
Plant Production and Leaf Anatomy of Mertensia maritima (L.) Gray: Comparison of In Vitro Culture Methods to Improve Acclimatization

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and In Vitro Establishment
2.2. Multiple Shoot Induction
2.3. Comparison between Solid and TIS Culture Systems for Shoot Micropropagation
2.4. Comparison between Solid and TIS Culture Systems for Rooting Induction
2.5. Acclimatization Tests
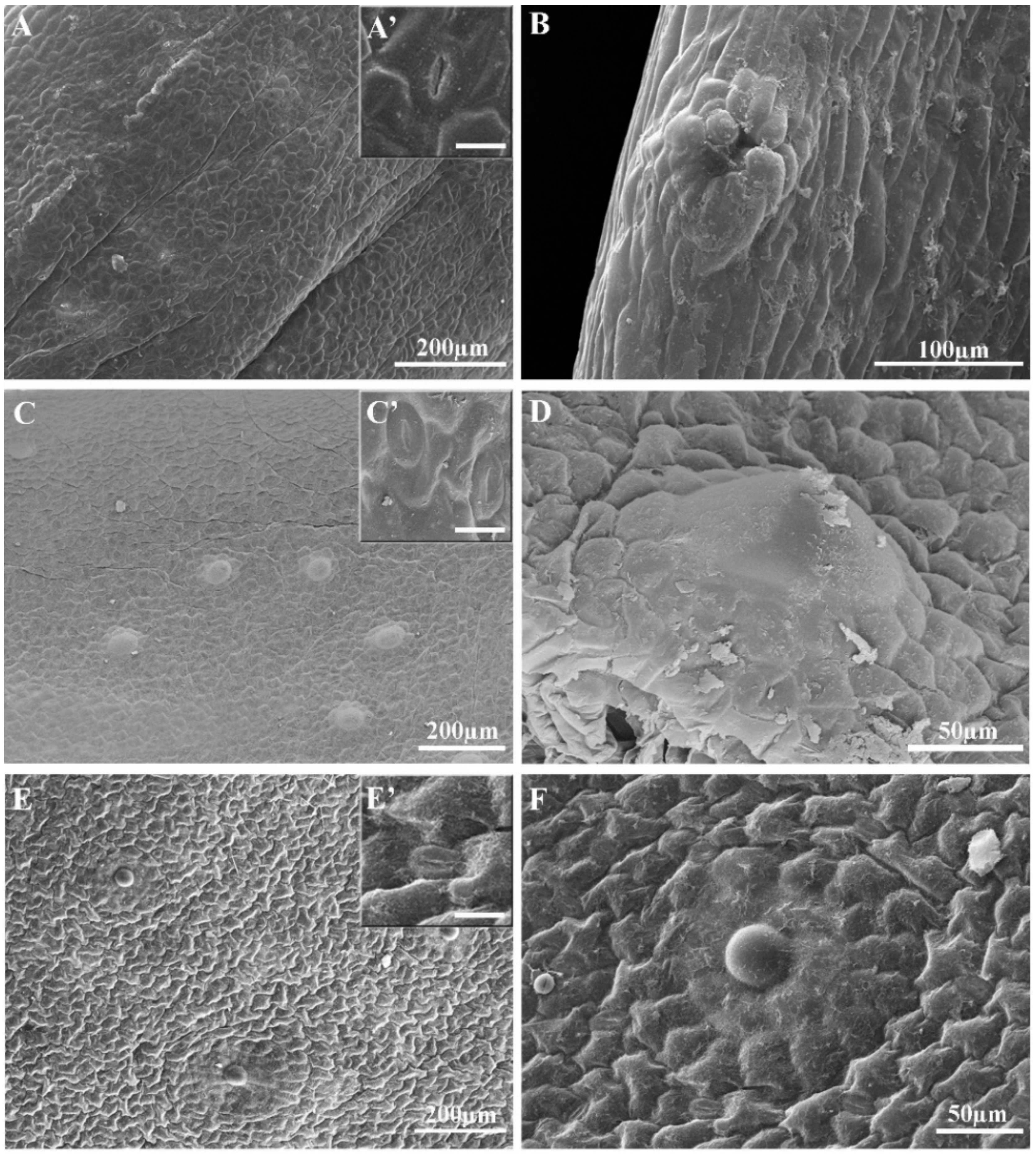
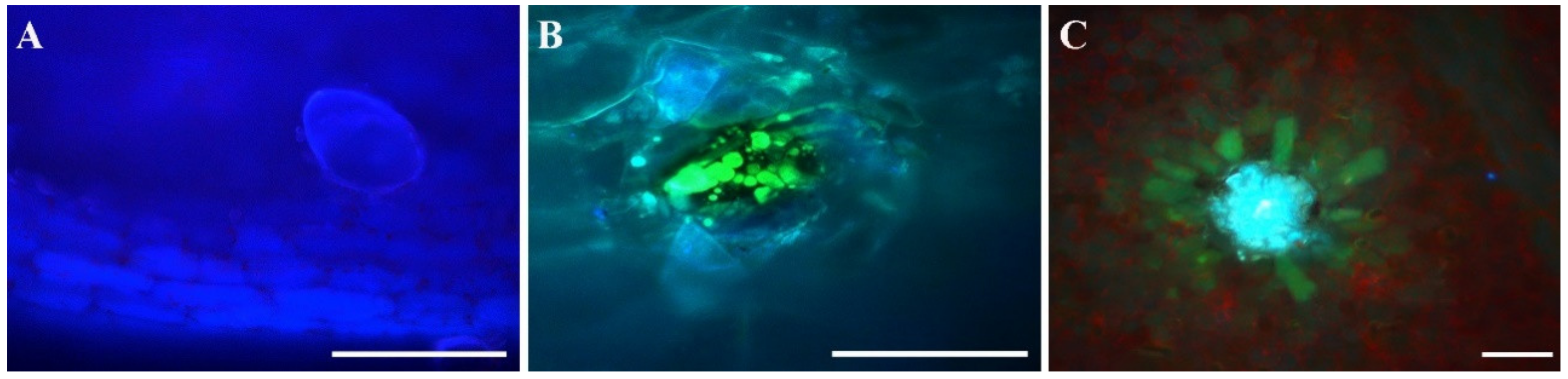
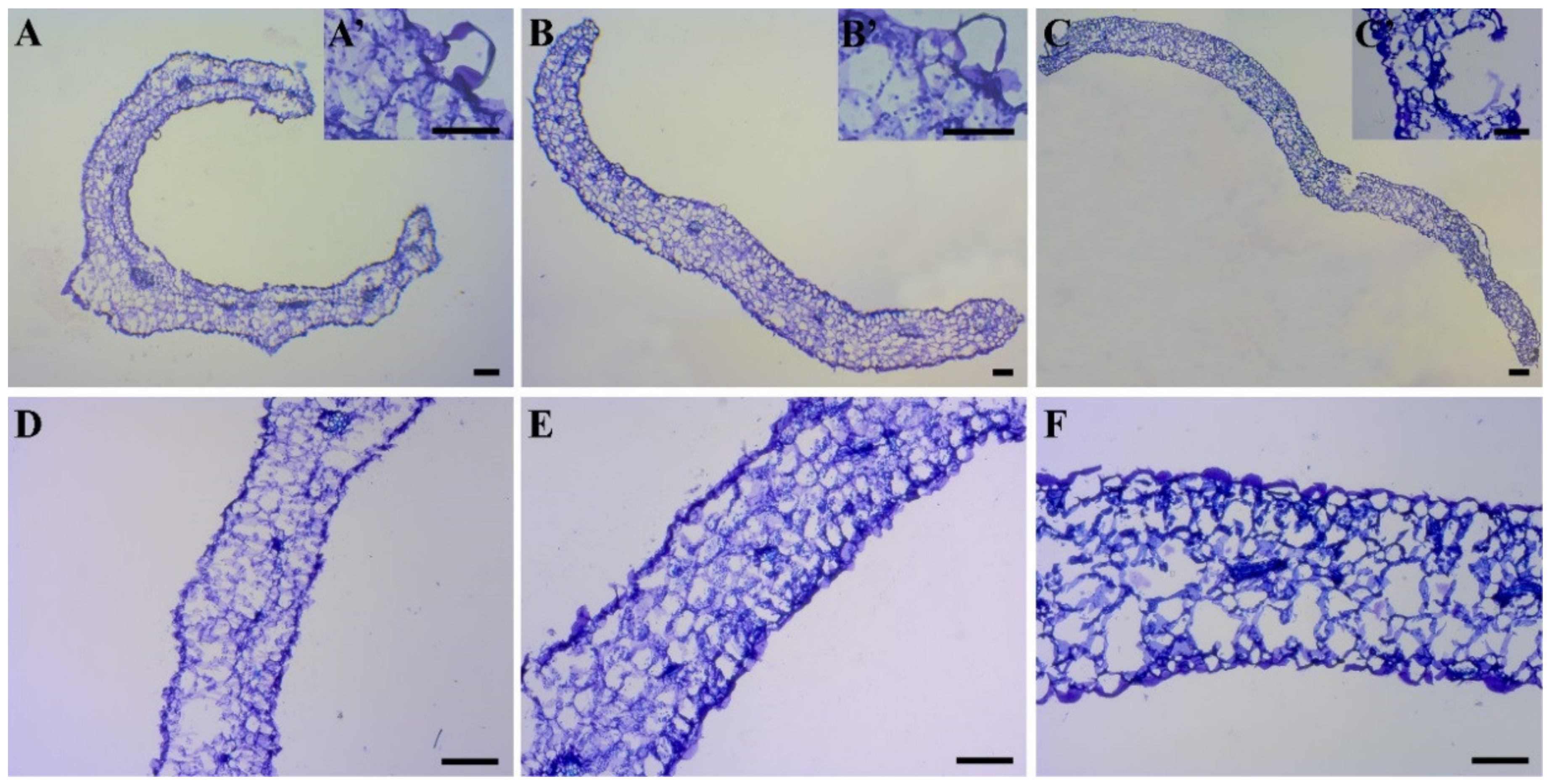
2.6. Leaf Morphological and Anatomical Examination
2.7. Statistical Analysis
3. Results
3.1. Shoot Micropropagation
3.2. Rooting Induction
3.3. Plant Acclimatization
3.4. Leaf Anatomical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alton, S.; FitzGerald, R. Mertensia maritima Boraginaceae. Curtis’s Bot. Mag. 2009, 26, 96–110. [Google Scholar] [CrossRef]
- Delort, E.; Jaquier, A.; Chapuis, C.; Rubin, M.; Starkenmann, C.J. Volatile composition of oyster leaf (Mertensia maritima (L.) Gray). J. Agric. Food Chem. 2012, 60, 11681–11690. [Google Scholar] [CrossRef]
- Burkovskaya, E.V.; Khrolenko, Y.A. Ecological features of the photosynthetic complex structure of Mertensia maritima (Boraginaceae) from different geographic locations. Bull. KrasGAU. Biol. Sci. 2015, 8, 53–58. (In Russian) [Google Scholar]
- Skarpaas, O.; Stabbetorp, E. Diaspore ecology of Mertensia maritima: Effects of physical treatments and their relative timing on dispersal and germination. Oikos 2003, 95, 374–382. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, D.H.; Saini, R.K.; Gopal, J.; Keum, Y.S.; Sivanesan, I. Micropropagation and quantification od bioactive compounds in Mertensia maritima (L.) Gray. Int. J. Mol. Sci. 2019, 20, 2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.; Sivanesan, I.; Ak, G.; Zengin, G.; Cziàky, Z.; Jeko, J.; Rengasamy, K.R.R.; Lee, O.N.; Kim, D.H. Screening of biological activities of calli, shoots, and seedlings of Mertensia maritima (L.) Gray. Plants 2020, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Ruffoni, B.; Savona, M. The temporary immersion system (T.I.S.) for the improvement of micropropagation of ornamental plants. Acta Hortic. 