3.2. Canopy Growth, Light Microclimate and Disease Development
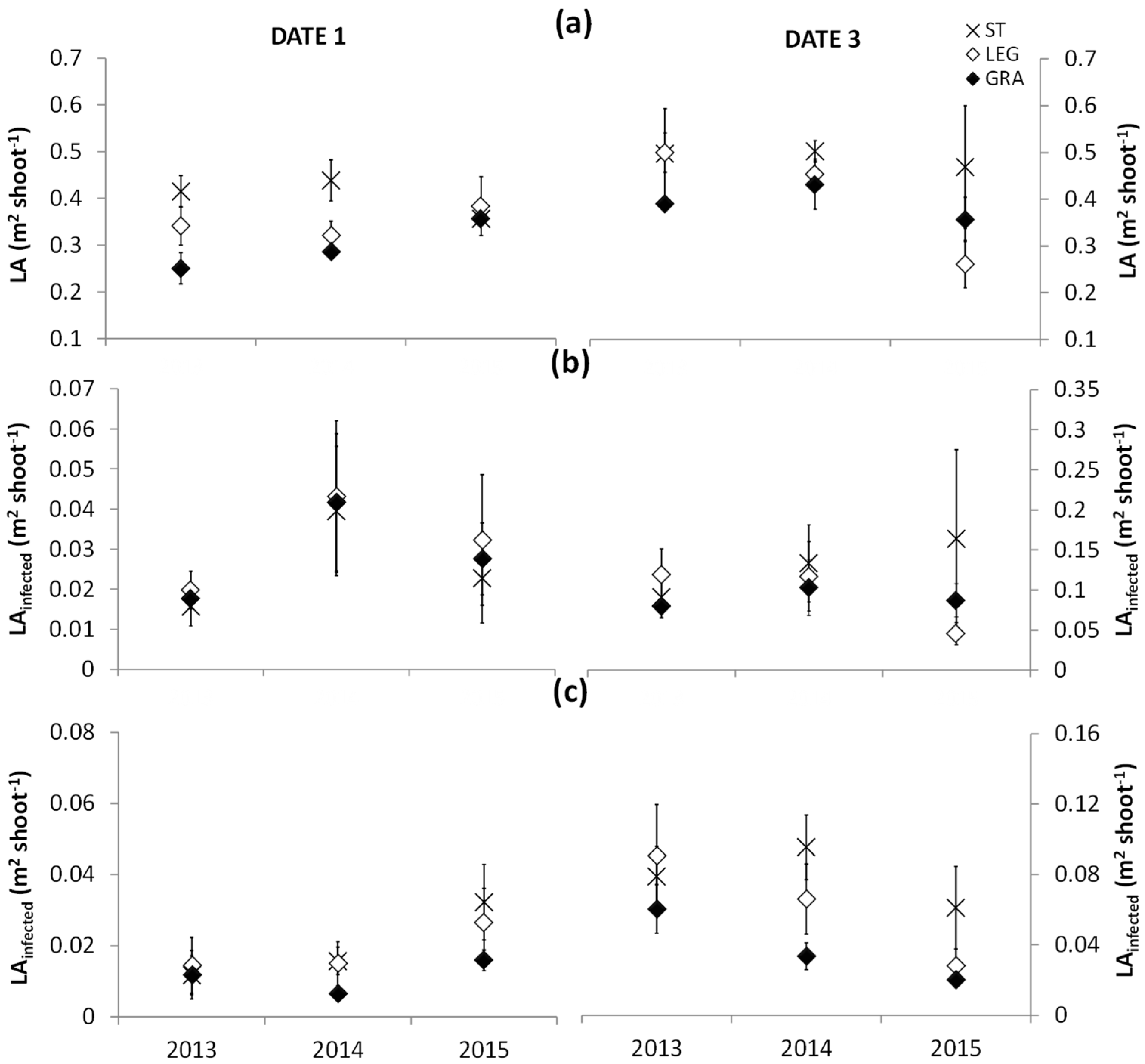
The estimation of LA growth in dates 1 and 3 showed statistically significant differences between the soil tillage and cover-crop treatments in the three study seasons (
Figure 4a). During the first period of disease monitoring, which also corresponded to intense LA development, the grapevines grown in tilled soil presented higher values of total LA compared to those managed with cover crops, mostly due to a higher leaf size both for main and lateral leaves, since the number of leaves per shoot was not statistically different among treatments (
Appendix A Table A1). The LA composed of lateral shoots, in 2013–2014–2015, represented, respectively, 40–40–30% of the total LA, while the LEG and GRA plants presented, respectively, 30–40–30% and 