The Small and the Dead: A Review of Ancient DNA Studies Analysing Micromammal Species


Department of Earth Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, UK
*
Authors to whom correspondence should be addressed.
Genes 2017, 8(11), 312; https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110312
Submission received: 23 August 2017
/
Revised: 13 October 2017
/
Accepted: 13 October 2017
/
Published: 8 November 2017
(This article belongs to the Special Issue Novel and Neglected Areas of Ancient DNA Research)
Abstract
:The field of ancient DNA (aDNA) has recently been in a state of exponential growth, largely driven by the uptake of Next Generation Sequencing (NGS) techniques. Much of this work has focused on the mammalian megafauna and ancient humans, with comparatively less studies looking at micromammal fauna, despite the potential of these species in testing evolutionary, environmental and taxonomic theories. Several factors make micromammal fauna ideally suited for aDNA extraction and sequencing. Micromammal subfossil assemblages often include the large number of individuals appropriate for population level analyses, and, furthermore, the assemblages are frequently found in cave sites where the constant temperature and sheltered environment provide favourable conditions for DNA preservation. This review looks at studies that include the use of aDNA in molecular analysis of micromammal fauna, in order to examine the wide array of questions that can be answered in the study of small mammals using new palaeogenetic techniques. This study highlights the bias in current aDNA studies and assesses the future use of aDNA as a tool for the study of micromammal fauna.
1. Introduction
Techniques to enable extraction and sequencing of DNA from museum specimens, zooarchaeological and sub fossil material [1] have continued to develop since the initial successful recovery of ancient DNA (aDNA) in 1984 [2]. Whilst many of the earliest aDNA studies focused on taxonomy and resolving the phylogenetic placement of extinct species, the development of new techniques, and particularly the use of next generation sequencing (NGS), has allowed the field to expand into ancient pathogens [3,4], population demographic studies [5], conservation issues [6], domestication events [7] and palaeoecological studies [8]. The field of aDNA has, however, largely focused on mammal species [9,10], with a strong bias towards large, charismatic species such as mammoths [11,12], and humans and their closest relatives [13,14,15]. This bias, however, does little to reflect real world species diversity with its plethora of small-bodied mammals. For example, half of all placental diversity is contained within the order Rodentia, a highly speciose order principally composed of small bodied taxa [16].
Here, we define small-bodied or micromammals as any mammal species under 500 g. Definitions of micromammals have typically included members of several orders including Rodentia, Eulipotyphla and occasionally Lagomorpha, Carnivora (family: Mustelidae), and Chiroptera [17]. There is a wealth of research questions that can be addressed via the study of micromammal fauna. These species show rapid generational turnover [18], which is associated with a high mutation rate [19,20,21] and an associated ability to swiftly adapt to new and/or changing environments [22]. There is a strong correlation between mutation rate and systematic components of mammalian life history traits, with rates varying consistently with parameters associated with body size, population dynamics and lifestyle [23]. Mitochondrial substitution rates within the Rodentia, for example, have been estimated to be around 20 times faster than those observed in the Cetacea (whales, dolphins and porpoises), the latter of which show larger body mass, increased longevity and slower generational turnover times [21]. This allows the identification of population level, biogeographic and adaptive responses to past changes in both biotic and abiotic environmental variables. Small bodied mammals typically have high dispersal abilities [24]—a factor that facilitates frequent island colonisation events and thus further facilitates studies relating to biogeography and speciation. Micromammals are also considered key biostratigraphic indicators, providing a source of valuable proxy data with which to infer palaeoecological, palaeoclimatic and palaeogeographical change [25,26,27]. This is due to their widespread abundance in faunal assemblages, their excellent fossil record, and their rapid rate of morphological and genetic change [28,29].
Whilst micromammals can make excellent model study species covering both diverse areas of interest and important concepts in evolutionary biology, palaeontology and conservation, they have been vastly under-represented in ancient DNA studies. Consequently, a review to highlight the potential of working with aDNA from small mammals is long overdue. This review catalogues past efforts to use aDNA to study micromammals and discusses prospects for the future of this field.
2. Methods
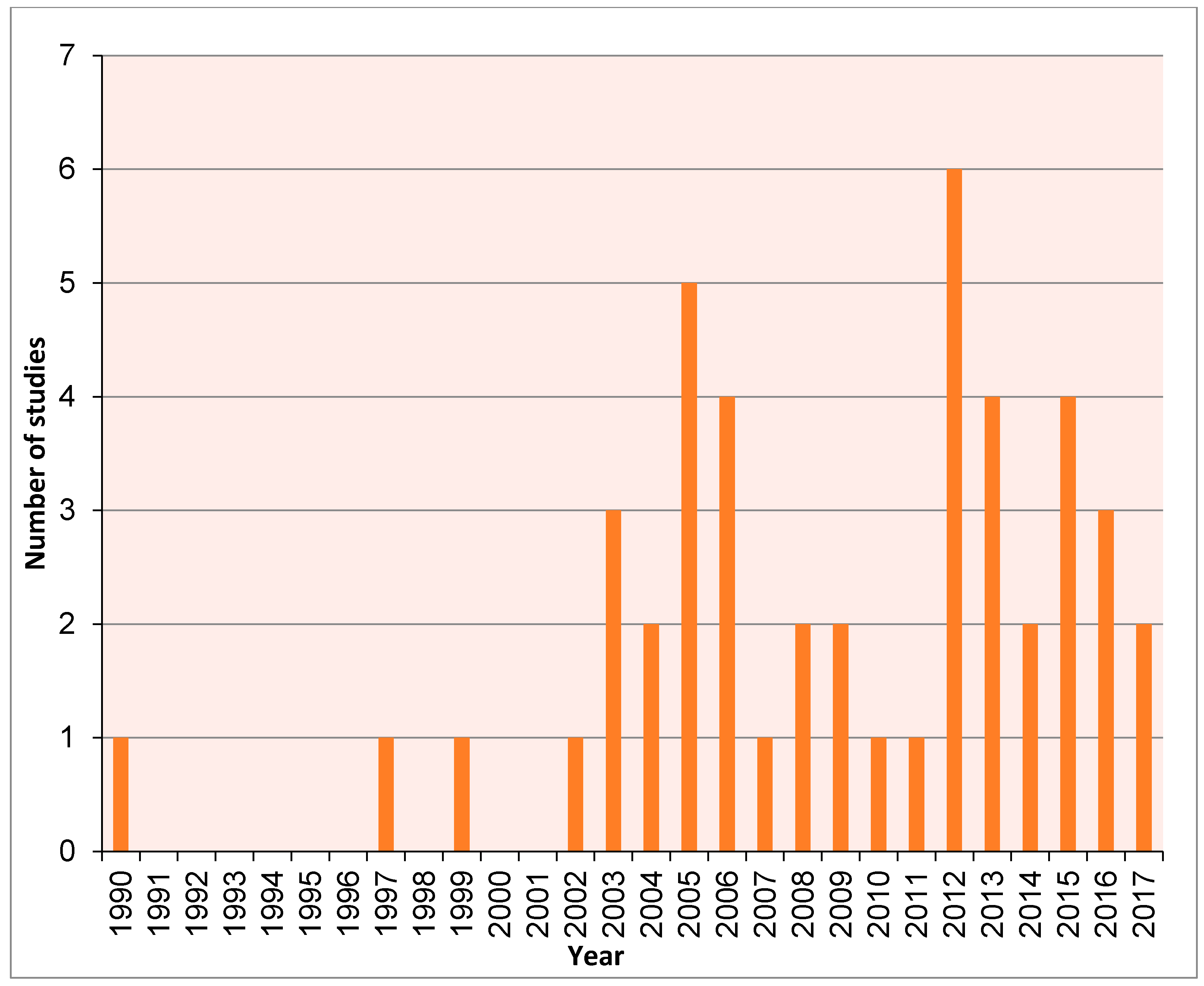
This review includes a meta-analysis literature review of studies that utilise ancient DNA techniques to examine micromammal fauna, encompassing a range of materials from degraded museum specimens to sub-fossils. Articles compiled for this review were extracted from ISI Web of Science, PubMed and Google Scholar in 2017 using several search terms including: “ancient DNA”, “aDNA”, “historic DNA”, “palaeogenomic”, “paleogenomic”, “palaeogenetic”, “paleogenetic”, “museum” and “micromammal”, “micro mammal”, “small mammal”. We then assessed a number of articles over time (Figure 1) and potential taxonomic bias (Figure 2).
