Distinct Roles of Velvet Complex in the Development, Stress Tolerance, and Secondary Metabolism in Pestalotiopsis microspora, a Taxol Producer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal and Bacterial Strains, Culture Conditions
2.2. Construction of Deletion Vector
2.3. Disruption and Restoration of veA, velB and laeA by Agrobacterium Tumefaciens-Mediated Genetic Transformation
2.4. Characterization of Deletion and Restoration Strains
2.5. RNA Preparation, Reverse-Transcription Polymerase Chain Reaction , and Quantitative Real-Time Polymerase Chain Reaction.
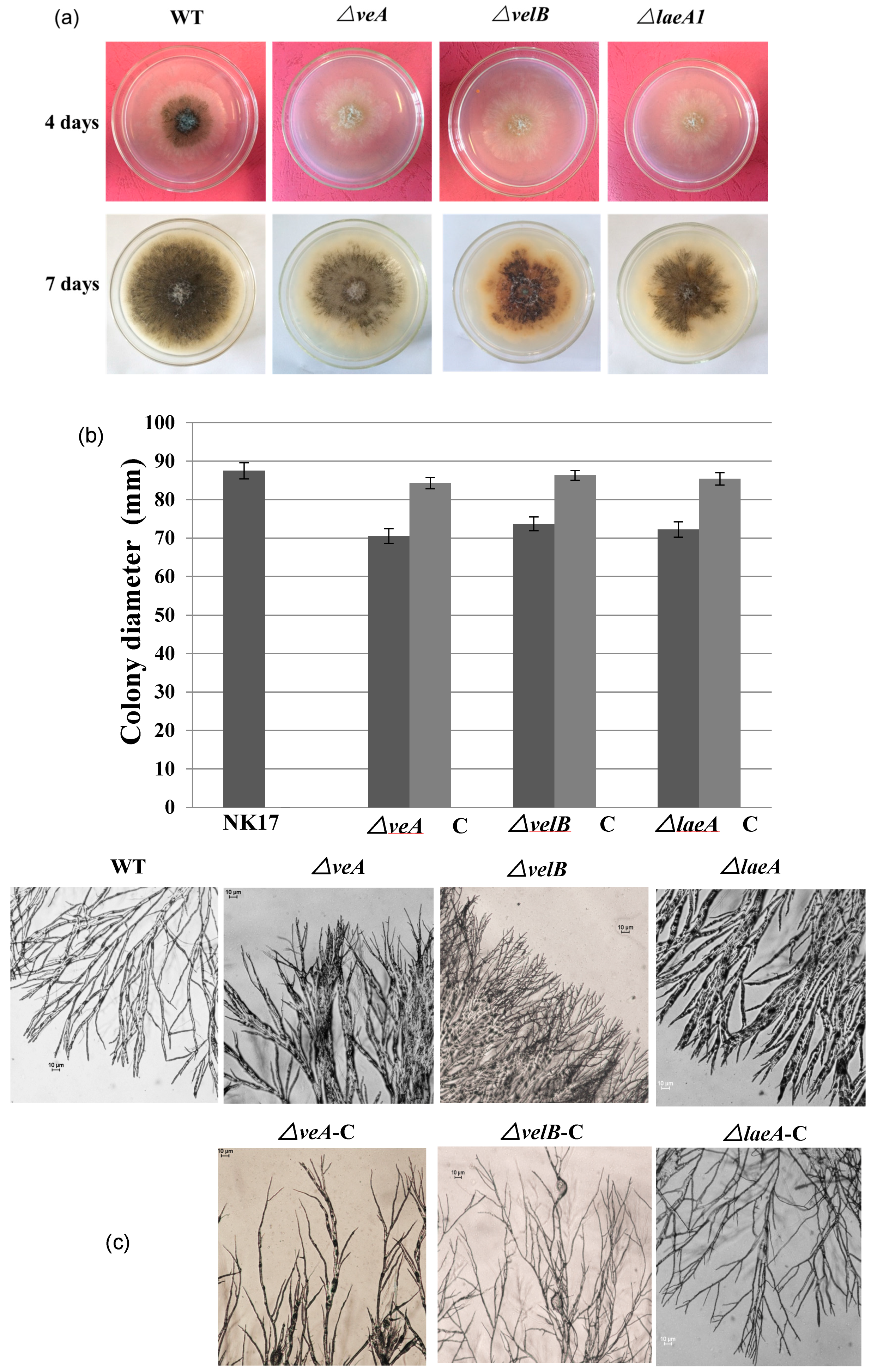
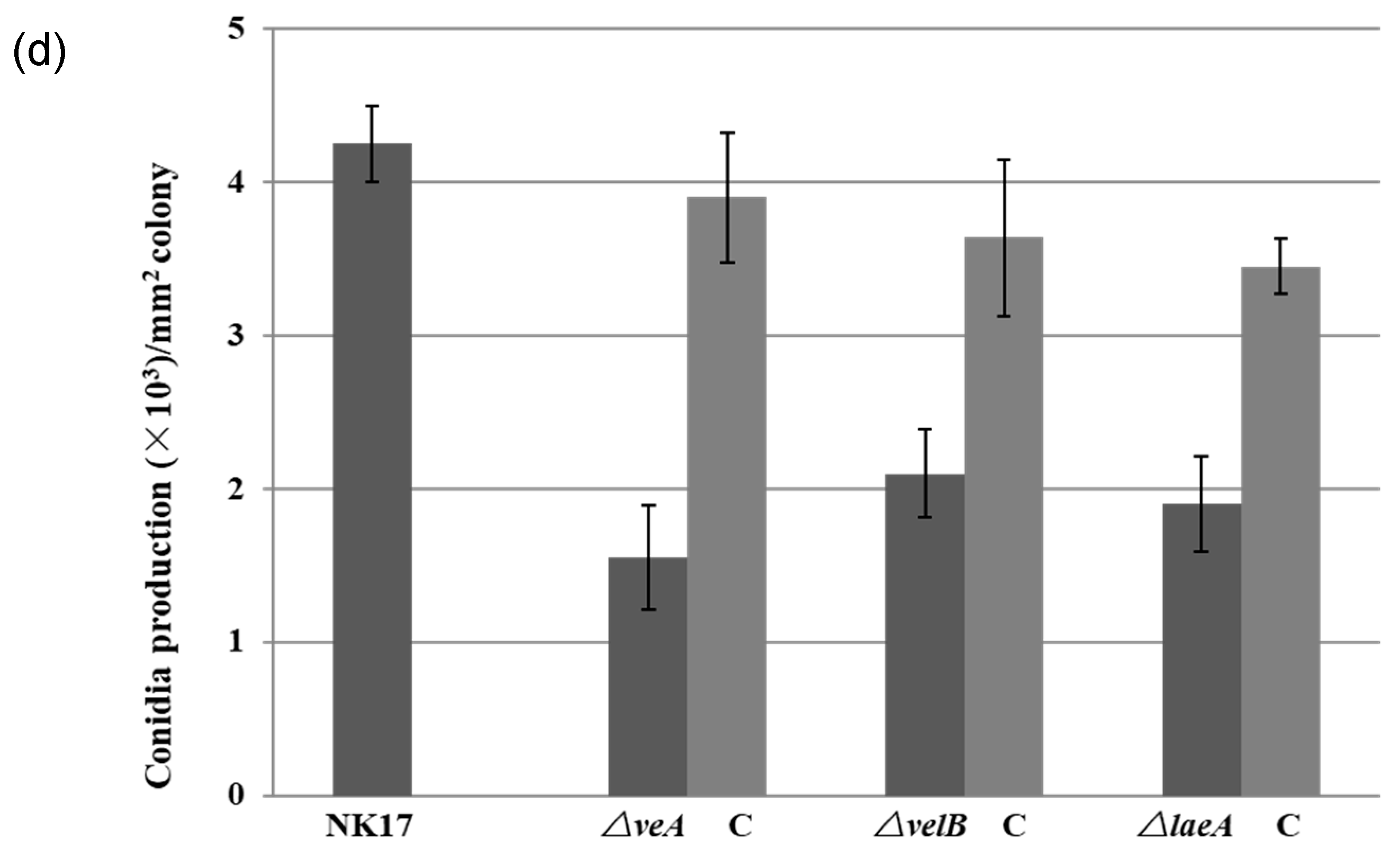
2.6. Mycelial Growth and Sporulation Assessment
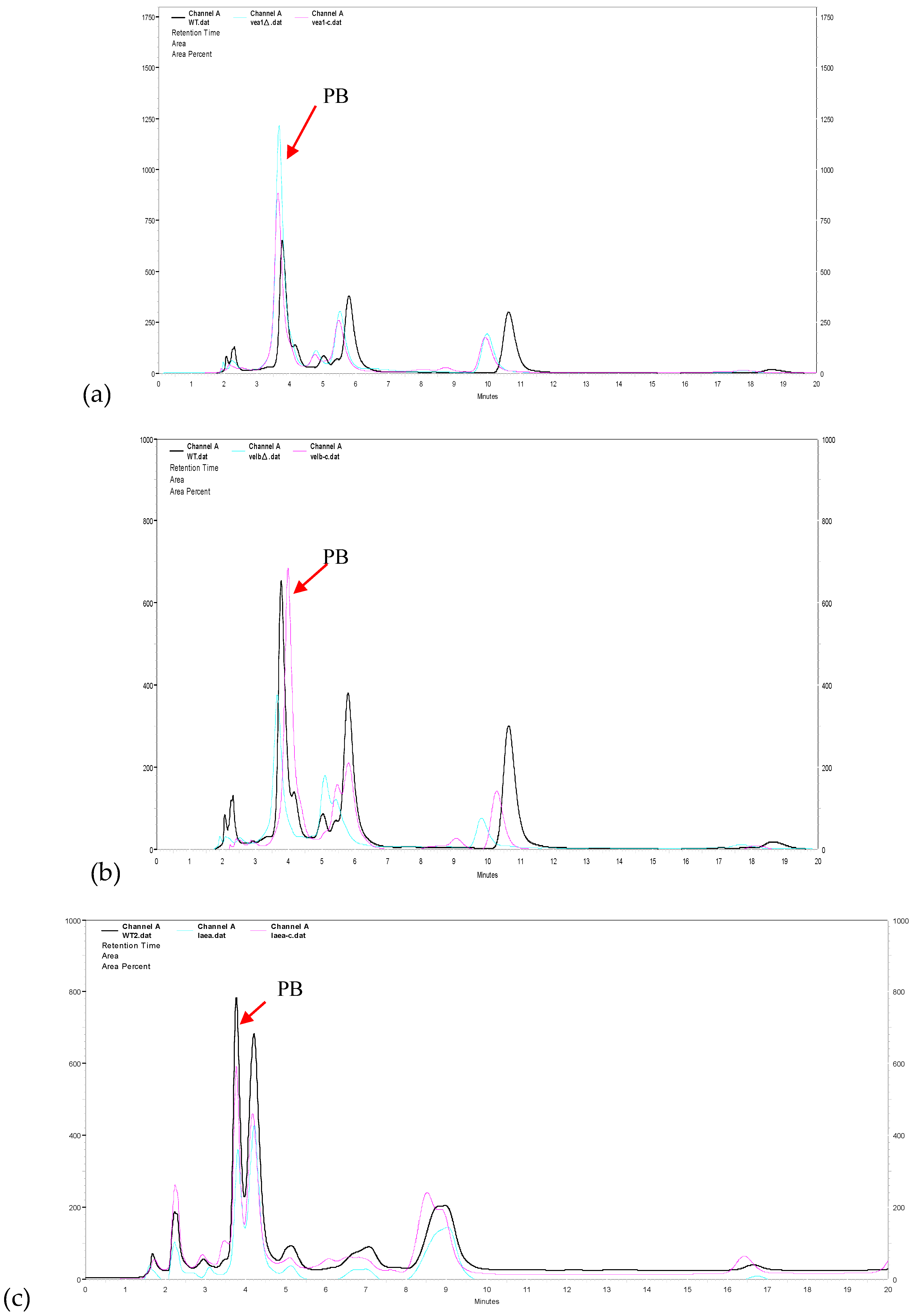
2.7. Profiling of Secondary Metabolites by High-Performance Liquid Chromatography.
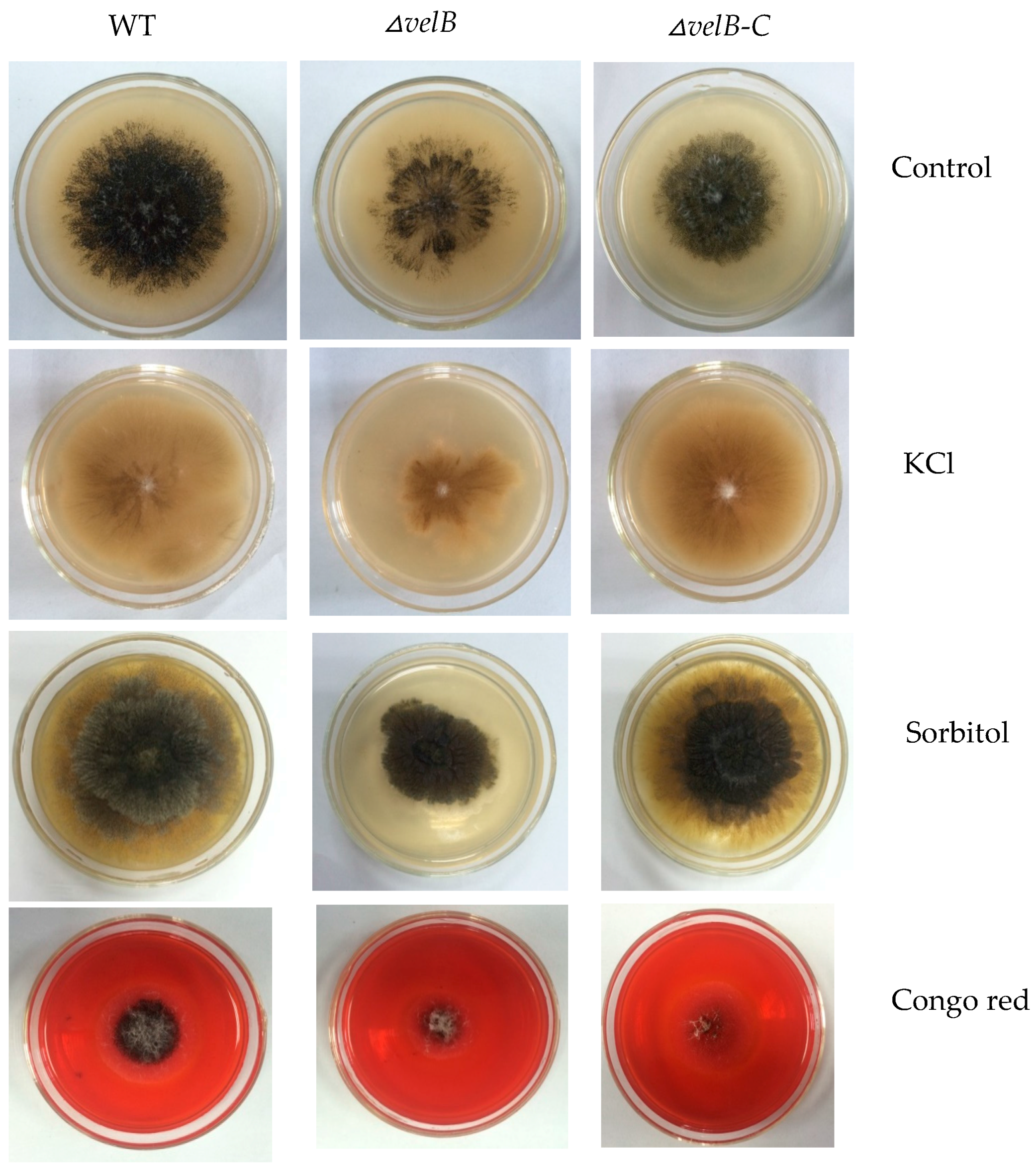
2.8. Stress Response Assays
3. Results
3.1. Identification of the VeA, VelB, and LaeA Orthologs in Pestalotiopsis microspora
3.2. Involvement of VeA, VelB, and LaeA in Hyphal Growth and Conidiation
3.3. Determination of Sensitivity of the Mutants to External Stresses
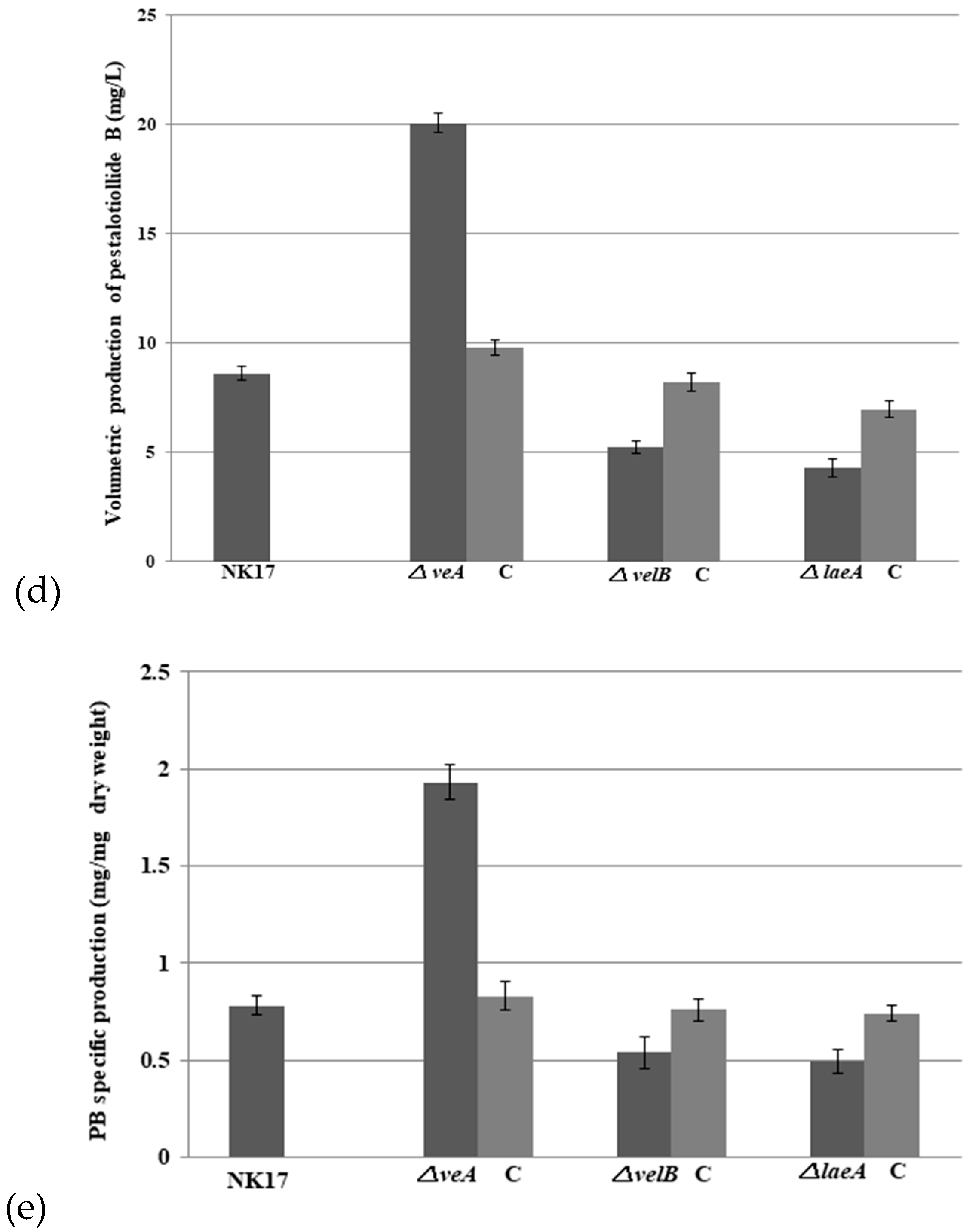
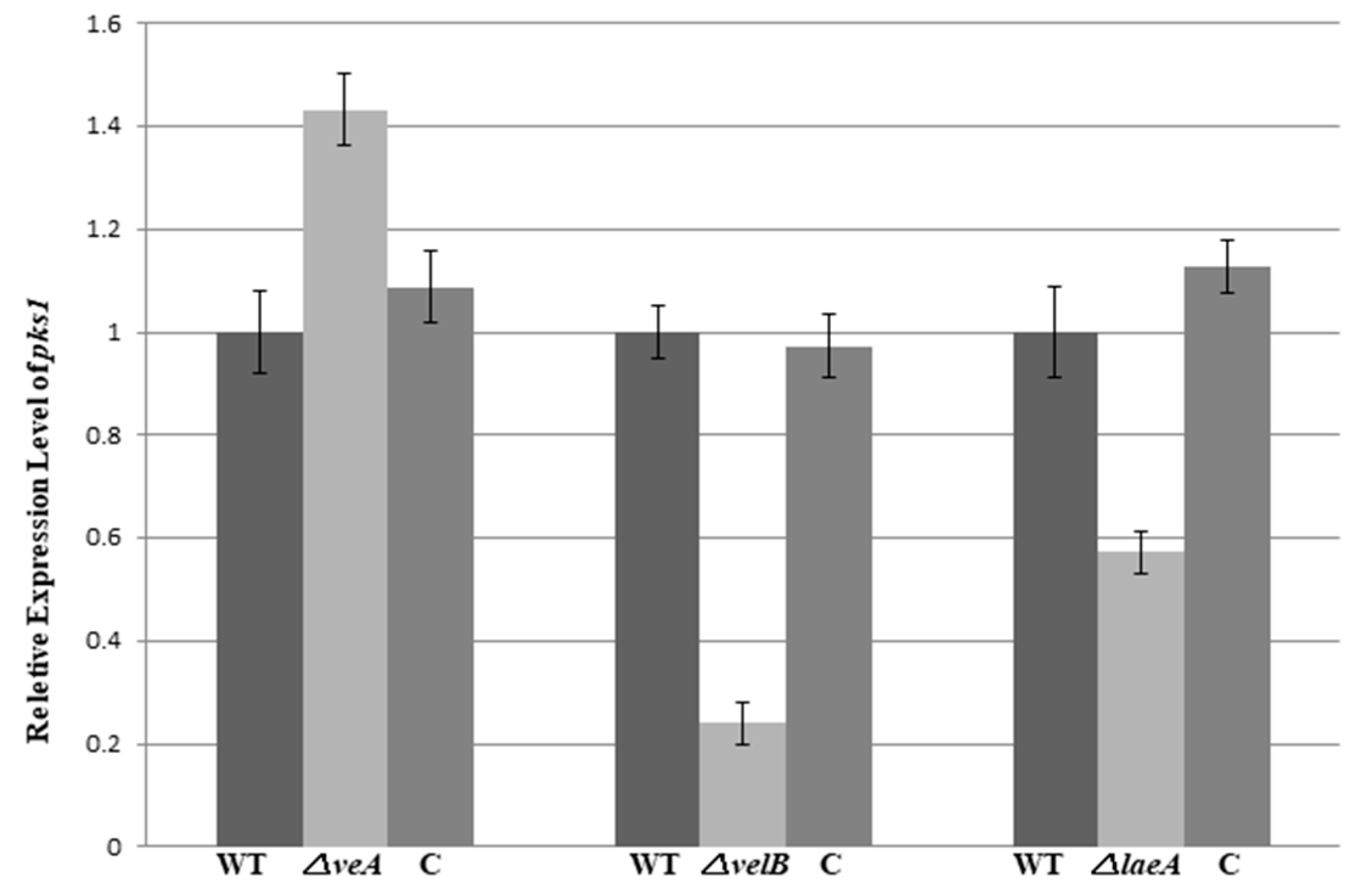
3.4. VeA, VelB, and LaeA Regulate Biosynthesis of Pestalotiollide B and Mycelial Pigments
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maharachchikumbura, S.S.; Guo, L.D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Strobel, G.; Yang, X.; Sears, J.; Kramer, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Ford, E.; Worapong, J.; Harper, J.K.; Arif, A.M.; Grant, D.M.; Fung, P.C.; Chau, M.W. Isopestacin, an isobenzofuranone from Pestalotiopsis microspora, possessing antifungal and antioxidant activities. Phytochemistry 2002, 60, 179–183. [Google Scholar] [CrossRef]
