- Lorenz, M.G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev. 1994, 58, 563–602. [Google Scholar] [PubMed]
- Majewski, J.; Cohan, F.M. DNA sequence similarity requirements for interspecific recombination in Bacillus. Genetics 1999, 153, 1525–1533. [Google Scholar] [PubMed]
- McCutcheon, J.P.; McDonald, B.R.; Moran, N.A. Origin of an alternative genetic code in the extremely small and GC-rich genome of a bacterial symbiont. PLoS Genet. 2009, 5, e1000565. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.A.C.; Hickey, D.A. Nucleotide bias causes a genomewide bias in the amino acid composition of proteins. Mol. Biol. Evol. 2000, 17, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Boël, G.; Letso, R.; Neely, H.; Price, W.N.; Wong, K.-H.; Su, M.; Luff, J.; Valecha, M.; Everett, J.K.; Acton, T.B.; et al. Codon influence on protein expression in E. coli correlates with mRNA levels. Nature 2016, 529, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, J.; Gill, R.T. Amino acid content of recombinant proteins influences the metabolic burden response. Biotechnol. Bioeng. 2005, 90, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.M.; Kemper, A.; Gruber-Vodicka, H.; Cardini, U.; van der Geest, M.; Kleiner, M.; Bulgheresi, S.; Mußmann, M.; Herbold, C.; Seah, B.K.; et al. Chemosynthetic symbionts of marine invertebrate animals are capable of nitrogen fixation. Nat. Microbiol. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Dietrich, C.; Brune, A. Genome analysis of Endomicrobium proavitum suggests loss and gain of relevant functions during the evolution of intracellular symbionts. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.F.; Scharf, M.E. Lower termite associations with microbes: Synergy, protection, and interplay. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [PubMed]
- Gil, R.; Vargas, C.; López-Madrigal, S.; Santos-García, D.; Latorre, A.; Moya, A. Tremblaya phenacola PPER: An evolutionary beta-gammaproteobacterium collage. ISME J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.-Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Penz, T.; Schmitz-Esser, S.; Kelly, S.E.; Cass, B.N.; Müller, A.; Woyke, T.; Malfatti, S.A.; Hunter, M.S.; Horn, M. Comparative genomics suggests an independent origin of cytoplasmic incompatibility in Cardinium hertigii. PLoS Genet. 2012, 8, e1003012. [Google Scholar] [CrossRef] [PubMed]
- Sait, M.; Aitchison, K.; Wheelhouse, N.; Wilson, K.; Lainson, F.A.; Longbottom, D.; Smith, D.G. Genome sequence of Lawsonia intracellularis strain N343, isolated from a sow with hemorrhagic proliferative enteropathy. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.J.; Joardar, V.; Williams, K.P.; Driscoll, T.; Hostetler, J.B.; Nordberg, E.; Shukla, M.; Walenz, B.; Hill, C.A.; Nene, V.M.; et al. A Rickettsia genome overrun by mobile genetic elements provides insight into the acquisition of genes characteristic of an obligate intracellular lifestyle. J. Bacteriol. 2012, 194, 376–394. [Google Scholar] [CrossRef] [PubMed]