20–20–40% of the LA composed of lateral shoots. When the last disease development observations were performed (date 3), the differences in total LA among treatments were reduced and became statistically insignificant. However, an increase in the development of lateral shoots in the cover-crop plots, particularly with legumes, promoted canopy growth. In the cover-crop treatments, total LA reached similar values than those observed under soil tillage and the LA composed by lateral shoots in 2013–2014–2015 was, respectively, 50–40–50% in ST, 40–30–20% in LEG, and 30–40–40% in GRA. No significant differences in the main and lateral LA infected by downy mildew were detected among treatments (
Figure 4b). Yet, it is important to highlight that the differences in LA per shoot between ST, LEG, and GRA resulted in significant differences and increases in the powdery mildew-infected LA when maximum canopy development was reached (
Figure 4c). In fact, the ST canopies presented significantly higher average values of LA infected by powdery mildew than GRA in date 3 (
Table 2). Among the two cover crops, higher values were recorded in LEG (
Figure 4c), though not statistically significant. These results confirmed those obtained by Valdés-Gómez et al. [
40], who observed a major explanatory role of the number of leaves per shoot, and thus of canopy vigor on powdery mildew development in early season.
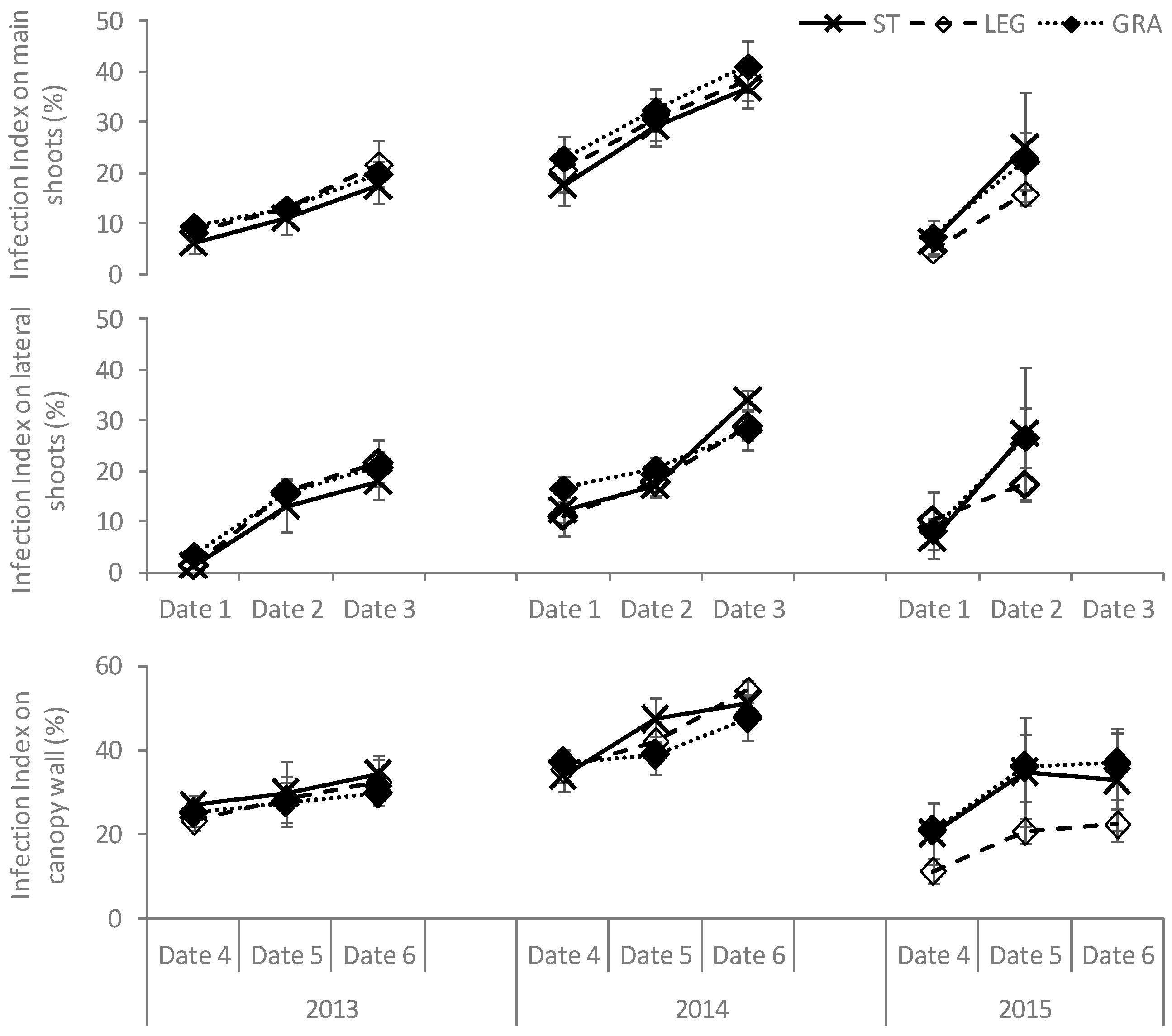
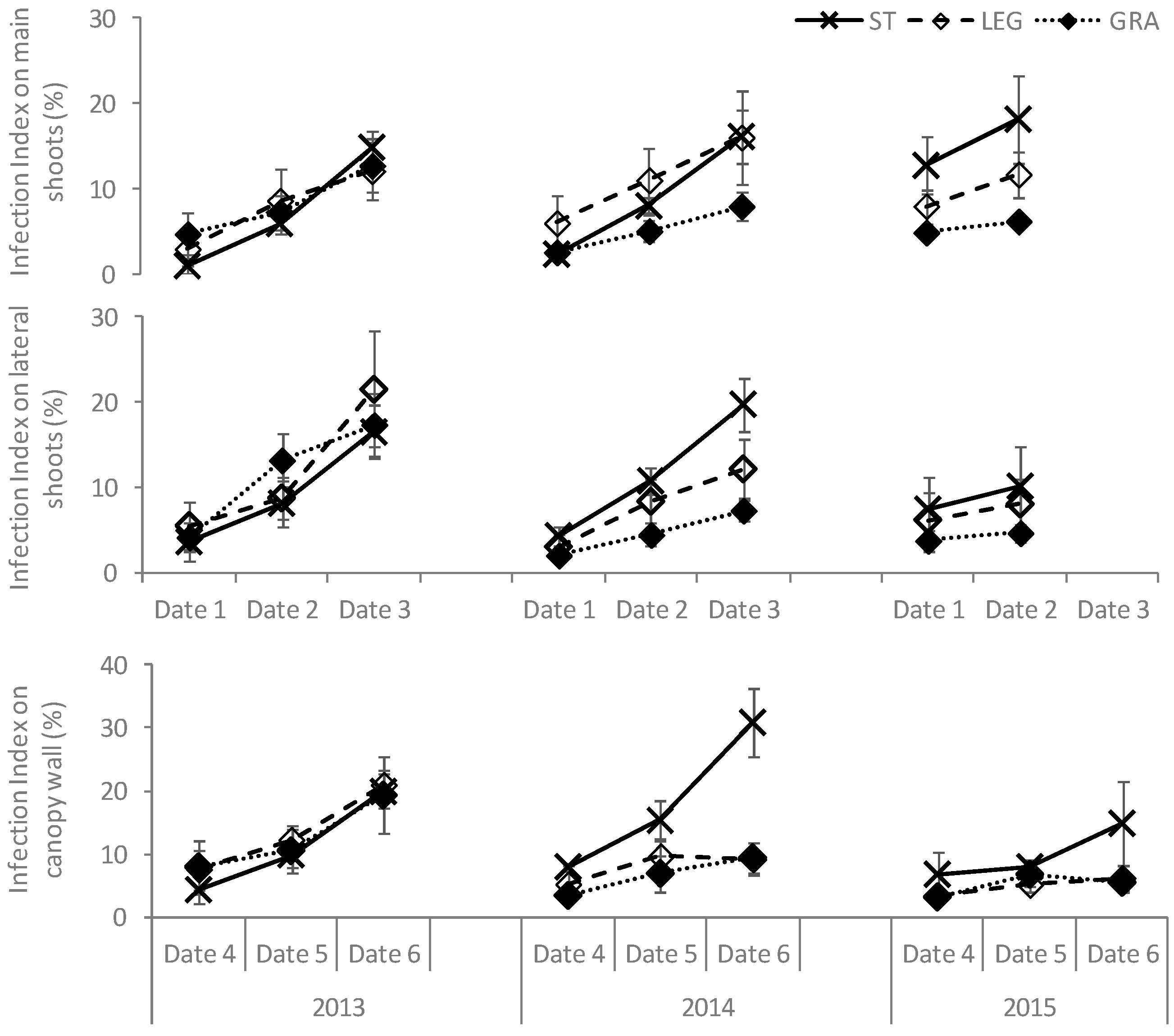
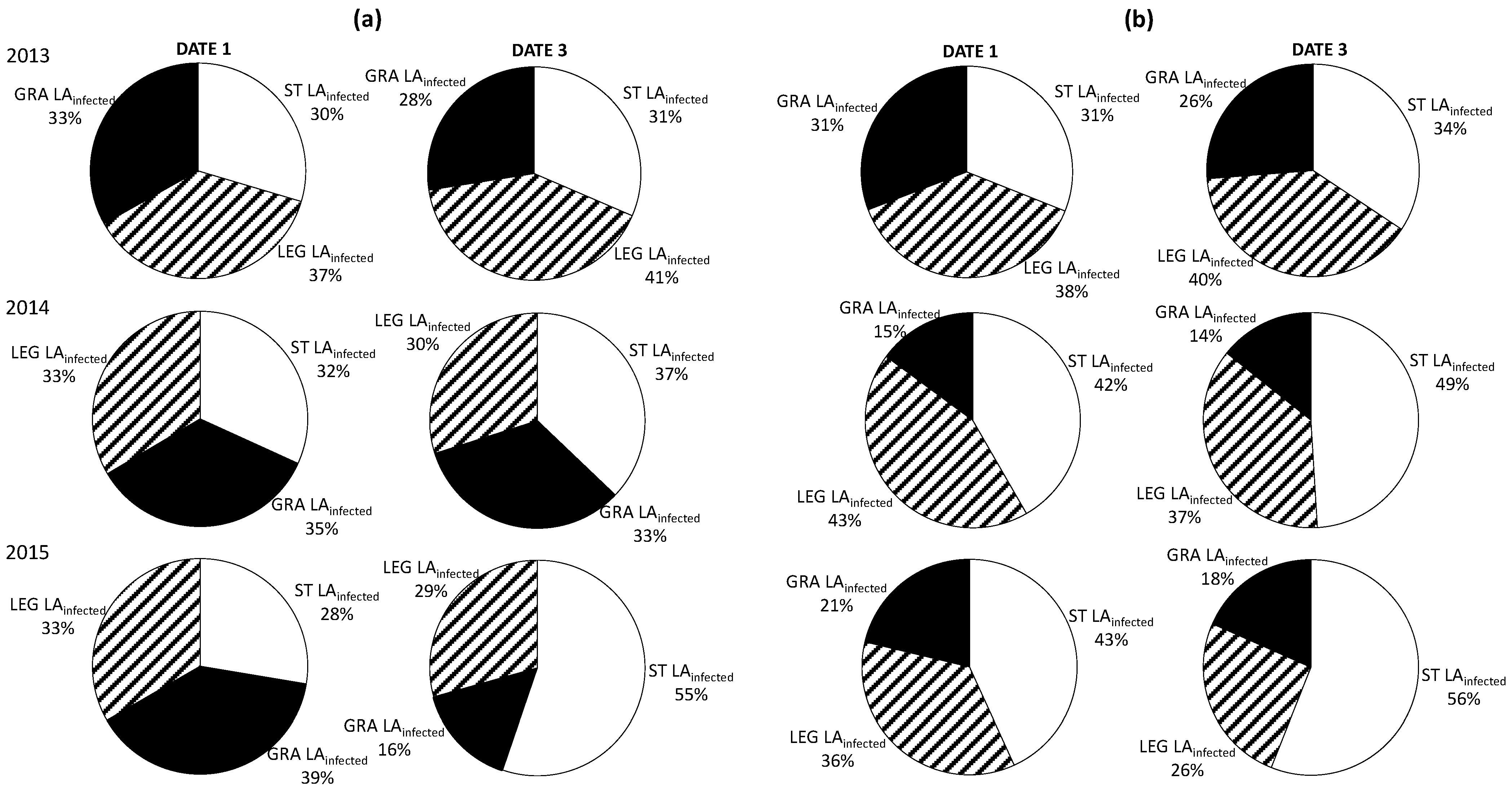
However, soil management treatments influenced the development of both diseases in the long term. In 2013, the proportion of infected LA was similar in the three treatments for both diseases (