- Ding, G.; Liu, S.; Guo, L.; Zhou, Y.; Che, Y. Antifungal metabolites from the plant endophytic fungus Pestalotiopsis foedan. J. Nat. Prod. 2008, 71, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Jiang, L.; Guo, L.; Chen, X.; Zhang, H.; Che, Y. Pestalazines and pestalamides, bioactive metabolites from the plant pathogenic fungus Pestalotiopsis theae. J. Nat. Prod. 2008, 71, 1861–1865. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Y.L.; Gerke, J.; Park, H.S.; Bayram, Ö.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.C.; Yu, J.H.; Braus, G.H.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-ĸB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Han, K.Y.; Kim, K.J.; Han, D.M.; Jahng, K.Y.; Chae, K.S. The veA gene activates sexual development in Aspergillus nidulans. Fungal Genet. Biol. 2002, 37, 72–80. [Google Scholar] [CrossRef]
- Chettri, P.; Bradshaw, R.E. LaeA negatively regulates dothistromin production in the pine needle pathogen Dothistroma septosporum. Fungal Genet. Biol. 2016, 97, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Brooks, W.; Calvo, A.M. The expression of sterigmatocystin and penicillin genes in Aspergillus nidulans is controlled by veA, a gene required for sexual development. Eukaryot. Cell 2003, 2, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Spröte, P.; Brakhage, A.A. The light-dependent regulator velvet A of Aspergillus nidulans acts as a repressor of the penicillin biosynthesis. Arch. Microbiol. 2007, 188, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Brown, D.W.; Kleigrewe, K.; Bok, J.W.; Keller, N.P.; Humpf, H.U.; Tudzynski, B. FfVel1 and FfLae1, components of a velvet-like complex in Fusarium fujikuroi, affect differentiation, secondary metabolism and virulence. Mol. Microbiol. 2010, 77, 972–994. [Google Scholar] [CrossRef] [PubMed]
