
Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation



Abstract
:1. Introduction
2. The Effect of Deficit Irrigation on Morphology, Growth, and Quality of Ornamental Plants

{kind=link}
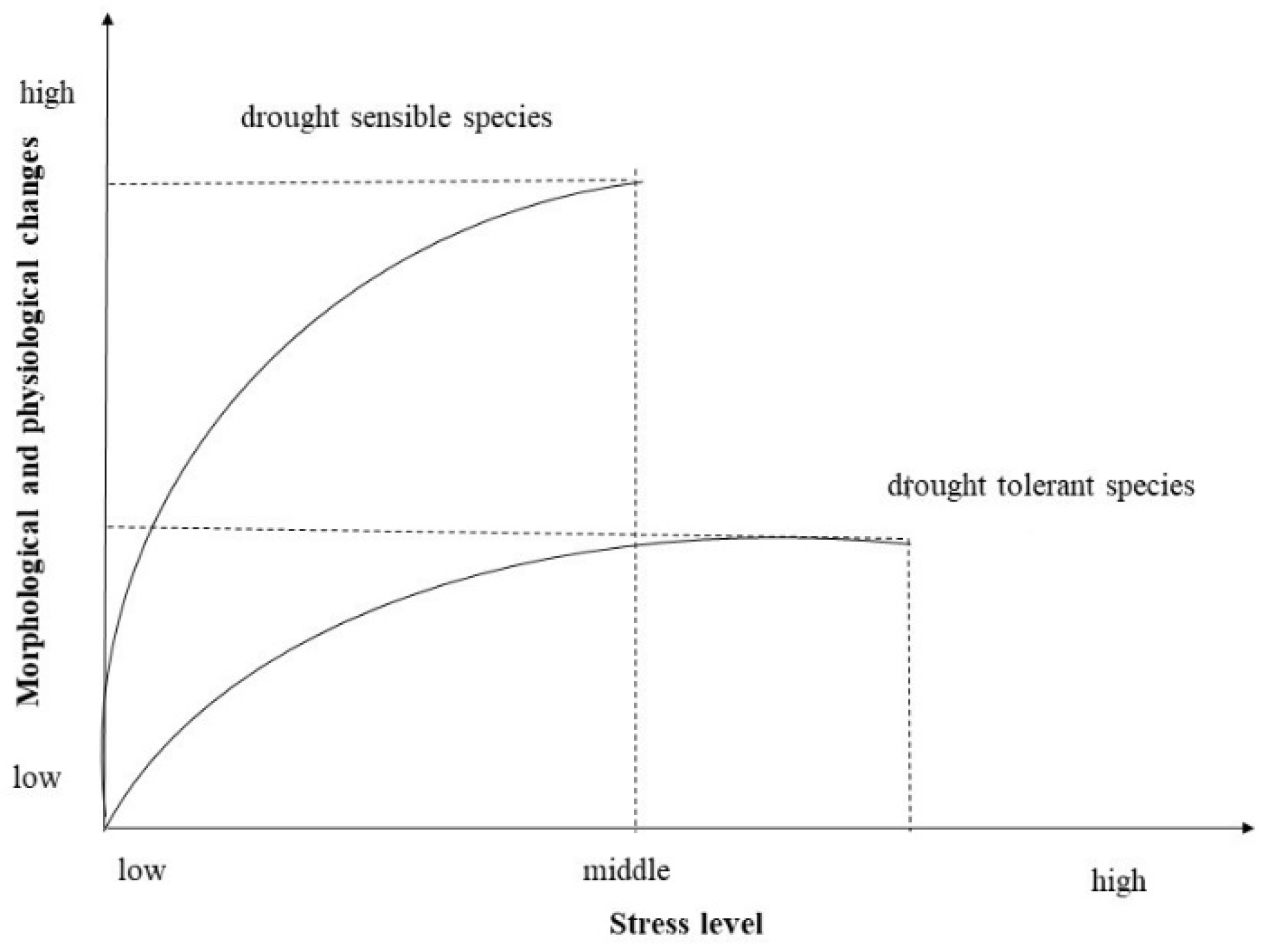{kind=link}
| Species | Plant Habit | Deficit Irrigation Treatment | Plant Growth Stage at the Beginning of Treatment | Modulation of Growth and Morphology by Water Stress | References |
|---|---|---|---|---|---|
| Lantana camara, Ligustrum lucidum | Shrub | C = 100% of water container capacity; Stress: 75%, 50%, and 25% of C | Two month old rooted cuttings | Dry weight (−) Leaf number (−) Leaf area (−) Leaf thickness (−) Thickness of the spongy and palisade tissue (+) | [24] |
| Polygala and Viburnum | 10%, 20%, 30%, 40% of water content of the pot volume | Thickness of the spongy and palisade tissue (+) | [25] | ||
| Malva sylvestris, Althea rosea, Callistephus chinensis and Rudbeckia hirta | Herbaceous plants | C = 100% of ET0 (local reference evapotranspiration) Stress: 25%, 50%, 75% of ET0 | 1-month-old seedlings grown in the field and acclimatized for one month before treatment begun | Root length, root volume, root density: (−) in C. chinensis, and M. sylvestris) Root length: (+) in Althea rosea) Root density: (+) in R. hirta | [26] |
| Bougainvillea glabra var. Sanderiana, Bougainvillea buttiana ‘Rosenka’, Bougainvillea ‘Lindleyana’ (=B. ‘Aurantiaca’) | Rooted cuttings | C = 100% of substrate moisture Stress: 50% and 25% of control | Plants grown in greenhouse into pots filled with 3 L of peat-moss, irrigated with water and nutrient solution | Total dry biomass (−) Leaves number and leaf area (−) Number and flower index (no. dm−2 leaf area) (+) N, P, K (−) | [38] |
| Geranium macrorrhizum L. (Bevan variety from UK, and wild type from Hungary) | Cuttings from rhizome division | Interruption of irrigation for six weeks | Plants grown in greenhouse for 5 months and then a lath house for 7 months, into pots filled with 90% turf, 10% clay, irrigated manually with water | Different leaf area ratio (ratio between the leaf area and total weight of the plant, LAR m2 kg−1). Different leaf mass fraction (LMF, leaf biomass/total biomass; kg kg−1) and root mass fraction (RMF, root biomass/total biomass; kg kg−1) | [41] |
| Nerium oleander L. | Seeds sampled in the wild | Interruption of irrigation for 15 and 30 days | One-year-old seedlings grown in greenhouse, into pots filled with peat-perlite-vermiculite (50%, 25%, 25%), irrigated with nutrient solution for a week before treatment begun | Stem elongation (−), Leaf fresh weight (−), Leaf water content percentage (−) K+/Na+ in roots (−) | [1] |
| Chrysanthemum morifolium Ramat. cv. Hj | Germplasm | 35–40%, 65–70%, 95–100% of soil water holding capacity (WHC), for 62 days | Four-month-old seedlings grown in greenhouse, into plastic pots | Ray florets (−) Disc floret (+) | [28] |
| Viburnum opulus L. and Photinia × fraseri ‘Red robin’ | Shrubs | C = 100% ET (Evapotraspiration) Moderate water deficit = 60% ET Severe water deficit = 30% ET, for 5 months | Plants grown in open air and grenhouse, into pots filled with peat, pumice, and osmocote. | Stem elongation (−) Leaf area (−) Number of leaves (−) Foliar biomass (−) Spongy tissue thickness (+) Shoot/root (+) | [27] |