Figure 5) while it increased significantly in ST as compared to the cover-cropped plots in the subsequent years. During the first season, higher proportions of both downy and powdery mildew-infected LA were detected in LEG grapevines. In 2014, the proportion of LA infected by
E. necator was significantly lower in GRA-treated vines, right from the first dates of disease observation, while the differences in total LA affected by
P. viticola were not significant between the cover-crop treatments. Finally, in 2015, the spread of downy mildew in GRA plants was significantly lower in date 3 and the differences between the two cover crops increased, while for powdery mildew the differences between LEG and GRA were flattened.
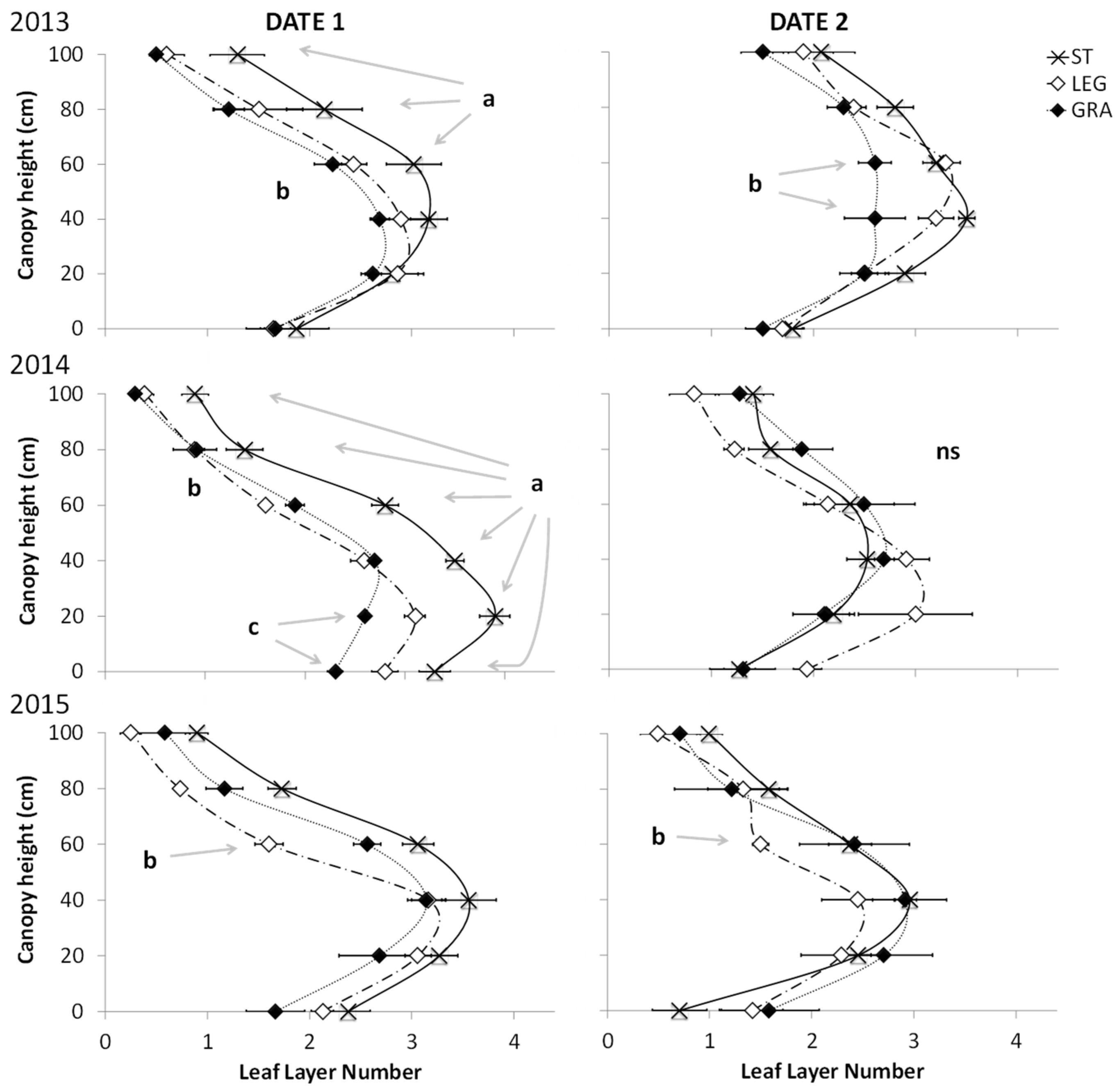