3. Results and Discussion
The following sections of this review address key trends associated with ancient DNA studies utilising micromammal fauna. The list is not exhaustive but rather represents the major themes observed in relation to: technology, source material and the varied questions that small mammal aDNA studies are being employed to address.
3.1. Next Generation Sequencing
The study of ancient DNA has been revolutionised by the emergence of new technologies, namely the shift from PCR and traditional Sanger sequencing to NGS [30]. This shift has impacted the field in numerous ways including the challenges faced by researchers hoping to extract endogenous genetic material from degraded specimens. Previous PCR based studies relied on designing primers, a difficult task when studying extinct species with no previously recorded molecular data. The benefit of species-specific primers however is that they target both the region and species of interest, thus reducing the likelihood of sequencing contaminant DNA. In contrast, NGS shotgun approaches have no specificity, with all DNA in a sample, endogenous and contaminant being sequenced. As an alternative to shotgun sequencing, target capture enrichment techniques can be applied prior to sequencing to increase the specificity of the sequencing process, but, as with PCR, the process requires reference sequences from which to design the “baits” to capture the DNA [31,32]. Different micromammal species have different levels of potential for these approaches. For species that are widely used in genetic studies (such as rats or mice), reference sequences are available, for other less studied, rare or extinct species, reference sequences may be unavailable, restricting analyses to either de novo assembly or aligning to the evolutionarily closest (presently often quite distant) sequenced relative [33,34].
3.2. DNA Metabarcoding
Micromammal remains, particularly rodents and insectivores, are often found in large quantities that provide limited morphological value. These large assemblages are perhaps optimally analysed using metabarcoding to retrieve viable data for ecological, taxonomic or biogeographical studies. Metabarcoding permits mass sequencing of genetic data using NGS. This technique uses universal PCR primers to mass-amplify DNA barcodes from large collections of specimens or from environmental DNA (eDNA). The resulting sequence data can then be verified and identified to species or genus level using a database of existing DNA data. A study by Guimaraes et al. [35] successfully used DNA barcoding to identify rodent species from a variety of differently preserved specimens and as part of this study correctly identified 75.9% of museum samples and 81.8% of species from owl pellets. The study additionally analysed up to 44,000-year-old rodent material from a North African cave, illustrating that barcoding can also be applied to samples typically identified as difficult to sequence.
3.3. Cave Sites
Micromammal remains are often found in cave environments where they form a large proportion of recovered faunal elements [36]. This may be the result of several factors including ecology (the cave was the natural habitat of the species), taphonomy (the small mammals were prey species for some cave-living avian or mammalian predator), or demography (the larger population size of small mammals simply increases the likelihood of their preservation). DNA recovery also seems to be higher in material from cave sites, possibly due to the low variance in temperature which may help to slow down hydrolytic processes involved in DNA degradation, thus preserving DNA that may, outside of the cave environment, have become further fragmented [37,38]. Many aDNA studies including Guimaraes et al. [35], have taken advantage of cave site remains. Gutiérrez-García et al. [39] is a further study where aDNA was successfully analysed from rodents (in this study Ototylomys phyllotis) that were excavated from a cave site in a hot/humid climate. This further highlights that micromammals are an excellent study species for aDNA studies, particularly in places such as the tropics where DNA preservation outside of cave environments is poor.
3.4. Avian Pellets
Recent and palaeontological pellets from raptors, particularly owls, can provide a valuable source of micromammal skeletal material. Recent pellets can provide a non-invasive method to sample current populations [40], whilst palaeontological pellets can provide a temporal snapshot of the micromammal fauna from a specific place and time [17]. Skeletal and dental material recovered from owl pellets can show excellent morphological preservation in some instances [41,42,43]. However, many studies require identification to species level and this can be problematic unless taxonomically informative elements, which are also in a good state of structural preservation, are recovered. In the absence of such specimens (i.e., in cases where only uninformative postcranial bones are recovered from the pellet), molecular methods may be the most feasible means to address ecological, biogeographical and taxonomic questions.
Pellets are commonly used to find and collect genetic material for extant micromammal populations, as they can provide large numbers of individuals and a variety of species. They can additionally be employed to estimate the geographic distribution of micromammals if the range of the predator species is known. An initial study by Taberlet and Fumagalli [44] successfully sequenced micromammal DNA from owl pellets, but found that the DNA quality was poor. A second study that applied specific aDNA techniques further suggested that extracting DNA from even the most recent specimens recovered from owl pellets warranted an aDNA approach [45]. A recent study [46] confirmed that the quantity of DNA preserved in owl pellets was low but additionally reported high variability in preservation between bones even within the same pellet. It therefore seems likely that the continued development of aDNA techniques will serve to increase the use of owl pellets as a valuable source of genetic data for the study of micromammal fauna, but that the inclusion of a screening step may prove useful to account for the variability in DNA preservation.
3.5. Resolving the Phylogenetic Placement of Extinct Species
aDNA studies have been particularly useful in resolving the phylogenetic placement of rare or extinct groups or individual species. In the case of specious groups, such as insectivores and rodents, aDNA has helped resolve long-standing evolutionary questions. An example is the extinct Caribbean eulipotyphlan insectivore Nesophontes. Brace et al. [47] applied ancient DNA techniques to resolve the phylogenetic placement of Nesophontes by recovering a near-complete mitochondrial genome and 17 nuclear genes from a 750-year-old specimen. This study showed that, despite their morphological differences, Nesophontes formed a monophylectic clade with the extant Caribbean insectivore Solenodon. It further highlighted that this was an ancient divergence, as the two genera shared a common ancestor more than 40 million years ago. Other aDNA studies of an extinct island endemic include the Lava mouse Malpaisomys insularis [48]. Here, a more recent divergence was identified as this study revealed that M. insularis diverged from its morphologically incongruous closest mainland relative only 6.9 Ma.
3.6. Species Identification
Until the advent of ancient DNA analyses, the principle methods of identifying zooarchaeological and subfossil micromammals were via visual inspection and/or measurements taken from skeletal or dental elements e.g., [49,50]. The development of geometric morphometric methods greatly improved the sensitivity of statistical techniques for species identification but these usually incorporate some degree of error [51,52,53]. In addition, all morphological techniques are vulnerable to the confounding factors of physical damage and weak taxonomic signal. Many micromammal groups are highly species rich (e.g., Rodentia [54]) and rapid rates of diversification can result in genetic speciation without a concordant proliferation of morphological differences [55]. In addition, some species groups show high levels of phenotypic plasticity (e.g., shrews (Sorex) [56] and Arvicoline rodents [28,57]). When high levels of intra-specific variation are coupled with low levels of inter-specific distinction, true evolutionary relationships can be obscured due to morphological overlap.