2003, 683, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Barberini, S.; Savona, M.; Ruffoni, B. Temporary immersion culture of Lilium bulbiferum. Acta Hortic. 2010, 900, 377–383. [Google Scholar] [CrossRef]
- Savona, M.; Sacco, E.; Ruffoni, B. Improving micropropagation performances in Hydrangea spp.: Temporary immersion shoot culture and induction of morphogenetic events. Acta Hortic. 2011, 961, 457–464. [Google Scholar] [CrossRef]
- Ruta, C.; De Mastro, G.; Ancona, S.; Tagarelli, A.; De Cillis, F.; Benelli, C.; Lambardi, M. Large-scale plant production of Licium barbarum L. by liquid culture in temporary immersion system and possible application to the synthesis of bioactive substance. Plants 2020, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell Tissue Organ Cult. 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Koroch, A.R.; Villani, T.S.; Pyne, R.M.; Simon, J.E. Rapid staining method to detect and identify downy mildew (Peronospora belbahrii) in basil. Appl. Plant Sci. 2013, 1, 1300032. [Google Scholar] [CrossRef] [PubMed]
- Chieco, C.; Rotondi, A.; Morrone, L.; Rapparini, F.; Baraldi, R. An ethanol-based fixation method for anatomical and micro-morphological characterization of leaves of various tree species. Biotech. Histochem. 2013, 88, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan Red/B or Fluoral Yellow 088 in polyethylene glycol-glicerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Tateo, F.; Bononi, M.; Mariotti, M.G.; Cornara, L.; Serrato-Valenti, G. Trichomes on vegetative and reproductive organs of Stevia rebaudiana (Asteraceae). Structure and secretory products. Plant Biosyst. 2001, 135, 25–37. [Google Scholar] [CrossRef]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999; ISBN 0-19-508956-1. [Google Scholar]
- Bassolino, L.; Giacomelli, E.; Giovanelli, S.; Pistelli, L.; Cassetti, A.; Damonte, G.; Bisio, A.; Ruffoni, B. Tissue culture and aromatic profile in Salvia dolomitica Codd. Plant Cell Tiss. Organ. Cult. 2015, 121, 83–95. [Google Scholar] [CrossRef]