During the period of intense canopy growth and up to fruit set (date 1), soil tillage induced a higher canopy density, both at the cluster level and in the above parts of the canopy (
Figure 6). The higher density was characterized by a higher LLN, lower porosity (particularly in the top 80–100 cm), and a higher PIL (
Table 3,
Figure 6).
When the last disease monitoring observations were performed (date 6), differences among treatments concerning canopy density were only statistically significant at the mid canopy height in 2013, when lower values were recorded in GRA. Furthermore, the two-way ANOVA showed the significant effects of season on the mean canopy density variables in the last date of disease monitoring, and a significant effect of soil management in the first date of disease observation (
Table 3), with a lower porosity, as well as higher LLN and PIL in ST plants compared to those under cover crops. The percentage of exposed clusters was not statistically different between treatments and years.
The measurements of canopy wall size also showed a higher and wider structure and a larger exposed LA in the vines with soil tillage than in the cover-cropped ones. The highest values were measured in date 2, in ST canopies (about 2.8 and 3.3 m2 per vine of exposed LA and 40 and 43 cm of average width, in 2013 and 2014, respectively), and the lowest values were measured in GRA (about 2.9 and 3.1 m2 exposed LA and 39 and 37 cm of average width, in the first two seasons). However, both the exposed LA and leaf wall width measured in LEG were not statistically different from that observed in the GRA or ST plants (respectively, about 2.9 m2 and 42 cm in 2013 and 3.2 m2 and 38 cm in 2014) (data not shown).