Ancient DNA provides a crucial second line of evidence in scenarios where species cannot be easily identified on the basis of morphology alone and can also be used in isolation where morphological structures are not well preserved. However, it should be kept in mind that this approach also presents several challenges. Incorrect phylogenetic placement may occur due to low read coverage (a pervasive problem in shotgun sequenced NGS datasets with low endogenous DNA content, [30,58]), and associated incorporation of mis-called bases into the final consensus sequence. In addition, interspecific introgression of mitochondrial DNA has been commonly observed among small mammals (for example, in the rodent genus Myodes [59,60]) and can present significant problems in phylogenetic reconstruction [61]. Invasive sampling required for aDNA analyses are, by nature, destructive and this is particularly problematic with remains of small-bodied mammals. Molecular techniques are also limited with regards to the number of specimens that can be included in an analysis, while morphological methods can incorporate many hundreds of individuals.
Nevertheless, ancient DNA is a valuable tool in identification of species in the zooarchaeological and palaeontological record. Notably, it is particularly powerful when used to test hypotheses proposed by morphology. These studies often focus on extinct species or populations, and can provide additional data such as estimates of genetic distinction. Haring et al. [62] found that species delimitation based on genetic data from cave deposited specimens agreed with the previous morphometric analysis when examining far-eastern grey voles (Genus: Alexandromys). Rodrigues et al. [63] used both modern DNA and DNA extracted from museum specimens in order to test previous taxonomic hypotheses for the Egyptian weasel (Mustela subpalmata). This study found no evidence for genetic distinction between M. subplamata and the least weasel (Mustela nivalis) contrary to previous morphology based studies. Rodrigues et al. [63] suggested that the large size difference between these geographic populations and sexual dimorphism characteristic of M. subplamata are due to ecotypic rather than genetic variation. As the cost of sequencing continues to decrease, aDNA analyses become increasingly viable as a collaborative technique to identify species from archaeological sites.
3.7. Biogeographic Hypotheses and Island Micromammals
Micromammals are often adept dispersers and can therefore be found on many island systems [23,64]. As extinction rates on islands are higher than the mainland [65], aDNA studies play a pivotal role in our understanding of extinct or rare island micromamal evolution. Examples of studies that have used ancient DNA to examine island micromammals include: Caribbean rodents [66,67] and insectivores [47], the Easter island Pacific rat (Rattus exulans) [63,68,69], and the Christmas Island rat (Rattus macleari) [70]. A further aDNA study that examined the Orkney vole (Microtus arvalis orcadensis) explored colonisation dynamics, founder effects and genetic drift, all of which are associated with island populations [71].
3.8. Interactions with Humans
Using aDNA to explore our own human past history typically generates significant interest [72]. However, reconstructing historic human migrations can be a difficult task. Early PCR based studies were plagued by contamination from modern human samples [9] and whilst NGS techniques largely circumvent the problem, it remains relevant and validation of authenticity is required. When sample availability and permissions are also taken into account, an alternative approach to examining past human movements can be appealing. Due to their often-close interactions with humans, micromammals can make a useful proxy. Some species are domesticates (e.g., rats [73]), others have adapted to live alongside us (e.g., house mice [74]) to exploit the benefits we inadvertently provide, such as food or shelter, whilst humans have intentionally or accidentally transported and introduced other micromammals to new habitats [75]. Barnes et al. [68] analysed aDNA from the Pacific rat (Rattus exulans), a rodent intentionally transported by ancient Polynesians as a food item, in order to study the colonisation route of ancient humans in the Pacific.
3.9. Population Genetics
The use of ancient DNA in population level genetic and genomic studies can be somewhat problematic. Large numbers of individuals and/or markers are often required to fully resolve population level processes, and this is often difficult or impossible to achieve. Limiting factors in aDNA population level genetic analyses include availability/access to specimens and the variable quality of the DNA across samples. In contrast to many larger bodied species, micromammals are often found at high population densities [76], a factor that contributes to the increased likelihood of accessing larger numbers of micromammal specimens.
Hadley et al. [77] introduced a novel approach that used a combination of serial coalescent analyses with Approximate Bayesian Computation (ABC) [78] to examine past population structure, specifically a historic population bottleneck in the tuco-tucos (Ctenomys sociabillis). The species, a social subterranean rodent, had previously been identified as having almost no genetic variation (mitochondrial gene cytB) over the last 1000 years [79]. A further study [80] looked at samples from 3,000–10,000 years ago and found far higher levels of variation, suggesting a severe population decline and a bottleneck. It was to this bottleneck that Chan et al. [80] applied the ABC coalescent method. Studies such as these were amongst the first to show that aDNA could improve our estimation of the timing of events, such as population declines, allowing greater understanding of past populations and even the ability to inform practises for current populations.
3.10. Conservation
The use of ancient DNA data to inform conservation management and planning (termed conservation palaeogenetics; [6] or, alternately, conservation archaeogenetics; [81]) is an emerging field which holds much promise for wildlife conservation. Heterochronous aDNA datasets allow comparison of contemporary levels of genetic diversity, population size, population structure and geographical distributions with those that existed in the past [6,81,82]. This potentially allows conservation biologists to design and implement appropriate management programs based on data that is not biased by observations based only on extant populations that may be severely affected by human activity. While micromammals comprise some of the most globally abundant and widespread species (e.g., Microtus spp. [83]), they also contain a number of rare niche specialists (e.g., star nosed moles Condylura cristata [84]). To date, however, few conservation palaeogenetics studies have undertaken this focus.
Studies of Caribbean hutias [67,85] have shown that aDNA techniques can be used to analyse degraded museum and zooacheological specimens to examine living, highly endangered or rare species when sampling from wild populations is impractical. Endangered micromammals due to their small size can be particularly elusive in the wild. Museum samples can offer an alternative genetic resource to assess population structure, identify potential source populations and species boundaries to inform conservation efforts for endangered micromammal taxa [6,86,87,88]. For example, DNA extracted from museum specimens of the common hamster (Cricetus cricetus) allowed past levels of gene flow and population structure to be determined and this data subsequently formed the basis of population augmentation schemes in areas of the species range where it has suffered major declines [89].
3.11. Climate Change
Due to biological factors and their abundant presence in many archaeological and palaeontological sites, micromammals can act as excellent proxy indicators for past climate change. While early studies were constrained to use only presence-absence data (e.g., [25,26,27]), more recent research has been able to take advantage of ancient DNA evidence (e.g., [82]). Some (e.g., [90]) have combined traditional Ecological Niche Modelling with aDNA data to predict the results of future climate change. Other studies have used historic and ancient samples to investigate patterns of species continuity [35] and/or extinctions over time and fluctuating climates [91]. A further key factor in relation to climate change studies is that, prior to the large-scale, Neolithic, transition to agriculture [92], micromammals are free from the influence of anthropogenic activity. This allows the confounding factor of human-mediated effects to be removed. For example, Brace et al. [91] analysed collard lemming (Dicrostonyx torquatus) from Belgium over a 40,000 year time period (10–50 Kyr ago). They observed dramatic loss in genetic diversity during this period and identified a series of extinction and recolonisation events that could be related to abrupt climatic change.