| Sedum spurium, S. ochroleucum, S. album, and S. sediforme | Herbs, and sub-shrubs | C = irrigation twice a week Stress: interruption of irrigation per 4 weeks | Two-month-old seedlings grown in growth chamber, into pots filled with peat, perlite, and vermiculite, irrigated with nutrient solution | Total stem length (−) Leaves fresh weight (−) | [22] |
| Antirhinum majus cv. butterfly | Seeds | C = 80% of soil water content Stress = 60%, 40%, 20% of soil water content, for 10 weeks | Seedlings grown in greenhouse, into pots filled with sandy loamy soil, irrigated with tap water for three weeks before treatment begun | Shoot height and diameter (−) Number and leaf area (−) Fresh and dry weight of flowers (−) N, P, K, Mg and Ca content (−) | [32] |
| Passiflora spp. (P. edulis, P. gibertii, P. cincinnata, P. alata, P. setacea) | Germplasm | C = 100% of field capacity Stress = interuption of irrigation until apparent wilting (about 96 days) | Seedling grown in greenhouse, into pots. | Plant height (−) Plants dry weight (−) Leaf area (−) Leaves number (−) Different variation of leaf anatomy Stomatal density (+) | [29] |
| Paeonia ostii (Paeonia section Moutan DC) | C = plants watered daily Stress = interruption of irrigation for 4, 8, 12 days | 3-year-old plants grown into pots and watered daily | Change of chloroplasts shape | [30] |
3. Effect of Water Stress on Physiological Parameters, Hormonal Activity, and Biochemical Changes
3.1. Gaseous Exchange
3.2. Chlorophyll Content and Photosynthesis
3.3. Oxidative Stress: ROS Production and Adaptive Responses
3.4. Biochemical Changes
3.5. Hormonal Activity
| Species | Plant Habit | Deficit Irrigation Treatment | Plant Growth Stage at the Beginning of Treatment | Modulation of Physiological Parameters by Water Stress | References |
|---|---|---|---|---|---|
| Sweet briar rose (Rosa rubiginosa L.) | Shrub | Reduced irrigation for 30 days: 11.2 L of water in control plants and 3.6 L in stressed plants (67.9% less). Plants did not receive water in the last three days of experiment | Seedlings, grown in a garden tunnel, into plastic boxes filled with Klasmann-Deilmann TS1 substrate and sand (v/v: 1:2) and irrigated with 11.2 L of water per box | ABA (+3-fold) Gibberellin (+/−). Auxine (+/−). Cytokinin (+/−). | [81] |
| Adonis amurensis and Adonis pseudoamurensis | Middle and lower part of the hillside grassland | C = 32% of soil moisture Stress: interruption of irrigation for 5, 10, 20, and 30 days | Seedlings grown in natural conditions, into polyethylene plastic pots filled with turf and sand, irrigated with water for 4 weeks before experiment begun | RLWC (−); H2O2 (+); MDA (+); Pro (+); Total phenols (+); flavonoids (+); CAT, POD, APX, SOD (+/−); ABA (+); GA (+/−) | [7] |
| Malva sylvestris, Althea rosea, Callistephus chinensis and Rudbeckia hirta | Herbaceous plants | C = 100% of ET0 (local reference evapotranspiration) Stress: 25%, 50%, 75% of ET0 | 1-month-old seedlings grown in the field and acclimated for one month before treatment begun | RLWC (−); Chl a and Chl b (−); Pro (+), Total phenolic compounds (+); EL (+) | [26] |