- Malik, S.; Sharma, M.; Ahuja, P.S. An efficient and economic method for in vitro propagation of Arnebia euchroma using liquid culture system. Am. J. Biotech. Med. Res. 2016, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Welander, M.; Persson, J.; Asp, H.; Zhu, L.H. Evaluation of a new vessel system based on temporary immersion system for micropropagation. Sci. Hortic. 2014, 179, 227–232. [Google Scholar] [CrossRef]
- Ahmadian, M.; Babaei, A.; Shokri, S.; Hessami, S. Micropropagation of carnation (Dianthus caryophyllus L.) in liquid medium by temporary immersion bioreactor in comparison with solid culture. J. Genet. Eng. Biotechnol. 2017, 15, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Gianguzzi, V.; Inglese, P.; Barone, E.; Sottile, F. In vitro regeneration of Capparis spinosa L. by using a temporary immersion system. Plants 2019, 8, 177. [Google Scholar] [CrossRef] [Green Version]
- Mc Alister, B.; Finnie, J.; Watt, M.P.; Blakeway, F. Use of the temporary immersion bioreactor system (RITA®) for production of commercial Eucalyptus clonesin mondi Forest (SA). Plant Cell Tiss. Organ. Cult. 2005, 81, 347–358. [Google Scholar] [CrossRef]
- Yan, H.; Liang, C.; Li, Y. Improved growth and quality of Siraitia grosvenorii plantlets using a temporary immersion system. Plant Cell Tiss. Organ. Cult. 2010, 103, 131–135. [Google Scholar] [CrossRef]
- Akdemir, H.; Suzerer, V.; Onay, A.; Tilkat, E.; Ersali, Y.; Ciftci, Y.O. Micropropagation of the pistachio and its rootstocks by temporary immersion system. Plant Cell Tiss. Organ. Cult. 2014, 117, 65–76. [Google Scholar] [CrossRef]
- Kumar, K.; Rao, I.U. Morphophysiological problems in acclimatization of micropropagated plants in ex vitro conditions—A reviews. J. Ornam. Hortic. Plants 2012, 2, 271–283. [Google Scholar]
- Martinez-Estrada, E.; Islas-Luna, B.; Pérez-Sato, A.; Bello-Bello, J.J. Temporary immersion improves in vitro multiplication and acclimatization of Anthurium andreanum Lind. Sci. Hortic. 2019, 249, 185–191. [Google Scholar] [CrossRef]