- López-Berges, M.S.; Hera, C.; Sulyok, M.; Schäfer, K.; Capilla, J.; Guarro, J.; Di Pietro, A. The velvet complex governs mycotoxin production and virulence of Fusarium oxysporum on plant and mammalian hosts. Mol. Microbiol. 2013, 87, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.; Keller, N.P.; Yu, J.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.S.; Bayram, Ö.; Valerius, O.; Park, H.S.; Irniger, S.; Gerke, J.; Ni, M.; Han, K.H.; Yu, J.H.; Braus, G.H. LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genet. 2010, 6, e1001226. [Google Scholar]
- Amaike, S.; Keller, N.P. Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Keller, N.P. LaeA, a regulator of morphogenetic fungal virulence factors. Eukaryot. Cell 2005, 4, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.Y.; Hou, Y.; Chen, Y.; Oh, J.H.; Lee, I.; Bugni, T.S.; Keller, N.P. Genome-based cluster deletion reveals an endocrocin biosynthetic pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 2012, 78, 4117–4125. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Ji, Y.; Pan, J.; Yu, Y.; Chen, H.; Zhu, X. A new taxol-producing fungus (Pestalotiopsis malicola) and evidence for taxol as a transient product in the culture. Afr. J. Biotechnol. 2011, 10, 6647–6654. [Google Scholar]
- Niu, X.; Hao, X.; Hong, Z.; Chen, L.; Yu, X.; Zhu, X. A putative histone deacetylase modulates the biosynthesis of pestalotiollide B and conidiation in Pestalotiopsis microspora. J. Microbiol. Biotechnol. 2014, 25, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, D.; Zhang, Q.; Yu, X.; Wang, Y.; Zhu, X. Orotidine 5′-phosphate decarboxylase-based reusable in situ genetic editing system: Development and application in taxol-producing Pestalotiopsis microspora. Eng. Life Sci. 2015, 15, 542–549. [Google Scholar] [CrossRef]
- Käfer, E. Meiotic and mitotic recombination in Aspergillus and its chromosomal aberrations. Adv. Genet. 1977, 19, 33–131. [Google Scholar] [PubMed]