| Geranium macrorrhizum L. (Bevan variety from UK, and wild type from Hungary) | Cuttings from rhizome division | Interruption of irrigation for six weeks | Plants grown in greenhouse for 5 months and then a lath house for 7 months, into pots filled with 90% turf, 10% clay, irrigated manually with water | Water potential (Ψ) (+) Different amounts of water that the aerial parts (WSL) and roots (WSR) were able to store | [41] |
| Chrysanthemum morifolium Ramat. cv. Hj | Germplasm | 35–40%, 65–70%, 95–100% of soil water holding capacity (WHC), for 62 days | Four-month-old seedlings grown in greenhouse, into plastic pots | Chlorogenic acid; luteoloside, and 3,5-dicaffeoylquinic acid (−) | [28] |
| Viburnum opulus L. and Photinia × fraseri ‘Red robin’ | Shrubs | C = 100% ET Moderate water deficit = 60% ET Severe water deficit = 30% ET, for 5 months | Plants grown in open air and grenhouse, into pots filled with peat, pumice, and osmocote. | Leaf water potential (−) gs (−); ET (−); Pn (−) WUE (+) | [27] |
| Sedum spurium, S. ochroleucum, S. album and S. sediforme | Herbs and sub-shrubs | C = irrigation twice a week Stress: interruption of irrigation per 4 weeks | Two-month-old seedlings grown in growth chamber, into pots filled with peat, perlite, and vermiculite, irrigated with nutrient solution | Chlorophyll a, b and carotenoids (−); MDA (+); Total phenols (+); Total flavonoids (+); Pro (+) | [22] |
| Callistemon citrinus cv Firebrand’ (Crimson bottlebrush) | Rooted cuttings of 2 year-old | C = 100% of container capacity Stress: moderate stress (50% of control) and severe stress (25% of control) | Two-year-old seedlings grown in greenhouse, into pots filled with coconut fiber, peat, and perlite, irrigated with water for three weeks before treatment begun | ET (−); RLWC (−); gs (−); WUE (+) | [14] |
| Rhododendron delavayi | Shrub | C = daily irrigation Stress = interruption of irrigation for 5 and 9 days | Five-year-old plants grown in greenhouse, into pots filled with peat and coconut coir | A (−); ROS (+); Damage to chloroplast ultrastructures | [50] |
| Matthiola incana L. (PanAmerican and Cinderella cultivar) | Seeds | C = 100% of field capacity Stress: 90%, 80%, 70% 60% of field capacity | Seedlings grown in greenhouse, into plastic pots filled with loam, decayed leaves, rotten manure, and river sand (50:25:12.5:12.5), irrigated with tap water, until plants reached the eighth true leaf | Chl (−); CAT (+); anthocyanin content (+); phenolic compounds (+) Pro (+) | [33] |
| Conocarpus lancifolius Engl. (Combretaceae) | Shrub | C = daily irrigation Stress = interruption of irrigation for 12 days | Shoots at the 13-15 leaf growth stage grown in greenhouse, into pots filled with sandy soil and peat, irrigated with distilled water | A (−); Electrons transport (−); Ascorbic acid (−); Flavonoids (+), Phenols (+) | [63] |