4. Conclusions
Molecular analysis of sub fossil material, zooacheological skeletal remains and degraded museum specimens has transformed how we explore past histories, and has the potential to continue to do so as new techniques develop. Whilst many studies continue to apply the latest techniques to charismatic species such as extinct mega fauna and humans, groups such as micromammals conversely continue to remain an understudied and untapped information resource. This is despite the large numbers of specimens available, which, due to cave environments, often exhibit superior preservation. We identify micromammals as an important future direction for the field of ancient DNA. Our study has further highlighted that, even as an understudied group, there exists a bias towards rodent species in aDNA studies of micro-mammals (see Table 1). This bias could be a result of the widespread and speciose nature of the order Rodentia, or availability of specimens and close reference sequences. Many other micromammal orders, such as eulipotyphlan insectivores, remain drastically understudied in comparison, despite their potential to resolve taxonomy and provide information on past climates and environments. The general utility of micromammal assemblages to assess the response of small mammals to past climate change, however, is becoming an increasingly important and studied area of research, the results of which can also be applied to better inform studies of present day threats to individual species and biodiversity. As ancient DNA studies continue to move away from solely species identification and taxonomy and towards answering wider evolutionary questions, the inclusion of micromammals will become increasingly important.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hagelberg, E.; Hofreiter, M.; Keyser, C. Introduction. Ancient DNA: The first three decades. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20130371. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Bowman, B.; Freiberger, M.; Ryder, O.A.; Wilson, A.C. DNA sequences from the quagga, an extinct member of the horse family. Nature 1984, 312, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Bos, K.I.; Jäger, G.; Schuenemann, V.J.; Vågene, Å.J.; Spyrou, M.A.; Herbig, A.; Nieselt, K.; Krause, J. Parallel detection of ancient pathogens via array-based DNA capture. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20130375. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.; Ingham, S. Using ancient DNA analysis in palaeopathology: A critical analysis of published papers, with recommendations for future work. Int. J. Osteoarchaeol. 2008, 18, 600–613. [Google Scholar] [CrossRef]
- Hofreiter, M.; Barnes, I. Diversity Lost: Are All Holarctic Large Mammal Species Just Relict Populations? BMC Biol. 2010, 8, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.A. Ancient DNA applications for wildlife conservation. Mol. Ecol. 2008, 17, 4186–4196. [Google Scholar] [CrossRef] [PubMed]
- MacHugh, D.E.; Larson, G.; Orlando, L. Taming the Past: Ancient DNA and the study of animal domestication. Annu. Rev. Anim. Biosci. 2016, 5, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Anderson-Carpenter, L.L.; McLachlan, J.S.; Jackson, S.T.; Kuch, M.; Lumibao, C.Y.; Poinar, H.N. Ancient DNA from lake sediments: Bridging the gap between paleoecology and genetics. BMC Evol. Biol. 2011, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Hofreiter, M.; Serre, D.; Poinar, H.N.; Kuch, M.; Pääbo, S. Ancient DNA. Nat. Rev. Genet. 2001, 2, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Pääbo, S.; Poinar, H.; Serre, D.; Jaenicke-Despres, V.; Hebler, J.; Rohland, N.; Kuch, M.; Krause, J.; Vigilant, L.; Hofreiter, M. Genetic analyses from ancient DNA. Annu. Rev. Genet. 2004, 38, 645–679. [Google Scholar] [CrossRef] [PubMed]
- Palkopoulou, E.; Mallick, S.; Skoglund, P.; Enk, J.; Rohland, N.; Li, H.; Omrak, A.; Vartanyan, S.; Poinar, H.; Götherström, A.; et al. Complete genomes reveal signatures of demographic and genetic declines in the woolly mammoth. Curr. Biol. 2015, 25, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Knapp, M.; Enk, J.; Lippold, S.; Kircher, M.; Lister, A.; MacPhee, R.D.E.; Widga, C.; Czechowsk, P.; Sommer, R.; et al. The evolutionary and phylogeographic history of woolly mammoths: A comprehensive mitogenomic analysis. Sci. Rep. 2017, 7, 44585. [Google Scholar] [CrossRef] [PubMed]
- Dalton, R. Ancient DNA set to rewrite human history. Nature 2010, 465, 148–149. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Posth, C.; Hajdinjak, M.; Petr, M.; Mallick, S.; Fernandes, D.; Furtwängler, A.; Haak, W.; Meyer, M.; Mittnik, A.; et al. The genetic history of Ice Age Europe. Nature 2016, 534, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Llamas, B.; Willerslev, E.; Orlando, L. Human evolution: A tale from ancient genomes. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20150484. [Google Scholar] [CrossRef] [PubMed]
- Blanga-Kanfi, S.; Miranda, H.; Penn, O.; Pupko, T.; DeBry, R.W.; Huchon, D. Rodent phylogeny revised: Analysis of six nuclear genes from all major rodent clades. BMC Evol. Biol. 2009, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Reed, D.N. Taphonomic implications of roosting behavior and trophic habits in two species of African owl. J. Archaeol. Sci. 2005, 32, 1669–1676. [Google Scholar] [CrossRef]
- Martin, A.P.; Palumbi, S.R. Body size, metabolic rate, generation time, and the molecular clock. Proc. Natl. Acad. Sci. USA 1993, 90, 4087–4091. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Tanimura, M.; Sharp, P.M. An evaluation of the molecular clock hypothesis using mammalian DNA sequences. J. Mol. Evol. 1987, 25, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Bromham, L.; Rambaut, A.; Harvey, P.H. Determinants of rate variation in mammalian DNA sequence evolution. J. Mol. Evol. 1996, 43, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Nabholz, B.; Glémin, S.; Galtier, N. Strong variations of mitochondrial mutation rate across mammals—The longevity hypothesis. Mol. Biol. Evol. 2008, 25, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Orr, H.A. The genetic theory of adaptation: A brief history. Nat. Rev. Genet. 2005, 6, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Bromham, L. Why do species vary in their rate of molecular clock evolution? Biol. Lett. 2009, 5, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Gaines, M.S.; McClenaghan, L.R., Jr. Dispersal in Small Mammals. Annu. Rev. Ecol. Syst. 1980, 11, 163–196. [Google Scholar] [CrossRef]
- Lister, A.M. Mammalian fossils and quaternary biostratigraphy. Quat. Sci. Rev. 1992, 11, 329–344. [Google Scholar] [CrossRef]