- Sjödin, A.; Svensson, K.; Ohrman, C.; Ahlinder, J.; Lindgren, P.; Duodu, S.; Johansson, A.; Colquhoun, D.J.; Larsson, P.; Forsman, M. Genome characterisation of the genus Francisella reveals insight into similar evolutionary paths in pathogens of mammals and fish. BMC Genomics 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Kellner, R.L.L.; Dettner, K. Differential efficacy of toxic pederin in deterring potential arthropod predators of Paederus (Coleoptera: Staphylinidae) offspring. Oecologia 1996, 107, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Kellner, R.L.L. Molecular identification of an endosymbiotic bacterium associated with pederin biosynthesis in Paederus sabaeus (Coleoptera: Staphylinidae). Insect Biochem. Mol. Biol. 2002, 32, 389–395. [Google Scholar] [CrossRef]
- Manley, G.V. Paederus fuscipes [Col.: Staphylinidae]: A predator of rice fields in west Malaysia. Entomophaga 1977, 22, 47–59. [Google Scholar] [CrossRef]
- Jain, R.; Rivera, M.C.; Lake, J.A. Horizontal gene transfer among genomes: The complexity hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 3801–3806. [Google Scholar] [CrossRef] [PubMed]
- Cohen, O.; Gophna, U.; Pupko, T. The complexity hypothesis revisited: Connectivity rather than function constitutes a barrier to horizontal gene transfer. Mol. Biol. Evol. 2011, 28, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Szabó, G.; Schulz, F.; Toenshoff, E.R.; Volland, J.-M.; Finkel, O.M.; Belkin, S.; Horn, M. Convergent patterns in the evolution of mealybug symbioses involving different intrabacterial symbionts. ISME J. 2017, 11, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Comas, I.; Moya, A.; Azad, R.K.; Lawrence, J.G.; Gonzalez-Candelas, F. The evolutionary origin of Xanthomonadales genomes and the nature of the horizontal gene transfer process. Mol. Biol. Evol. 2006, 23, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- López-Madrigal, S.; Latorre, A.; Porcar, M.; Moya, A.; Gil, R. Mealybugs nested endosymbiosis: Going into the “matryoshka” system in Planococcus citri in depth. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- López-Madrigal, S.; Latorre, A.; Moya, A.; Gil, R. The link between independent acquisition of intracellular gamma-endosymbionts and concerted evolution in Tremblaya princeps. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
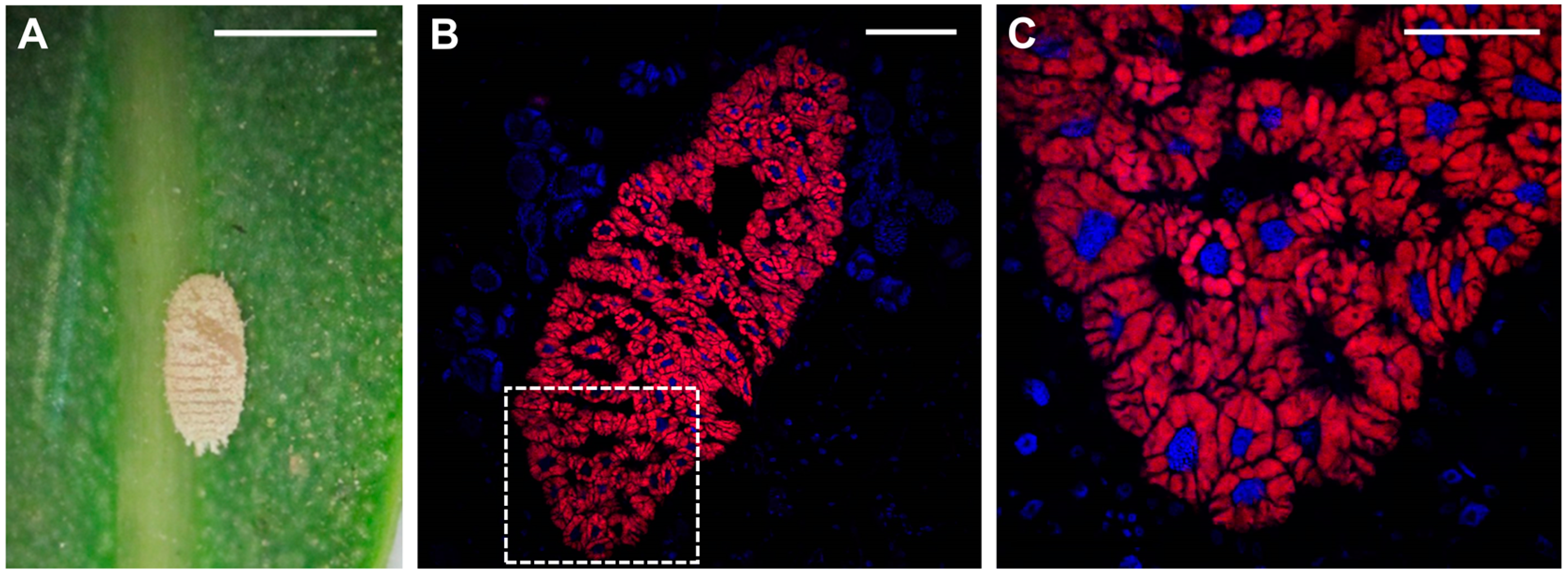
Figure 1.