- Paz, Z.; García-Pedrajas, M.D.; Andrews, D.L.; Klosterman, S.J.; Baeza-Montañez, L.; Gold, S.E. One step construction of Agrobacterium recombination-ready-plasmids (OSCAR), an efficient and robust tool for ATMT based gene deletion construction in fungi. Fungal Genet. Biol. 2011, 48, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, Y.; Pan, J.; Wei, D.; Zhu, X. High frequency of homologous gene disruption by single-stranded DNA in the taxol-producing fungus Pestalotiopsis microspora. Ann. Microbiol. 2015, 65, 2151–2160. [Google Scholar] [CrossRef]
- Hao, X.; Ji, Y.; Liu, S.; Bi, J.; Bi, Q.; Pan, J.; Zhu, X. Optimized integration of T-DNA in the taxol-producing fungus Pestalotiopsis malicola. Afr. J. Biotechnol. 2012, 11, 771–776. [Google Scholar]
- Zhang, Q.; Chen, L.; Yu, X.; Liu, H.; Akhberdi, O.; Pan, J.; Zhu, X. A B-type histone acetyltransferase Hat1 regulates secondary metabolism, conidiation, and cell wall integrity in the taxol-producing fungus Pestalotiopsis microspora. J. Basic Microbiol. 2016, 56, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Huo, L.; Liu, H.; Chen, L.; Wang, Y.; Zhu, X. Melanin is required for the formation of the multi-cellular conidia in the endophytic fungus Pestalotiopsis microspora. Microbiol. Res. 2015, 179, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Krappmann, S.; Seiler, S.; Vogt, N.; Braus, G.H. Neurospora crassa ve-1 affects asexual conidiation. Fungal Genet. Biol. 2008, 45, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Palonen, E.K.; Raina, S.; Brandt, A.; Meriluoto, J.; Keshavarz, T.; Soini, J.T. Transcriptomic complexity of Aspergillus terreus velvet gene family under the influence of butyrolactone I. Microorganisms 2017, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Estiarte, N.; Lawrence, C.B.; Sanchis, V.; Ramos, A.J.; Crespo-Sempere, A. LaeA and VeA are involved in growth morphology, asexual development, and mycotoxin production in Alternaria alternata. Int. J. Food Microbiol. 