- Schreve, D.C. Mammalian evidence from Middle Pleistocene fluvial sequences for complex environmental change at the oxygen isotope substage level. Quat. Int. 2001, 79, 65–74. [Google Scholar] [CrossRef]
- Currant, A.; Jacobi, R. A formal mammalian biostratigraphy for the Late Pleistocene of Britain. Quat. Sci. Rev. 2001, 20, 1707–1716. [Google Scholar] [CrossRef]
- Chaline, J.; Brunet-Lecomte, P.; Montuire, S.; Viriot, L. Anatomy of the arvicoline radiation (Rodentia): Palaeogeographical, palaeoecological history and evolutionary data. Ann. Zool. Fennici 1999, 36, 239–267. [Google Scholar]
- Abramson, N.I.; Lebedev, V.S.; Tesakov, A.S. Bannikova Supraspecies relationships in the subfamily Arvicolinae (Rodentia, Cricetidae): An unexpected result of nuclear gene analysis. Mol. Biol. 2009, 43, 834–846. [Google Scholar] [CrossRef]
- Knapp, M.; Hofreiter, M. Next generation sequencing of ancient DNA: Requirements, strategies and perspectives. Genes 2010, 1, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Enk, J.M.; Devault, A.M.; Kuch, M.; Murgha, Y.E.; Rouillard, J.M.; Poinar, H.N. Ancient whole genome enrichment using baits built from modern DNA. Mol. Biol. Evolut. 2014, 31, 1292–1294. [Google Scholar] [CrossRef] [PubMed]
- Burbano, H.A.; Hodges, E.; Green, R.E.; Briggs, A.W.; Krause, J.; Meyer, M.; Good, J.M.; Maricic, T.; Johnson, P.L.; Xuan, Z.; et al. Targeted investigation of the Neandertal genome by array-based sequence capture. Science 2010, 328, 723–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westbury, M.; Baleka, S.; Barlow, A.; Hartmann, S.; Paijmans, J.L.; Kramarz, A.; Forasiepi, A.M.; Bond, M.; Gelfo, J.N.; Reguero, M.A.; et al. A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nat. Commun. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, B.; Hofreiter, M. A paleogenomic perspective on evolution and gene function: New insights from ancient DNA. Science 2014, 343, 1236573. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, S.; Pruvost, M.; Daligault, J.; Stoetzel, E.; Bennett, E.A.; Cote, N.M.-L.; Nicolas, V.; Lalis, A.; Denys, C.; Geigi, E.-M.; et al. A cost-effective high-throughput metabarcoding approach powerful enough to genotype ~44,000 year-old rodent remains from Northern Africa. Mol. Ecol. Resour. 2016. [Google Scholar] [CrossRef]
- Avery, D. Holocene climatic change in southern Africa: The contribution of micromammals to its study. S. Afr. J. Sci. 1990, 86, 407–412. [Google Scholar]
- Willerslev, E.; Cooper, A. Ancient DNA. Proc. Biol. Sci. 2005, 272, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Kehlmaier, C.; Barlow, A.; Hastings, A.K.; Vamberger, M.; Paijmans, J.L.A.; Steadman, D.W.; Albury, N.A.; Franz, R.; Hofreiter, M.; Fritz, U. Tropical ancient DNA reveals relationships of the extinct Bahamian giant tortoise Chelonoidis alburyorum. Proc. R. Soc. B Biol. Sci. 2016, 21, 1–7. [Google Scholar]
- Gutiérrez-García, T.; Vázquez-Domínguez, E.; Arroyo-Cabrales, J.; Kuch, M.; Enk, J.; King, C.; Poinar, H.N. Ancient DNA and the tropics: A rodent’s tale. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Avenant, N.L. Barn owl pellets: A useful tool for monitoring small mammal communities? Belgian J. Zool. 2005, 135, 39–43. [Google Scholar]
- Smoke, N.D.; Stahl, P.W. Post-burial fragmentation of microinvertebrate skeletons. J. Archaeol. Sci. 2004, 31, 1093–1100. [Google Scholar] [CrossRef]
- Oritz, P.E.; Madozzo Jaén, M.C.; Jayat, J.P. Micromammals and palaeoenvironments: Climatic oscillatons in the Monte desert of Catamarca (Argentina) during the last two millenia. J. Arid Environ. 2012, 77, 103–109. [Google Scholar] [CrossRef]
- McCrae, C. A comparative study of Late Holocene and Plio-Pleistocene-aged micromammalian owl accumulations from South Africa. Palaeontol. Afr. 2009, 44, 190–191. [Google Scholar]
- Taberlet, P.; Fumagalli, L. Owl pellets as a source of DNA for genetic studies of small mammals. Mol. Ecol. 1996, 5, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Poulakakis, N.; Lymberakis, P.; Paragamian, K.; Mylonas, M. Isolation and amplification of shrew DNA from barn owl pellets. Biol. J. Linn. Soc. 2005, 85, 331–340. [Google Scholar] [CrossRef]
- Guimaraes, S.; Fernandez-Jalvo, Y.; Stoetzel, E.; Gorgé, O.; Bennett, E.A.; Denys, C.; Grange, T.; Geigl, E.M. Owl pellets: A wise DNA source for small mammal genetics. J. Zool. 2016, 298, 64–74. [Google Scholar] [CrossRef]
- Brace, S.; Thomas, J.A.; Dalén, L.; Burger, J.; MacPhee, R.D.; Barnes, I.; Turvey, S.T. Evolutionary history of the Nesophontidae, the last unplaced Recent mammal family. Mol. Biol. Evol. 2016, 33, 3095–3103. [Google Scholar] [CrossRef] [PubMed]
- Pagès, M.; Chevret, P.; Gros-Balthazard, M.; Hughes, S.; Alcover, J.A.; Hutterer, R.; Rando, J.C.; Michaux, J.; Hänni, C. Paleogenetic analyses reveal unsuspected phylogenetic affinities between mice and the extinct Malpaisomys insularis, an endemic rodent of the Canaries. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, M.A.J. Middle Pleistocene Smaller Mammals from the Monte Peglia (Orvieto, Italy) with Special Reference to the Phylogeny of Microtus (Arvicolidae, Rodentia). Quaternaria 1973, 17, 1–144. [Google Scholar]
- Avery, D.M. An assessment of the Lower Pleistocene micromammalian fauna from Swartkrans members 1–3, Gauteng, South Africa. Geobios 1998, 31, 393–414. [Google Scholar] [CrossRef]
- Matthews, T.; Stynder, D.D. An analysis of two Myosorex species (Soricidae) from the Early Pliocene site of Langebaanweg (West coast, South Africa) using geometric morphometrics, linear measurements, and non-metric characters. Geobios 2011, 44, 87–99. [Google Scholar] [CrossRef]
- Adams, D.C.; Otárola-Castillo, E. Geomorph: An r package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Cornette, R.; Herrel, A.; Stoetzel, E.; Moulin, S.; Hutterer, R.; Denys, C.; Baylac, M. Specific information levels in relation to fragmentation patterns of shrew mandibles: Do fragments tell the same story? J. Archaeol. Sci. 2015, 53, 323–330. [Google Scholar] [CrossRef]
- Churakov, G.; Sadasivuni, M.K.; Rosenbloom, K.R.; Huchon, D.; Brosius, J.; Schmitz, J. Rodent evolution: Back to the root. Mol. Biol. Evol. 2010, 27, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Markova, E.; Beeren, Z.; van Kolfschoten, T.; Strukova, T.; Vrieling, K. Differentiating sibling species in the Quaternary fossil record: A comparison of morphological and molecular methods to identify Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia). J. Syst. Palaeontol. 2012, 10, 585–597. [Google Scholar] [CrossRef]
- Fumagalli, L.; Taberlet, P.; Stewart, D.T.; Gielly, L.; Hausser, J.; Vogel, P. Molecular phylogeny and evolution of Sorex shrews (Soricidae: Insectivora) inferred from mitochondrial DNA sequence data. Mol. Phylogenet. Evol. 1999, 11, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Chaline, J.; Graf, J.D. Phylogeny of the Arvicolidae (Rodentia): Biochemical and paleontological evidence. J. Mammal. 1988, 69, 22–33. [Google Scholar] [CrossRef]