| Purple coneflower (Echinacea purpurea L.) | C = 100% of field capacity Stress = 20, 40, 60% of field capacity, until full flowering stage | Seedlings grown in a farm on soil, irrigated until four leaf stage | Chl a and b (−), Carotenoids (+); Pro (+); MDA (−) Enzymes antioxidant activity (+); Phenols (+); Flavonoids (+) | [34] | |
| Paeonia ostii (Paeonia section Moutan DC) | C = plants watered daily Stress = interruption of irrigation for 4, 8, 12 days | 3-year-old plants grown into pots and watered daily | H2O2 (+); O2− (+); RLWC (−); Pro (+); MDA (+); Chl (−); Carotenoids (−); POD, APX activity (+); SOD activity (+/−); Fv/Fm (−) A (−) | [30] |
4. Stress Genes Involved in Plant Tolerance Mechanism against Water Stress
5. Agricultural Practices to Mitigate Water Stress in Ornamental Plants
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, D.; Al Hassan, M.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of salinity and drought on growth, ionic relations, compatible solutes and activation of antioxidant systems in oleander (Nerium oleander L.). PLoS ONE 2017, 12, e0185017. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, J.; Lai, C.; Wang, R.Y.; Chen, X.; Lian, Y. Drying tendency dominating the global grain production area. Glob. Food Secur. 2018, 16, 138–149. [Google Scholar] [CrossRef]
- Marín-de la Rosa, N.; Lin, C.W.; Kang, Y.J.; Dhondt, S.; Gonzalez, N.; Inzé, D.; Falter-Braun, P. Drought resistance is mediated by divergent strategies in closely related Brassicaceae. New Phytol. 2019, 223, 783–797. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Okunlola, G.O.; Olatunji, O.A.; Akinwale, R.O.; Tariq, A.; Adelusi, A.A. Physiological response of the three most cultivated pepper species (Capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Sherwood, S.C.; Alexander, M.J.; Brown, A.R.; McFarlane, N.A.; Gerber, E.P.; Feingold, G.; Scaife, A.A.; Grabowski, W.W. Climate processes: Clouds, aerosols and dynamics. In Climate Science for Serving Society: Research, Modeling and Prediction Priorities; Asrar, G.R., Hurrell, J.W., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 73–103. [Google Scholar] [CrossRef]
- Gao, S.; Wanga, Y.; Yua, S.; Huanga, Y.; Liua, H.; Chena, W.; He, X. Effects of drought stress on growth, physiology and secondary metabolites of two Adonis species in Northeast China. Sci. Hortic. 2020, 259, 108795. [Google Scholar] [CrossRef]
- Hameed, M.; Moradkhani, H.; Ahmadalipour, A.; Moftakhari, H.; Abbaszadeh, P.; Alipour, A. A review of the 21st century challenges in the food-energy-water security in the Middle East. Water 2019, 11, 682. [Google Scholar] [CrossRef] [Green Version]
- Lombardini, L.; Rossi, L. Ecophysiology of plants in dry environments. In Dryland Ecohydrology; Springer: Cham, Switzerland, 2019; pp. 71–100. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamera, K.; Hassana, F.; Alia, E.; Elshazlya, S.; Siddiqued, K.H.M.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Intergovernmental panel on climate change. In Proceeding of the 5th Assessment Report, WGII, Climate Change 2014: Impacts, Adaptation, and Vulnerability; Cambridge University Press: Cambridge, UK, 2014; Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 16 July 2018).