Phenacoccus peruvianus/”Candidatus Tremblaya phenacola” symbiotic system. (A) Early nymph of P. peruvianus; (B,C) Confocal images showing a complete bacteriome section (B) and the magnification of the area within the dashed square (C). DAPI (4′,6-diamidino-2-phenylindole)-stained nuclei appear in blue; EUB338-probed bacteria appear in red. Fluorescence in Situ Hybridization (FISH) procedure is described in [55]. Scale bars: 1 mm (A), 100 μm (B), 50 μm (C).
Figure 1.
Phenacoccus peruvianus/”Candidatus Tremblaya phenacola” symbiotic system. (A) Early nymph of P. peruvianus; (B,C) Confocal images showing a complete bacteriome section (B) and the magnification of the area within the dashed square (C). DAPI (4′,6-diamidino-2-phenylindole)-stained nuclei appear in blue; EUB338-probed bacteria appear in red. Fluorescence in Situ Hybridization (FISH) procedure is described in [55]. Scale bars: 1 mm (A), 100 μm (B), 50 μm (C).

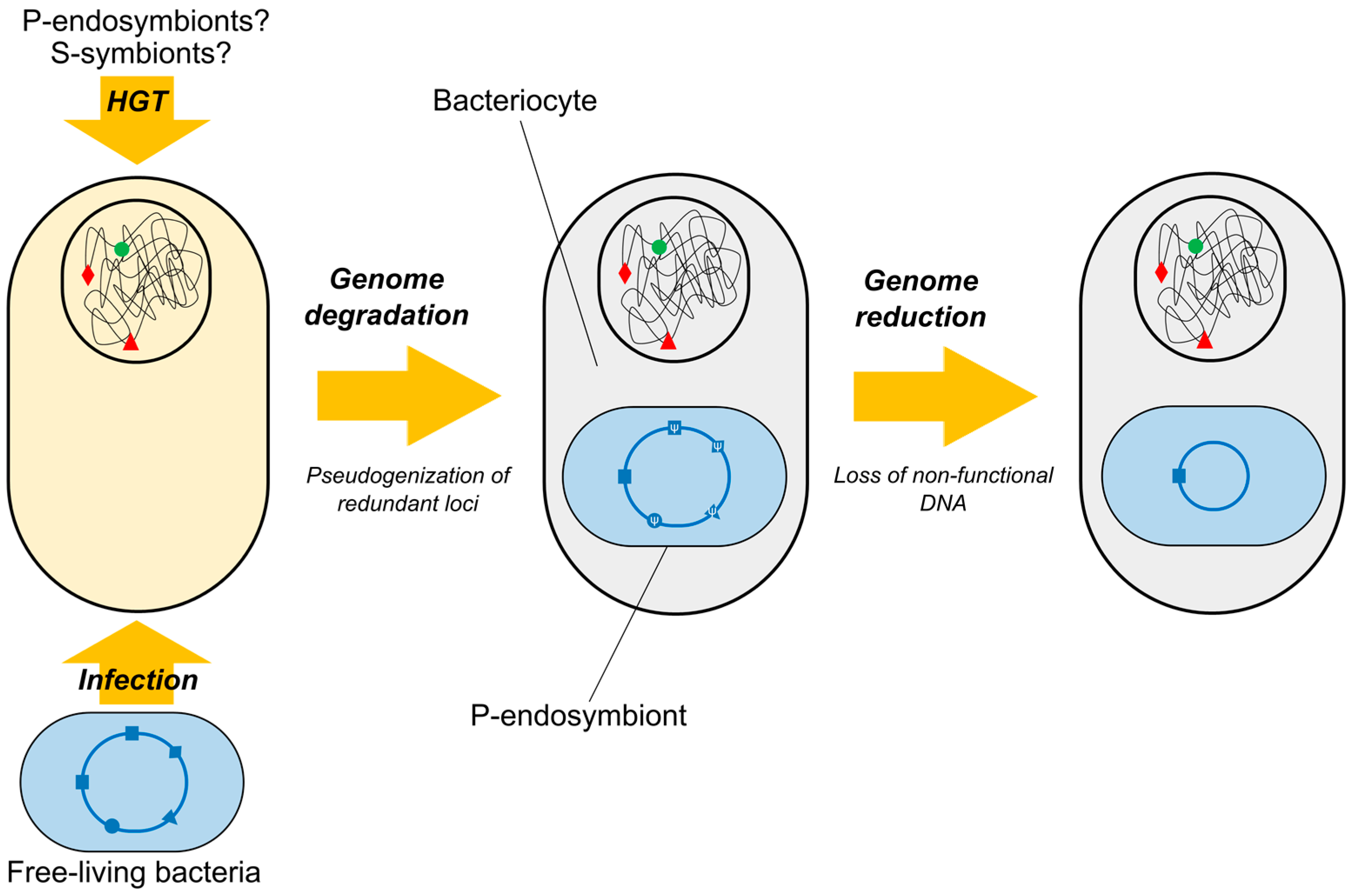
Figure 2.