- Hofreiter, M.; Paijmans, J.L.A.; Goodchild, H.; Speller, C.F.; Barlow, A.; Fortes, G.G.; Thomas, J.A.; Ludwig, A.; Collins, M.J. The future of ancient DNA: Technical advances and conceptual shifts. Bioessays 2015, 37, 284–293. [Google Scholar]
- Tegelström, H. Transfer of mitochondrial DNA from the northern red-backed vole (Clethrionomys rutilus) to the bank vole (C. glareolus). J. Mol. Evol. 1987, 24, 218–227. [Google Scholar]
- Runck, A.; Matocq, M.; Cook, J. Historic hybridization and persistence of a novel mito-nuclear combination in red-backed voles (genus Myodes). BMC Evol. Biol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Ballard, J.W.; Whitlock, M.C. The incomplete natural history of mitochondria. Mol. Ecol. 2004, 13, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Haring, E.; Voyta, L.L.; Däubl, B.; Tiunov, M.P. Comparison of genetic and morphological characters in fossil teeth of grey voles from the Russian Far East (Rodentia: Cricetidae: Alexandromys). Mamm. Biol. 2015, 80, 496–504. [Google Scholar] [CrossRef]
- Rodrigues, M.; Bos, A.R.; Hoath, R.; Schembri, P.J.; Lymberakis, P.; Cento, M.; Ghawar, W.; Ozkurt, S.O.; Santos-Reis, M.; Merilä, J.; et al. Taxonomic status and origin of the Egyptian weasel (Mustela subpalmata) inferred from mitochondrial DNA. Genetica 2016, 144, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Amori, G.; Gippoliti, S.; Helgen, K.M. Diversity, distribution, and conservation of endemic island rodents. Quat. Int. 2008, 182, 6–15. [Google Scholar] [CrossRef]
- Ricklefs, R.; Bermingham, E. The West Indies as a laboratory of biogeography and evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2393–2413. [Google Scholar] [CrossRef] [PubMed]
- Brace, S.; Turvey, S.T.; Weksler, M.; Hoogland, M.L.P.; Barnes, I. Unexpected evolutionary diversity in a recently extinct Caribbean mammal radiation. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Fabre, P.H.; Vilstrup, J.T.; Raghavan, M.; der Sarkissian, C.; Willerslev, E.; Douzery, E.J.P.; Orlando, L. Rodents of the Caribbean: Origin and diversification of hutias unravelled by next-generation museomics. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S.S.; Matisoo-Smith, E.; Hunt, T.L. Ancient DNA of the Pacific rat (Rattus exulans) from Rapa Nui (Easter Island). J. Archaeol. Sci. 2006, 33, 1536–1540. [Google Scholar] [CrossRef]
- Matisoo-Smith, E.; Sutton, D.G.; Ladefoged, T.N.; Lambert, D.M.; Allen, J.S. Prehistoric mobility in Polynesia: MtDNA variation in Rattus exulans from the Chatham and Kermadec Islands. Asian Perspect. 1999, 38, 186–199. [Google Scholar]
- Wyatt, K.B.; Campos, P.F.; Gilbert, M.T.P.; Kolokotronis, S.; Hynes, W.H.; DeSalle, R.; Daszak, P.; MacPhee, R.D.E.; Greenwood, A.D. Historical mammal extinction on Christmas Island (Indian Ocean) correlates with introduced infectious disease. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Martinkova, N.; Barnett, R.; Cucchi, T.; Struchen, R.; Pascal, M.; Pascal, M.; Fischer, M.C.; Higham, T.; Brace, S.; Simon, Y.W.H.; et al. Divergent evolutionary processes associated with colonization of offshore islands. Mol. Ecol. 2013, 22, 5205–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickrell, J.K.; Reich, D. Toward a new history and geography of human genes informed by ancient DNA. Trends Genet. 2014, 30, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Castle, W.E. The domestication of the rat. Proc. Natl. Acad. Sci. USA 1947, 33, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.; Skirnisson, K.; McGovern, T.H.; Gilbert, M.T.; Willerslev, E.; Searle, J.B. Fellow travellers: A concordance of colonization patterns between mice and men in the North Atlantic region. BMC Evol. Biol. 2012, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Clout, M.N.; Russell, J.C. The invasion ecology of mammals: A global perspective. Wildl. Res. 2008, 35, 180–184. [Google Scholar] [CrossRef]
- Withers, P.; Cooper, E.; Maloney, K.; Bozinovic, F.; Cruz, A. Introduction to mammals. In Ecological and Enviromental Physiology of Mammals; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Hadly, E.A.; Ramakrishnan, U.M.A.; Chan, Y.L.; Van Tuinen, M.; O’Keefe, K.; Spaeth, P.A.; Conroy, C.J. Genetic response to climatic change: Insights from ancient DNA and phylochronology. PLoS Biol. 2004, 2, e290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaumont, M.A.; Zhang, W.; Balding, D.J. Approximate Bayesian computation in population genetics. Genetics 2002, 162, 2025–2035. [Google Scholar] [PubMed]
- Hadly, E.A.; van Tuinen, M.; Chan, Y.; Heiman, K. Ancient DNA evidence of prolonged population persistence with negligible genetic diversity in an endemic Tuco-Tuco (Ctenomys sociabilis). J. Mammal. 2003, 84, 403–417. [Google Scholar] [CrossRef]
- Chan, Y.L.; Lacey, E.A.; Pearson, O.P.; Hadly, E.A. Ancient DNA reveals Holocene loss of genetic diversity in a South American rodent. Biol. Lett. 2005, 1, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Hofman, C.A.; Rick, T.C.; Fleischer, R.C.; Maldonado, J.E. Conservation archaeogenomics: Ancient DNA and biodiversity in the Anthropocene. Trends Ecol. Evol. 2015, 30, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Prost, S.; Smirnov, N.; Fedorov, V.B.; Sommer, R.S.; Stiller, M.; Nagel, D.; Knapp, M.; Hofreiter, M. Influence of climate warming on arctic mammals? new insights from ancient DNA studies of the collared lemming Dicrostonyx torquatus. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Jaarola, M.; Martínková, N.; Gündüz, İ.; Brunhoff, C.; Zima, J.; Nadachowski, A.; Amori, G.; Bulatova, N.S.; Chondropoulos, B.; Fraguedakis-Tsolis, S.; et al. Molecular phylogeny of the speciose vole genus Microtus (Arvicolinae, Rodentia) inferred from mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2004, 33, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Catania, K.C. Evolution of sensory specializations in insectivores. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1038–1050. [Google Scholar] [CrossRef] [PubMed]
- Brace, S.; Barnes, I.; Powell, A.; Pearson, R.; Woolaver, L.G.; Thomas, M.G.; Turvey, S.T. Population history of the Hispaniolan hutia Plagiodontia aedium (Rodentia: Capromyidae): Testing the model of ancient differentiation on a geotectonically complex Caribbean island. Mol. Ecol. 2012, 21, 2239–2253. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, L.D.; Lambert, D.M. Ancient DNA and conservation: Lessons from the endangered kiwi of New Zealand. Mol. Ecol. 2008, 17, 2174–2184. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Vanderpool, D.; Singhal, S.; Linderoth, T.; Moritz, C.; Good, J.M. Transcriptome-based exon capture enables highly cost-effective comparative genomic data collection at moderate evolutionary scales. BMC Genom. 2012, 13, 403. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.; Linderoth, T.; Vanderpool, D.; Good, J.M.; Nielsen, R.; Moritz, C. Unlocking the vault: Next-generation museum population genomics. Mol. Ecol. 2013, 22, 6018–6032. [Google Scholar] [CrossRef] [PubMed]