- Belaeva, T.N.; Butenkova, A.N. Leaf blade anatomy of the rare Siberian flora species Mertensia sibirica (L.) G. Don fil. (Boraginaceae). Ukr. J. Ecol. 2020, 10, 186–191. [Google Scholar] [CrossRef]
- Selvi, F.; Bigazzi, M. Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomy. Flora 2001, 196, 269–285. [Google Scholar] [CrossRef]
- Barykina, R.P.; Alyonkin, V. Pubescence of vegetative organs and trichome micromorphology in some Boraginaceae at different ontogenetic stages. Wulfenia 2016, 23, 1–29. [Google Scholar]
- Fedoreyev, S.A.; Inyushkina, Y.V.; Bulgakov, V.P.; Veselova, M.V.; Tchernoded, G.K.; Gerasimenko, A.V.; Zhuravlev, Y.N. Production of allantoin, rabdosiin and rosmarinic acid in callus cultures of the seacoastal plant Mertensia maritima (Boraginaceae). Plant Cell Tiss. Organ Cult. 2012, 110, 183–188. [Google Scholar] [CrossRef]
- Ventrella, M.C.; Marinho, C. Morphology and histochemistry of glandular trichomes of Cordia verbenacea DC. (Boraginaceae) leaves. Braz. J. Bot. 2008, 31, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Dassanayake, M.; Larkin, J.C. Making plants break a sweat: The structure, function, and evolution of plant salt glands. Front. Plant Sci. 2017, 8, 406. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.G. Diversity and classification of flowering plants: Eudicots. In Plant Systematics, 2nd ed.; Academic Press: Cambridge, MA, USA, 2010; pp. 275–448. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Jar | TIS |
|---|---|---|
| Shoot cluster fresh weight (g) | 0.909 ± 0.121 a | 1.040 ± 0.193 a |
| Shoot cluster dry weight (g) | 0.033 ± 0.003 a | 0.036 ± 0.013 a |
| Shoot number per cluster | 9 ± 1 a | 6 ± 1 b |
| Leaf number | 25 ± 2 a | 15 ± 2 a |
| Leaf length (mm) | 13 ± 1 b | 42 ± 2 a |
| Length of leaf blade (mm) | 5 ± 1 b | 15 ± 1 a |
| Width of leaf blade (mm) | 2 ± 1 b | 7 ± 1 a |
| Parameters | Jar | TIS |
|---|---|---|
| Shoot fresh weight (g) | 0.392 ± 0.033 a | 0.925 ± 0.102 b |
| Root number | 4 ± 1 a | 14 ± 1 b |
| Root length (mm) | 2 ± 1 a | 9 ± 1 b |
| Leaf number | 11 ± 1 a | 12 ± 1 a |
| Leaf length (mm) | 20 ± 1 a | 37 ± 1 b |