Reductive evolution of P-endosymbionts in insects whose genomes carry loci of prokaryotic origin. Horizontally acquired genes from ancient P-endosymbionts/S-symbionts (green/red symbols), as well as ongoing reductive evolution of current P-endosymbiont through inactivation and loss of redundant loci (square, triangle, rhombus, circle) are shown. HGT: Horizontal Gene Transfer.
Figure 2.
Reductive evolution of P-endosymbionts in insects whose genomes carry loci of prokaryotic origin. Horizontally acquired genes from ancient P-endosymbionts/S-symbionts (green/red symbols), as well as ongoing reductive evolution of current P-endosymbiont through inactivation and loss of redundant loci (square, triangle, rhombus, circle) are shown. HGT: Horizontal Gene Transfer.


{kind=link}
{kind=link}
Table 1.
Bacteria-to-host genetic transfers. Horizontally acquired genes mediating insect-bacteria nutritional symbioses.
Table 1.
Bacteria-to-host genetic transfers. Horizontally acquired genes mediating insect-bacteria nutritional symbioses.
| Host | Endosymbiont | Gene Number | Function | Source | Ref. |
|---|---|---|---|---|---|
| Acyrthosiphon pisum | Buchnera aphidicola | 12 | peptidoglycan metabolism (defensive, control) | Rickettsia, Wolbachia | [88,89] |
| Planococcus citri | “Ca. Tremblaya princeps” “Ca. Moranella endobia” | 22 | Lys, Met, riboflavin and biotin biosynthesis (nutritional) peptidoglycan metabolism (control) | Arsenophonus, Cardinium, Rickettsia, Serratia, Sodalis, Wolbachia | [90] |
| Pachypsylla venusta | “Ca. Carsonella ruddii” | 10 | Phe, Arg, riboflavin biosynthesis (nutritional) DNA mismatch repair (informational) | Carsonella, Rickettsia, Wolbachia | [91] |
| Bemisia tabaci | “Ca. Portiera aleyrodidarum” “Ca. Hamiltonella defensa” | 10 | Arg, Lys, Phe, thiamine biosynthesis/ urea degradation (nutritional) | Pantoea & stinkbugs gut symbionts, Rickettsiales, Niastella, Isosphaera | [92] |
Table 2.
Bacteria-to-bacteria horizontal gene transfer (HGT) events involving nutritional endosymbionts of insects.
Table 2.
Bacteria-to-bacteria horizontal gene transfer (HGT) events involving nutritional endosymbionts of insects.
| Host | Endosymbiont | Gene Number | Function | Source | Ref. |
|---|---|---|---|---|---|
| Diaphorina citri | “Ca. Carsonella ruddii” “Ca. Profftella armatura” | 20 | diaphorin biosynthesis (defensive) | Paederus-associated Pseudomonas | [108] |
| Cimex lectularius | Wolbachia pipientis | 9 | biotin and thiamine biosynthesis (nutritional) | Cardinium, Rickettsia | [128] |
| Phenacoccus peruvianus | “Ca. Tremblaya phenacola” | 80 | nutritional informational | Sodalis-allied clade | [129] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
López-Madrigal, S.; Gil, R. Et tu, Brute? Not Even Intracellular Mutualistic Symbionts Escape Horizontal Gene Transfer. Genes 2017, 8, 247. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8100247
AMA Style
López-Madrigal S, Gil R. Et tu, Brute? Not Even Intracellular Mutualistic Symbionts Escape Horizontal Gene Transfer. Genes. 2017; 8(10):247. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8100247
Chicago/Turabian StyleLópez-Madrigal, Sergio, and Rosario Gil. 2017. "Et tu, Brute? Not Even Intracellular Mutualistic Symbionts Escape Horizontal Gene Transfer" Genes 8, no. 10: 247. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8100247
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.