- Smulders, M.J.M.; Snoek, L.B.; Booy, G.; Vosman, B. Complete loss of MHC genetic diversity in the Common Hamster (Cricetus cricetus) population in The Netherlands. Consequences for conservation strategies. Conserv. Genet. 2003, 4, 441–451. [Google Scholar] [CrossRef]
- Provan, J.; Bennett, K.D. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2008, 23, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Brace, S.; Palkopoulou, E.; Dalén, L.; Lister, A.M.; Miller, R.; Otte, M.; Germonpré, M.; Blockley, S.P.; Stewart, J.R.; Barnes, I. Serial population extinctions in a small mammal indicate Late Pleistocene ecosystem instability. Proc. Natl. Acad. Sci. USA 2012, 109, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Ammerman, A.J.; Luigi, L.C. The Neolithic Transition and the Genetics of Populations in Europe; Princeton University Press: Princeton, NJ, USA, 2014. [Google Scholar]
- Thomas, W.K.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Spatial and temporal continuity of kangaroo rat populations shown by sequencing mitochondrial DNA from museum specimens. J. Mol. Evol. 1990, 31, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Matisoo-Smith, E.; Allen, J.S.; Ladefoged, T.N.; Roberts, R.; Lambert, D.M. Ancient DNA from Polynesian rats: Extraction, amplification and sequence from single small bones. Electrophoresis 1997, 18, 1534–1537. [Google Scholar] [CrossRef] [PubMed]
- Matisoo-Smith, E.; Robins, J.H. Origins and dispersals of Pacific peoples: Evidence from mtDNA phylogenies of the Pacific rat. Proc. Natl. Acad. Sci. USA 2004, 101, 9167–9172. [Google Scholar] [CrossRef] [PubMed]
- Matisoo-Smith, E.; Robins, J. Mitochondrial DNA evidence for the spread of Pacific rats through Oceania. Biol. Invasions 2009, 11, 1521–1527. [Google Scholar] [CrossRef]
- Robins, J.H.; Tintinger, V.; Aplin, K.P.; Hingston, M.; Matisoo-Smith, E.; Penny, D.; Lavery, S.D. Phylogenetic species identification in Rattus highlights rapid radiation and morphological similarity of New Guinean species. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Orlando, L.; Mauffrey, J.F.; Cuisin, J.; Patton, J.L.; Hänni, C.; Catzeflis, F. Napoleon Bonaparte and the fate of an amazonian rat: New data on the taxonomy of Mesomys hispidus (Rodentia: Echimyidae). Mol. Phylogenet. Evol. 2003, 27, 113–120. [Google Scholar] [CrossRef]
- Demastes, J.W.; Butt, A.L.; Hafner, M.S.; Light, J.E. Systematics of a rare species of pocket gopher, Pappogeomys alcorni. J. Mammal. 2003, 84, 753–761. [Google Scholar] [CrossRef]
- Spaeth, P.A.; van Tuinen, M.; Chan, Y.L.; Terca, D.; Hadly, E.A. Phylogeography of Microtus. Longicaudus. in the Tectonically and Glacially Dynamic Central Rocky Mountains. J. Mammal. 2009, 90, 571–584. [Google Scholar] [CrossRef]
- Prost, S.; Guralnick, R.P.; Waltari, E.; Fedorov, V.B.; Kuzmina, E.; Smirnov, N.; Kolfschoten, T.; Hofreiter, M.; Vrieling, K. Losing ground: Past history and future fate of Arctic small mammals in a changing climate. Glob. Chang. Biol. 2013, 19, 1854–1864. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.L.; Hadly, E.A. Genetic variation over 10,000 years in Ctenomys: Comparative phylochronology provides a temporal perspective on rarity, environmental change and demography. Mol. Ecol. 2011, 20, 4592–4605. [Google Scholar] [CrossRef] [PubMed]
- Krystufek, B.; Buzan, E.V.; Hutchinson, W.F.; Hänfling, B. Phylogeography of the rare Balkan endemic Martino’s vole, Dinaromys bogdanovi, reveals strong differentiation within the western Balkan Peninsula. Mol. Ecol. 2007, 16, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Kerhoulas, N.J.; Arbogast, B.S. Molecular systematics and Pleistocene biogeography of Mesoamerican flying squirrels. J. Mammal. 2010, 91, 654–667. [Google Scholar] [CrossRef]
- Fulton, T.L.; Norris, R.W.; Graham, R.W.; Semken, H.A.; Shapiro, B. Ancient DNA supports southern survival of Richardson’s collared lemming (Dicrostonyx richardsoni) during the last glacial maximum. Mol. Ecol. 2013, 22, 2540–2548. [Google Scholar] [CrossRef] [PubMed]
- Palkopoulou, E.; Baca, M.; Abramson, N.I.; Sablin, M.; Socha, P.; Nadachowski, A.; Prost, S.; Germonpré, M.; Kosintsev, P.; Smirnov, N.G.; et al. Synchronous genetic turnovers across Western Eurasia in Late Pleistocene collared lemmings. Glob. Chang. Biol. 2016, 22, 1710–1721. [Google Scholar] [CrossRef] [PubMed]
- Kalkvik, H.M.; Stout, I.J.; Doonan, T.J.; Parkinson, C.L. Investigating niche and lineage diversification in widely distributed taxa: Phylogeography and ecological niche modeling of the Peromyscus maniculatus species group. Ecography 2012, 35, 54–64. [Google Scholar] [CrossRef]
- Brace, S.; Ruddy, M.; Miller, R.; Schreve, D.C.; Stewart, J.R.; Barnes, I. The colonization history of British water vole (Arvicola amphibius (Linnaeus, 1758)): Origins and development of the Celtic fringe. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160130. [Google Scholar] [CrossRef] [PubMed]
- Kuch, M.; Rohland, N.; Betancourt, J.L.; Latorre, C.; Steppan, S.; Poinar, H.N. Molecular analysis of an 11,700-year-old rodent midden from the Atacama Desert, Chile. Mol. Ecol. 2002, 11, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Pacioni, C.; Hunt, H.; Allentoft, M.E.; Vaughan, T.G.; Wayne, A.F.; Baynes, A.; Haouchar, D.; Dortch, J.; Bunce, M. Genetic diversity loss in a biodiversity hotspot: Ancient DNA quantifies genetic decline and former connectivity in a critically endangered marsupial. Mol. Ecol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Prost, S.; Klietmann, J.; van Kolfschoten, T.; Guralnick, R.; Waltari, E.; Vrieling, K.; Stiller, M.; Nagel, D.; Rabeder, G.; Hofreiter, M.; et al. Effects of late quaternary climate change on Palearctic shrews. Glob. Chang. Biol. 2013, 19, 1865–1874. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.; Sten, S.; Götherström, A. Neolithic Hedgehogs (Erinaceus europaeus) from the Island of Gotland show early contacts with the Swedish mainland. J. Archaeol. Sci. 2012, 39, 229–233. [Google Scholar] [CrossRef]
- Tougard, C.; Renvoisé, E. Rodents and palaeogenetics: New perspectives. Comptes Rendus Palevol 2008, 7, 125–134. [Google Scholar] [CrossRef]
Figure 1.
Number of ancient DNA (aDNA) studies including micromammals from 1990–2017.

Figure 2.
aDNA studies including micromammals between 1990–2015.


{kind=link}
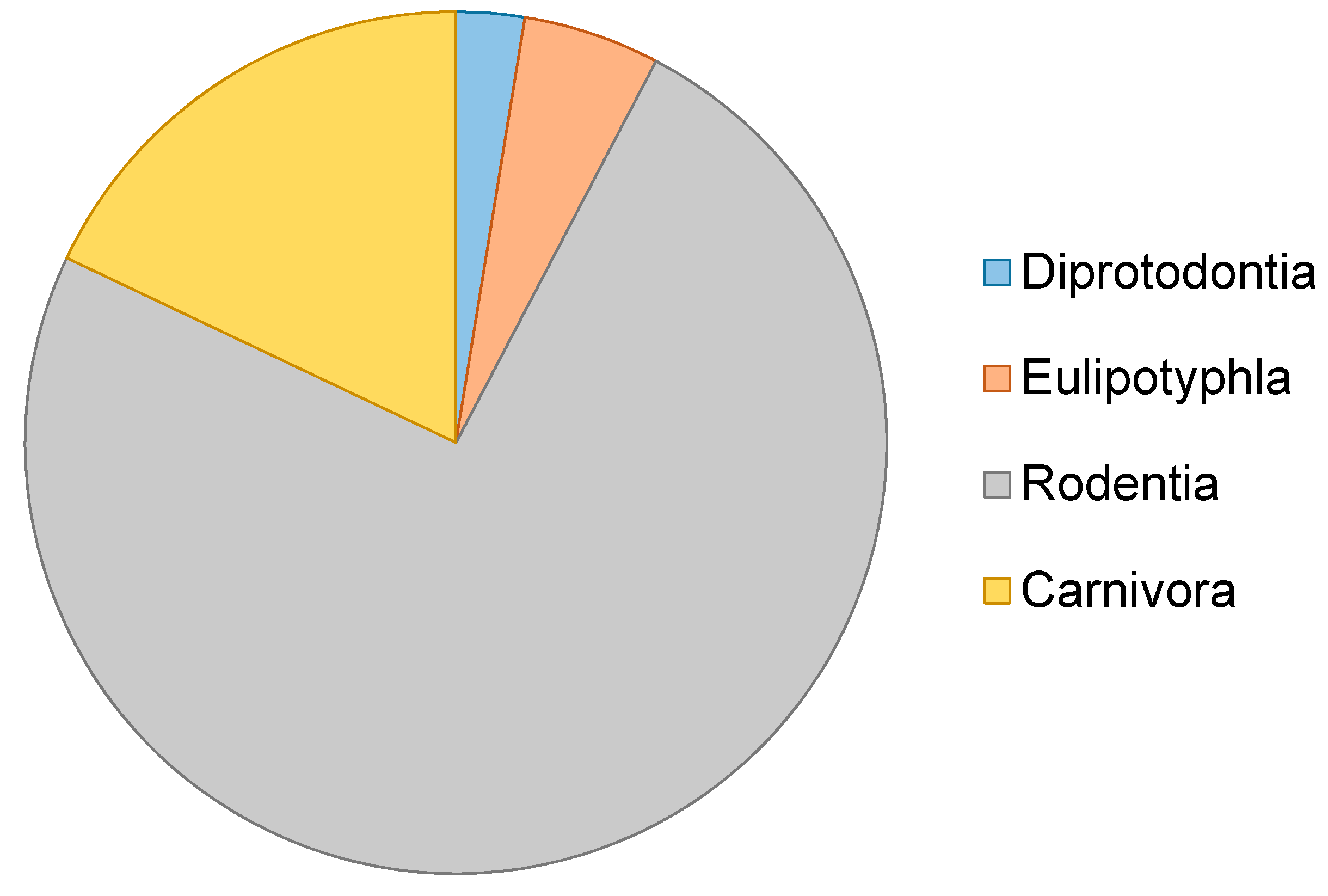{kind=link}
Table 1.