| Length of leaf blade (mm) | 6 ± 1 a | 12 ± 1 b |
| Width of leaf blade (mm) | 3 ± 1 a | 5 ± 1 b |
| Treatments 1 | Survival Rate (%) |
|---|---|
| JG | 3.3 ± 3.3 a |
| PG | 5.0 ± 2.9 a |
| JC | 11.7 ± 1.7 b |
| PC | 56.7 ± 4.4 c 2 |
| Treatments | Test | Target Compounds | Response |
|---|---|---|---|
| Jar | Toluidine Blue O | Polyphenols and tannins | - |
| Fluorol yellow | Terpenoids and lipids | - | |
| Ruthenium Red | Non cellulosic polysaccharides | - | |
| Sudan III | Total lipids | - | |
| Autofluorescence | Various | + | |
| Plantform | Toluidine Blue O | Polyphenols and tannins | +/− |
| Fluorol Yellow | Terpenoids and lipids | +/− | |
| Ruthenium Red | Non cellulosic polysaccharides | - | |
| Sudan III | Total lipids | - | |
| Autofluorescence | Various | + | |
| In vivo plant | Toluidine Blue O | Polyphenols and tannins | + |
| Fluorol Yellow | Terpenoids and lipids | + | |
| Ruthenium Red | Non cellulosic polysaccharides | + | |
| Sudan III | Total lipids | + | |
| Autofluorescence | Various | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copetta, A.; Bazzicalupo, M.; Cassetti, A.; Marchioni, I.; Mascarello, C.; Cornara, L.; Pistelli, L.; Ruffoni, B. Plant Production and Leaf Anatomy of Mertensia maritima (L.) Gray: Comparison of In Vitro Culture Methods to Improve Acclimatization. Horticulturae 2021, 7, 111. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050111
Copetta A, Bazzicalupo M, Cassetti A, Marchioni I, Mascarello C, Cornara L, Pistelli L, Ruffoni B. Plant Production and Leaf Anatomy of Mertensia maritima (L.) Gray: Comparison of In Vitro Culture Methods to Improve Acclimatization. Horticulturae. 2021; 7(5):111. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050111
Chicago/Turabian StyleCopetta, Andrea, Miriam Bazzicalupo, Arianna Cassetti, Ilaria Marchioni, Carlo Mascarello, Laura Cornara, Laura Pistelli, and Barbara Ruffoni. 2021. "Plant Production and Leaf Anatomy of Mertensia maritima (L.) Gray: Comparison of In Vitro Culture Methods to Improve Acclimatization" Horticulturae 7, no. 5: 111. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050111