- Zuccarini, P.; Galindo, A.; Torrecillas, A.; Pardossi, A.; Clothier, B. Hydraulic relations and water use of mediterranean ornamental shrubs in container. J. Hortic. Res. 2020, 28, 49–56. [Google Scholar] [CrossRef]
- Gu, Z.; Qi, Z.; Burghate, R.; Yuan, S.; Jiao, X.; Xu, J. Irrigation scheduling approaches and applications: A review. J. Irrig. Drain. Eng. 2020, 146, 04020007. [Google Scholar] [CrossRef]
- Alvarez, S.; Sanchez-Blanco, M.J. Changes in growth rate, root morphology and water use efficiency of potted Callistemon citrinus plants in response to different levels of water deficit. Sci. Hortic. 2013, 156, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Fita, A.; RodrõÂguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flörke, M.; Schneider, C.; McDonald, R.I. Water competition between cities and agriculture driven by climate change and urban growth. Nat. Sustain. 2018, 1, 51–58. [Google Scholar] [CrossRef]
- Nackley, L.L.; de Sousa, E.F.; Pitton, B.J.L.; Sisneroz, J.; Oki, L.R. Developing a water-stress index for potted Poinsettia production. HortScience 2020, 55, 1295–1302. [Google Scholar] [CrossRef]
- Grant, O.M.; Davies, M.J.; Longbottom, H.; Harrison-Murray, R. Evapotranspiration of container ornamental shrubs: Modelling crop-specific factors for a diverse range of crops. Irrig. Sci. 2012, 30, 1–12. [Google Scholar] [CrossRef]
- Ji, K.; Wang, Y.; Sun, W.; Lou, Q.; Mei, H.; Shen, S.; Chen, H. Drought-responsive mechanisms in rice genotypes with contrasting drought tolerance during reproductive stage. J. Plant Physiol. 2012, 169, 336–344. [Google Scholar] [CrossRef]
- Nxele, X.; Klein, A.; Ndimba, B.K. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 2017, 108, 261–266. [Google Scholar] [CrossRef]
- Koźmińskaa, A.; Al Hassana, M.; Wiszniewskab, A.; Hanus-Fajerskab, E.; Boscaiuc, M.; Vicentea, O. Responses of succulents to drought: Comparative analysis of four Sedum (Crassulaceae) species. Sci. Hortic. 2019, 243, 235–242. [Google Scholar] [CrossRef]
- Toscano, S.; Scuderi, D.; Giuffrida, F.; Romano, D. Responses of Mediterranean ornamental shrubs to drought stress and recovery. Sci. Hortic. 2014, 178, 145–153. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Tribulato, A.; Romano, D. Leaf physiological and anatomical responses of Lantana and Ligustrum species under different water availability. Plant Physiol. Biochem. 2018, 127, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Toscano, S.; Ferrante, A.; Tribulato, A.; Romano, D. Response of Mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Rafi, Z.N.; Kazemi, F.; Tehranifar, A. Morpho-physiological and biochemical responses of four ornamental herbaceous species to water stress. Acta Physiol. Plant. 2019, 41, 7. [Google Scholar] [CrossRef]
- Ugolini, F.; Bussotti, F.; Raschi, A.; Tognetti, R.; Roland Enno, A. Physiological performance and biomass production of two ornamental shrub species under deficit irrigation. Trees 2015, 29, 407–422. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, T.; Guo, Q.; Zou, Q.; Yang, F.; Lu, D.; Liu, J. Effect of soil moisture regimes in the early flowering stage on inflorescence morphology and medicinal ingredients of Chrysanthemum morifolium Ramat. Cv. ‘Hangju’. Sci. Hortic. 2020, 260, 108849. [Google Scholar] [CrossRef]
- Souza, P.U.; Kenneddy, L.; Lima, S.; Soares, T.L.; de Jesus, O.N.; Filho, M.A.C.; Girardi, E.A. Biometric, physiological and anatomical responses of Passiflora spp. to controlled water deficit. Sci. Hortic. 2018, 229, 77–90. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Fang, Z.; Wu, Y.; Tao, J. Physiological and transcriptomic analysis of tree Peony (Paeonia section Moutan DC.) in response to drought stress. Forests 2019, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Asrar, A.A.; Abdel-Fattah, G.M.; Elhindi, K.M. Improving growth, flower yield, and water relations of snapdragon (Antirhinum majus L.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi. Photosynthetica 2012, 50, 305–316. [Google Scholar] [CrossRef]
- Jafari, S.; Garmdare, S.E.H.; Azadegan, B. Effects of drought stress on morphological, physiological, and biochemical characteristics of stock plant (Matthiola incana L.). Sci. Hortic. 2019, 253, 128–133. [Google Scholar] [CrossRef]