2016, 238, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Bouhired, S.; Weber, M.; Kempf-Sontag, A.; Keller, N.P.; Hoffmeister, D. Accurate prediction of the Aspergillus nidulans terrequinone gene cluster boundaries using the transcriptional regulator LaeA. Fungal Genet. Biol. 2007, 44, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, X.; Yin, Y.; Ma, Z. Involvement of a velvet protein FgVeA in the regulation of asexual development, lipid and secondary metabolisms and virulence in Fusarium graminearum. PLoS ONE 2011, 6, e28291. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yun, Y.; Liu, Y.; Ma, Z. FgVELB is associated with vegetative differentiation, secondary metabolism and virulence in Fusarium graminearum. Fungal Genet. Biol. 2012, 49, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Dufour, N.; Rao, R.P. Secondary metabolites and other small molecules as intercellular pathogenic signals. FEMS Microbiol. Lett. 2011, 314, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Rohlfs, M.; Churchill, A.C. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genet. Biol. 2011, 48, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.X.; Yu, C.J.; Wang, M.; Sun, J.N.; Li, Y.Q.; Chen, J. Involvement of a velvet protein ClVelB in the regulation of vegetative differentiation, oxidative stress response, secondary metabolism, and virulence in Curvularia lunata. Sci. Rep. 2017, 7, 46054. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [PubMed]
- Purschwitz, J.; Müller, S.; Kastner, C.; Schöser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue-and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus nidulans VeA subcellular localization is dependent on the importin α carrier and on light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Mooney, J.L.; Yager, L.N. Light is required for conidiation in Aspergillus nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Hoff, B.; Kamerewerd, J.; Sigl, C.; Mitterbauer, R.; Zadra, I.; Kürnsteiner, H.; Kück, U. Two components of a velvet-like complex control hyphal morphogenesis, conidiophore development, and penicillin biosynthesis in Penicillium chrysogenum. Eukaryot. Cell 2010, 9, 1236–1250. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chen, Y.; Ma, Z. Involvement of BcVeA and BcVelB in regulating conidiation, pigmentation and virulence in Botrytis cinerea. Fungal Genet. Biol. 2013, 50, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Myung, K.; Guse, D.; Donkin, B.; Proctor, R.H.; Grayburn, W.S.; Calvo, A.M. FvVE1 regulates filamentous growth, the ratio of microconidia to macroconidia and cell wall formation in Fusarium verticillioides. Mol. Microbiol. 2006, 62, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, J.; Eichhorn, H.; Friedlin, E.; Kürnsteiner, H.; Kück, U. A homologue of the Aspergillus velvet gene regulates both cephalosporin C biosynthesis and hyphal fragmentation in Acremonium chrysogenum. Appl. Environ. Microbiol. 2007, 73, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Oide, S.; Zhang, N.; Choi, M.Y.; Turgeon, B.G. ChLae1 and ChVel1 regulate T-toxin production, virulence, oxidative stress response, and development of the maize pathogen Cochliobolus heterostrophus. PLoS Pathog. 2012, 8, e1002542. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Bok, J.; Brooks, W.; Keller, N.P. veA is required for toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Sempere, A.; Marin, S.; Sanchis, V.; Ramos, A.J. VeA and LaeA transcriptional factors regulate ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2013, 166, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Kopke, K.; Hoff, B.; Bloemendal, S.; Katschorowski, A.; Kamerewerd, J.; Kück, U. Members of the Penicillium chrysogenum velvet complex play functionally opposing roles in the regulation of penicillin biosynthesis and conidiation. Eukaryot. Cell 2013, 12, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Kosalková, K.; García-Estrada, C.; Ullán, R.V.; Godio, R.P.; Feltrer, R.; Teijeira, F.; Martín, J.F. The global regulator LaeA controls penicillin biosynthesis, pigmentation and sporulation, but not roquefortine C synthesis in Penicillium chrysogenum. Biochimie 2009, 91, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.; Zhang, Q.; Wang, D.; Akhberdi, O.; Wei, D.; Zhu, X. Improved pestalotiollide B production by deleting competing polyketide synthase genes in Pestalotiopsis microspora. J. Ind. Microbiol. Biotechnol. 2017, 44, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, D.; Hafner, F.T.; Li, V.; Schmeck, C.; Telser, J.; Vakalopoulos, A.; Wirtz, G. Dibenzodioxocinones—A new class of CETP inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 3611–3614. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG pathway is activated only by two-component signaling pathway in response to osmotic stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, H.; Niu, X.; Akhberdi, O.; Wei, D.; Wang, D.; Zhu, X. The Gα1-cAMP signaling pathway controls conidiation, development and secondary metabolism in the taxol-producing fungus Pestalotiopsis microspora. Microbiol. Res. 2017, 203, 29–39. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence, 5′→3′ |
|---|---|
| VeA-up-s | GGGGACAGCTTTCTTGTACAAAGTGGAAGGGTAGAACGGGACGACT |
| VeA-up-as | GGGGACTGCTTTTTTGTACAAACTTGTGCGGGAAGGGTAAGTGAG |
| VeA-down-s | GGGGACAACTTTGTATAGAAAAGTTGTTCCACCTATTGTTCGCTTGA |
| VeA-down-as | GGGGACAACTTTGTATAATAAAGTTGTGTCCTGGGCATCCTTGTT |
| Ura3(s) | GTCAAGACATCTGTTACCGTGG |
| Ura3(as) | GCAGGCGGGTAGTAGAGT |
| VeA(s) | GGGGACAGCTTTCTTGTACAAAGTGGAAGGGTAGAACGGGACGACT |
| VeA(as) | GGGGACAACTTTGTATAATAAAGTTGTCGACGGAAGCCTGTATCAA |
| Hyg(s) | CCGGTCGGCATCTACTCT |
| Hyg(s) | CGTTGCAAGACCTGCCTGAA |
| ART(F) | CATTCACAAGGCGGGAGA |
| ART(R) | CGAACAATAGGTGGAGGGTC |