Published studies from 1990 to 2017 in which ancient or degraded DNA was utilised to study micromammal fauna.
Table 1.
Published studies from 1990 to 2017 in which ancient or degraded DNA was utilised to study micromammal fauna.
| Order | Genus | Species | Date | Author | Genetic Marker |
|---|---|---|---|---|---|
| Rodentia | Dipodomys | panamintinus | 1990 | Thomas et al. [93] | Control Region |
| Rodentia | Rattus | exulans | 1999 | Matisoo-Smith et al. [69] | Control Region |
| Rodentia | Rattus | exulans | 1997 | Matisoo-Smith et al. [94] | Control Region |
| Rodentia | Rattus | exulans | 2004 | Matisoo-Smith et al. [95] | mtDNA (~240 bp D-loop) |
| Rodentia | Rattus | exulans | 2006 | Barnes et al. [68] | Control Region |
| Rodentia | Rattus | exulans | 2009 | Matisoo-Smith & Robins [96] | Control region, Cytochrome b, Cytochrome Oxidase Subunit I |
| Rodentia | Rattus | - | 2014 | Robins et al. [97] | Control region, Cytochrome Oxidase Subunit I |
| Rodentia | Mesomys | hispidus | 2003 | Orlando et al. [98] | Cytochrome b |
| Rodentia | Pappogeomys | alcorni | 2003 | Demastes et al. [99] | Cytochrome b |
| Rodentia | Megalomys | georginae, desmarestii | 2015 | Brace et al. [66] | Cytochrome b |
| Rodentia | Microtus | montanus | 2004 | Hadly et al. [77] | Cytochrome b |
| Rodentia | Microtus | longicaudus | 2009 | Spaeth et al. [100] | Cytochrome b |
| Rodentia | Microtus | arvalis, rossiaemeridionalis | 2012 | Markova et al. [55] | Cytochrome b |
| Rodentia | Microtus | gregalis | 2013 | Prost et al. [101] | Cytochrome b |
| Rodentia | Ctenomys | sociabilis | 2003 | Hadly et al. [79] | |
| Rodentia | Ctenomys | sociabilis | 2005 | Chan et al. [80] | Cytochrome b |
| Rodentia | Ctenomys | sociabilis | 2006 | Chan et al. [72] | Cytochrome b |
| Rodentia | Ctenomys | haigi, sociabilis | 2011 | Chan & Hadley [102] | Cytochrome b |
| Rodentia | Dinaromys | bogdanovi | 2007 | Krystufek et al. [103] | Cytochrome b |
| Rodentia | Glaucomys | volans | 2010 | Kerhoulas & Arbogast [104] | Cytochrome b |
| Rodentia | Dicrostonyx | torquatus | 2010 | Prost et al. [82] | Control Region |
| Rodentia | Dicrostonyx | torquatus | 2012 | Brace et al. [91] | Cytochrome b |
| Rodentia | Dicrostonyx | richardsoni | 2013 | Fulton et al. [105] | Cytochrome b |
| Rodentia | Dicrostonyx | torquatus | 2013 | Prost et al. [101] | Cytochrome b |
| Rodentia | Dicrostonyx | groenlandicus, richardsoni, torquatus | 2016 | Palkopoulou et al. [106] | Cytochrome b |
| Rodentia | Peromyscus | polionotus | 2012 | Kalkvik et al. [107] | Cytochrome b |
| Rodentia | Plagiodontia | aedium | 2012 | Brace et al. [85] | Cytochrome b |
| Rodentia | Ototylomus | phyllotis | 2014 | Gutierrez-Garcia et al. [39] | Cytochrome b |
| Rodentia | Alexandromys | fortis, indet, maximowiczii, oeconomus | 2015 | Haring et al. [62] | Control Region |
| Rodentia | Pennatomys | nivalis, luciae | 2015 | Brace et al. [66] | Cytochrome b |
| Rodentia | Arvicola | amphibius | 2016 | Brace et al. [108] | Control Region |
| Rodentia | Malpaisomys, Mus, Murinae | - | 2012 | Pages et al. [48] | Cytochrome b, Interphotoreceptor Retinoid Binding Protein (IRBP) |
| Rodentia | Mus, Meriones, Apodemus, Lemniscomys, Rattus, Gerbillus, Cricetulus, Microtus, Eliomys | - | 2016 | Guimaraes et al. [35] | Single Nucleotide Polymorphisms (SNPs) |
| Rodentia | - | - | 2002 | Kuch et al. [109] | 12S & 16S rDNA, Cytochrome b |
| Diprotodontia | Bettongia | penicillata | 2015 | Pacioni et al. [110] | Control Region, Microsatellites |
| Eulipotyphla | Sorex | araneus, tundrensis | 2013 | Prost et al. [111] | Cytochrome b |
| Eulipotyphla | Erinaceus | europaeus | 2012 | Fraser et al. [112] | Cytochrome b |
| Eulipotyphla | Nesophontes | paramicrus | 2016 | Brace et al. [47] | Mitochondrial genome, ADORA3, ADRA2B, ADRB2, APOB, APP, ATP7A, BCHE, BDNF, BMI1, BMP4, BRCA1, CREM, EDG1, GHR, PLCB4, RAG1, RAG2, RHO, TTN, TYR, VWF |
| - | - | - | 2005 | Avenant et al. [40] | Cytochrome b |
| - | - | - | 2005 | Poulakakis et al. [45] | Cytochrome b |
| - | - | - | 2016 | Guimaraes et al. [46] | MtDNA |
| - | - | - | 2008 | Tougard & Renvoise [113] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Woods, R.; Marr, M.M.; Brace, S.; Barnes, I. The Small and the Dead: A Review of Ancient DNA Studies Analysing Micromammal Species. Genes 2017, 8, 312. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110312
AMA Style
Woods R, Marr MM, Brace S, Barnes I. The Small and the Dead: A Review of Ancient DNA Studies Analysing Micromammal Species. Genes. 2017; 8(11):312. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110312
Chicago/Turabian StyleWoods, Roseina, Melissa M. Marr, Selina Brace, and Ian Barnes. 2017. "The Small and the Dead: A Review of Ancient DNA Studies Analysing Micromammal Species" Genes 8, no. 11: 312. https://0-doi-org.brum.beds.ac.uk/10.3390/genes8110312
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.