- Darvizheh, H.; Zahedi, M.; Abbaszadeh, B.; Razmjoo, J. Changes in some antioxidant enzymes and physiological indices of purple coneflower (Echinacea purpurea L.) in response to water deficit and foliar application of salicylic acid and spermine under field condition. Sci. Hortic. 2019, 247, 390–399. [Google Scholar] [CrossRef]
- Mejri, M.; Siddique, K.H.; Saif, T.; Abdelly, C.; Hessini, K. Comparative effect of drought duration on growth, photosynthesis, water relations, and solute accumulation in wild and cultivated barley species. J. Plant Nutr. Soil Sci. 2016, 179, 327–335. [Google Scholar] [CrossRef]
- Farhat, N.; Belghith, I.; Senkler, J.; Hichri, S.; Abdelly, C.; Braun, H.P.; Debez, A. Recovery aptitude of the halophyte Cakile maritima upon water deficit stress release is sustained by extensive modulation of the leaf proteome. Ecotoxicol. Environ. Saf. 2019, 179, 198–211. [Google Scholar] [CrossRef]
- Bañón, S.; Ochoa, J.; Franco, J.; Alarcón, J.; Sánchez-Blanco, M.J. Hardening of oleander seedlings by deficit irrigation and low air humidity. Environ. Exp. Bot. 2006, 56, 36–43. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Caputo, R.; Raimondi, G.; De Pascale, S. The influence of deficit irrigation on growth, ornamental quality, and water use efficiency of rhree potted Bougainvillea genotypes grown in two shapes. HortScience 2014, 49, 1284–1291. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar]
- Garg, B.K. Nutrient uptake and management under drought: Nutrient-moisture interaction. Curr. Agric. 2003, 27, 1–8. [Google Scholar] [CrossRef]
- Navarro Rocha, J.; Burillo-Alquézar, J.; Aibar-Lete, J.; González-Coloma, A. Adaptability of two accessions of Geranium macrorrhizum L. to drought stress conditions. J. Appl. Res. Med. Aromat. Plants 2017, 7, 149–152. [Google Scholar] [CrossRef]
- Forner, A.; Valladares, F.; Bonal, D.; Granier, A.; Grossiord, C.; Aranda, I. Extreme droughts affecting Mediterranean tree species’ growth and water-use efficiency: The importance of timing. Tree Physiol. 2018, 38, 1127–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, T.; Li, Z.; Song, C.; Song, S.; Jiao, J.; Liu, Y.; Dong, Z.; Zheng, X. Contrasting drought tolerance in two apple cultivars associated with difference in leaf morphology and anatomy. Am. J. Plant Sci. 2019, 10, 709–722. [Google Scholar] [CrossRef] [Green Version]
- Jin, N.; Ren, W.; Tao, B.; He, L.; Ren, Q.; Li, S.; Yu, Q. Effects of water stress on water use efficiency of irrigated and rainfed wheat in the Loess Plateau, China. Sci. Total Environ. 2018, 642, 1–11. [Google Scholar] [CrossRef]
- Campitelli, B.E.; Des Marais, D.L.; Juenger, T.E. Ecological interactions and the fitness effect of water-use efficiency: Competition and drought alter the impact of natural MPK12 alleles in Arabidopsis. Ecol. Lett. 2016, 19, 424–434. [Google Scholar] [CrossRef]
- Cameron, R.W.F.; Harrison-Murray, R.S.; Atkinson, C.J.; Judd, H.L. Regulated deficit irrigation: A means to control growth in woody ornamentals. J. Hortic. Sci. Biotechnol. 2006, 81, 435–443. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chiatanya, K.V.; Vivekanandan, M. Drought induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Álvarez, S.; Navarro, A.; Nicolás, E.; Sánchez-Blanco, M.J. Transpiration, photosynthetic responses, tissue water relations and dry mass partitioning in Callistemon plants during drought conditions. Sci. Hortic. 2011, 129, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Medrano, H. Energy dissipation in C3 plants under drought. Funct. Plant Boil. 2002, 29, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.-F.; Wang, J.H.; Zhang, L.; Song, J.; Peng, L.C.; Zhang, S.B. Physiological and transcriptomic analysis highlight key metabolic pathways in relation to drought tolerance in Rhododendron delavayi. Physiol. Mol. Biol. Plants 2019, 25, 991–1008. [Google Scholar] [CrossRef]
- Impa, S.M.; Nadaradjan, S.; Jagadish, S.V.K. Drought stress induced reactive oxygen species and anti-oxidants in plants. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 131–147. [Google Scholar] [CrossRef]