| VelB-up-s | GGGGACAGCTTTCTTGTACAAAGTGGAAACGACGGTTGTGGTTCAG |
| VelB-up-as | GGGGACTGCTTTTTTGTACAAACTTGTAGGGCAATCCAGGTAAGC |
| VelB-down-s | GGGGACAACTTTGTATAGAAAAGTTGTTAGGTTATGATACAATCGGGTTA |
| VelB-down-as | GGGGACAACTTTGTATAATAAAGTTGTAGACAAGAGGTGCGGAAA |
| VelB(s) | CGCACAGCCCATTTAGAG |
| VelB(as) | CCAAGCCATCAGATCGTG |
| BRT(F) | GATGGAACCTGGACATACAA |
| BRT(R) | ACGGAGGAGGCGTGATAG |
| LaeA-up-s | GGGGACAGCTTTCTTGTACAAAGTGGAAAACCCTCCACCTCAACAACA |
| LaeA-up-As | GGGGACTGCTTTTTTGTACAAACTTGTTAGCAAGCCAATACACGATG |
| LaeA-down-s | GGGGACAACTTTGTATAGAAAAGTTGTTGGTTGCCCTTGAGCCTTGAA |
| LaeA-down-as | GGGGACAACTTTGTATAATAAAGTTGTCGCCGTATAAATGTACCTTGC |
| LaeA(s) | GGGGACAGCTTTCTTGTACAAAGTGGAATGCCAGGTTGTTCAGTAA |
| LaeA(as) | GGGGACAACTTTGTATAATAAAGTTGTGAGTCGGGCGGGTAGATT |
| LRT(F) | ACCCTCCACCTCAACAAC |
| LRT(F) | CATTAGCAAGCCAATACAC |
| Actin1(s) | GTCGCTGCCCTCGTTATC |
| Actin1(as) | CGAGAATGGAACCACCGAT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhberdi, O.; Zhang, Q.; Wang, D.; Wang, H.; Hao, X.; Liu, Y.; Wei, D.; Zhu, X. Distinct Roles of Velvet Complex in the Development, Stress Tolerance, and Secondary Metabolism in Pestalotiopsis microspora, a Taxol Producer. Genes 2018, 9, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9030164
Akhberdi O, Zhang Q, Wang D, Wang H, Hao X, Liu Y, Wei D, Zhu X. Distinct Roles of Velvet Complex in the Development, Stress Tolerance, and Secondary Metabolism in Pestalotiopsis microspora, a Taxol Producer. Genes. 2018; 9(3):164. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9030164
Chicago/Turabian StyleAkhberdi, Oren, Qian Zhang, Dan Wang, Haichuan Wang, Xiaoran Hao, Yanjie Liu, Dongsheng Wei, and Xudong Zhu. 2018. "Distinct Roles of Velvet Complex in the Development, Stress Tolerance, and Secondary Metabolism in Pestalotiopsis microspora, a Taxol Producer" Genes 9, no. 3: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9030164