- Anjum, S.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Wang, L. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 8, 1056–1071. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Catala, M.; Silva, J.M.D.; Branquinho, C.; Barreno, E. The impact of dehydration rate on the production and cellular location of reactive oxygen species in an aquatic moss. Ann. Bot. 2012, 110, 1007–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, V.; Ranjan, S.; Deeba, F.; Pandey, A.K.; Singh, R.; Shirke, P.A.; Pathre, U.V. Desiccation-induced physiological and biochemical changes in resurrection plant, Selaginella bryopteris. J. Plant Physiol. 2010, 167, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Ines, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and non-enzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Moreno, D.A.; Ferreres, F.; del Mar Rubio-Wilhelmi, M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Bautista, I.; Boscaiu, M.; Lidon, A.; Llinares, J.V.; Lull, C.; Donat, M.P.; Mayoral, O.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 9. [Google Scholar] [CrossRef]
- Redha, A.; Al-Mansor, N.; Suleman, P.; Al-Hasan, R.; Afzal, M. Modulation of antioxidant defenses in Conocarpus lancifolius under variable abiotic stress. Biochem. Syst. Ecol. 2012, 43, 80–86. [Google Scholar] [CrossRef]
- Azzedine, F.; Gherroucha, H.; Baka, M. Improvement of salt tolerance in Durum wheat by ascorbic acid application. J. Stress Physiol. Biochem. 2011, 7, 27–37. [Google Scholar]
- Hessini, K.; Martinez, J.P.; Gandour, M.; Albouchi, A.; Soltani, A.; Abdelly, C. Effect of water stress on growth, osmotic adjustment, cell wall elasticity and water use efficiency in Spartina alterniflora. Environ. Exp. Bot. 2009, 67, 312–319. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Hessini, K.; Kronzucker, H.J.; Abdelly, C.; Cruz, C. Drought stress obliterates the preference for ammonium as an N source in the C4 plant Spartina alterniflora. J. Plant Physiol. 2017, 213, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.P.; Lutts, S.; Schanck, A.; Bajji, M.; Kinet, J.M. Is osmotic adjustment required for water stress resistance in the Mediterranean shrub (Atriplex halimus L.)? J. Plant Physiol. 2004, 161, 1041–1051. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Cicevan, R.; Al Hassan, M.; Sestras, A.F.; Prohens, J.; Vicente, O.; Sestras, R.E.; Boscaiu, M. Screening for drought tolerance in cultivars of the ornamental genus Tagetes (Asteraceae). PeerJ 2016, 4, e2133. [Google Scholar] [CrossRef] [Green Version]
- Magdy, M.; Mansour, M.; Farouk, E. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef]
- Pourghayoumi, M.; Rahemi, M.; Bakhshi, D.; Aalami, A.; Kamgar-Haghighi, A.A. Responses of pomegranate cultivars to severe water stress and recovery: Changes on antioxidant enzyme activities, gene expression patterns and water stress responsive metabolites. Physiol. Mol. Biol. Plants 2017, 23, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.S.; Elnemr, K.F. Plant response to drought stress simulated by ABA application: Changes in chemical composition of cuticular waxes. Environ. Exp. Bot. 2013, 86, 70–75. [Google Scholar] [CrossRef]
- Ban, S.G.; Selak, G.V.; Leskovar, D.I. Short- and long-term responses of pepper seedlings to ABA exposure. Sci. Hortic. 2017, 225, 243–251. [Google Scholar] [CrossRef]
- Kowitcharoen, L.; Wongs-Aree, C.; Setha, S.; Komkhuntod, R.; Srilaong, V.; Kondo, S. Changes in abscisic acid and antioxidant activity in sugar apples under drought conditions. Sci. Hortic. 2015, 193, 1–6. [Google Scholar] [CrossRef]
- Zhang, S.H.; Xu, X.F.; Sun, Y.M.; Zhang, J.L.; Li, C.Z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.H.; Han, H.Y.; Zhang, Y.L.; Zhang, W.F. Effects of drought and re-watering on endogenous hormone contents of cotton roots and leaves under drip irrigation with mulch. J. Appl. Ecol. 2013, 24, 1009–1016. [Google Scholar]
- Gadzinowska, J.; Dziurka, M.; Ostrowskaa, A.; Hura, K.; Hura, T. Phytohormone synthesis pathways in sweet briar rose (Rosa rubiginosa L.) seedlings with high adaptation potential to soil drought. Plant Physiol. Biochem. 2020, 154, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, J.J.; Zhang, W.J.; Yan, S.N.; Wang, R.; Zhao, J.F.; Li, Y.J.; Qi, Z.G.; Sun, Z.X.; Zhu, Z.G. The putative auxin efflux carrier OsPIN3t is involved in the drought stress response and drought tolerance. Plant J. 2012, 72, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Tommasini, L.; Svensson, J.T.; Rodriguez, E.M.; Wahid, A.; Malatrasi, M.; Kato, K.; Wanamaker, S.; Resnik, J.; Close, T.J. Dehydrin gene expression provides an indicator of low temperature and drought stress: Transcriptome-based analysis of barley (Hordeum vulgare L.). Funct. Integr. Genom. 2008, 8, 387–405. [Google Scholar] [CrossRef] [PubMed]
- Klay, I.; Gouia, S.; Liu, M.; Mila, I.; Khoudi, H.; Bernadac, A.; Bouzayen, M.; Pirrello, J. Ethylene Response Factors (ERF) are differentially regulated by different abiotic stress types in tomato plants. Plant Sci. 2018, 274, 137–145. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundertmark, M.; Hincha, D.K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Wang, H.; Tong, Y.; Wang, Y. Insights into the Superoxide Dismutase Gene Family and Its Roles in Dendrobium catenatum under Abiotic Stresses. Plants 2020, 9, 1452. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xia, M.; Chen, J.; Yuan, R.; Deng, F.; Shen, F. Gene expression characteristics and regulation mechanisms of superoxide dismutase and its physiological roles in plants under stress. Biochemistry 2016, 81, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Massa, D.; Lenzi, A.; Montoneri, E.; Ginepro, M.; Prisa, D.; Burchi, G. Plant response to biowaste soluble hydrolysates in hibiscus grown under limiting nutrient availability. J. Plant Nutr. 2018, 41, 396–409. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J.; Harrison-Murray, R.; Atkinson, C.J.; Grant, O.M. Application of deficit irrigation to container-grown hardy ornamental nursery stock via overhead irrigation, compared to drip irrigation. Agric. Water Manag. 2016, 163, 244–254. [Google Scholar] [CrossRef] [Green Version]


| Gene | Species | Cellular or Subcellular Localisation | Activity during Water Stress | References |
|---|---|---|---|---|
| DcaCSD1-2-3-4 | Dendrobium catenatum | Chloroplast (DcaCSD1), citoplasm | Cu/ZnSOD synthesis | [88] |
| DcaFSD1-2-3 | Dendrobium catenatum | Chloroplast | Fe/SOD synthesis | [88] |
| DcaMSD1 | Dendrobium catenatum | Mitochondrion | [88] | |
| HjCYC2c | Chrysanthemum morifolium | Young inflorescence | Adjusting of shape flowers of Chrysanthemum morifolium | [28] |
| FLS | Chrysanthemum morifolium | Young inflorescence | Adjusting of pathways of flavonoids during water stress | [28] |
| Lhca, Lhcb (18 genes), Psa (11 genes), Psb (15 genes) (all involved in photosynthetic apparatus synthesis), F3H, DFR, ANS (flavonoids biosynthesis) PP2C (abscisic acid synthesis), BAK1 and BRI1 (brassinosteroids synthesis) | Rhododendron delavayi | Leaves | Response to stimulus; Biosynthesis of secondary metabolites (flavonoids and brassinosteroids); Synthesis of photosystem I and II proteins, and electron transport chain proteins; Synthesis of ATP synthase | [50] |
| F3H, CCOAOMT, CYP98A CAD, GLU, ZEP, NCED, CCD, TKL, RPI, FBP, KCS, ECH, PPT, LOX, CYP, ORP | Paeonia ostii | Leaves | Increase of proline, flavonoids, stilbenoid, diarylheptanoid, and gingerol. Reduction of chlorophylls, carotenoids, phenylpropane and fatty acids. | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, M.; Petropoulos, S.A.; Cirillo, C.; Rouphael, Y. Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation. Horticulturae 2021, 7, 107. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050107
Giordano M, Petropoulos SA, Cirillo C, Rouphael Y. Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation. Horticulturae. 2021; 7(5):107. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050107
Chicago/Turabian StyleGiordano, Maria, Spyridon A. Petropoulos, Chiara Cirillo, and Youssef Rouphael. 2021. "Biochemical, Physiological, and Molecular Aspects of Ornamental Plants Adaptation to Deficit Irrigation" Horticulturae 7, no. 5: 107. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050107