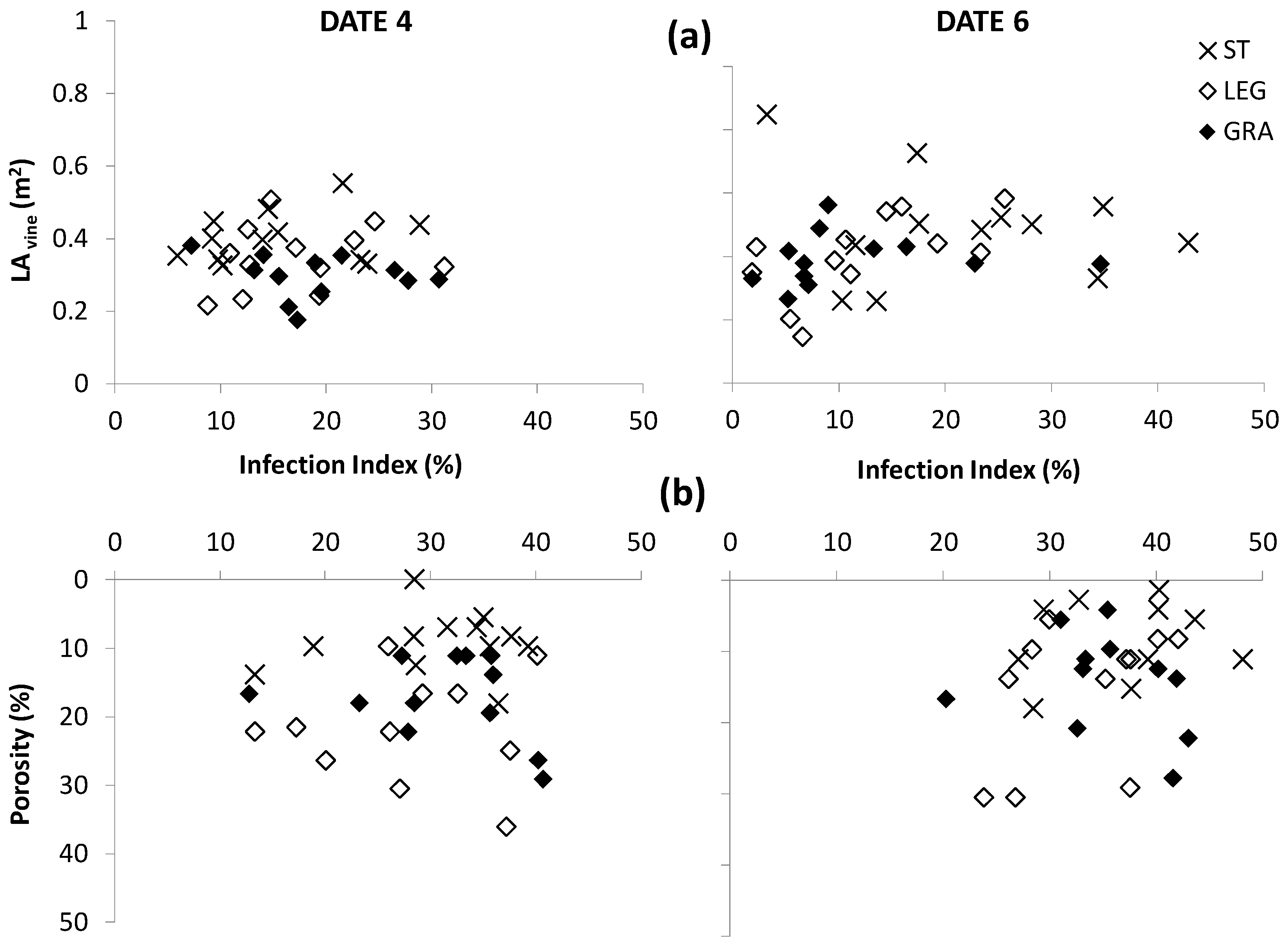
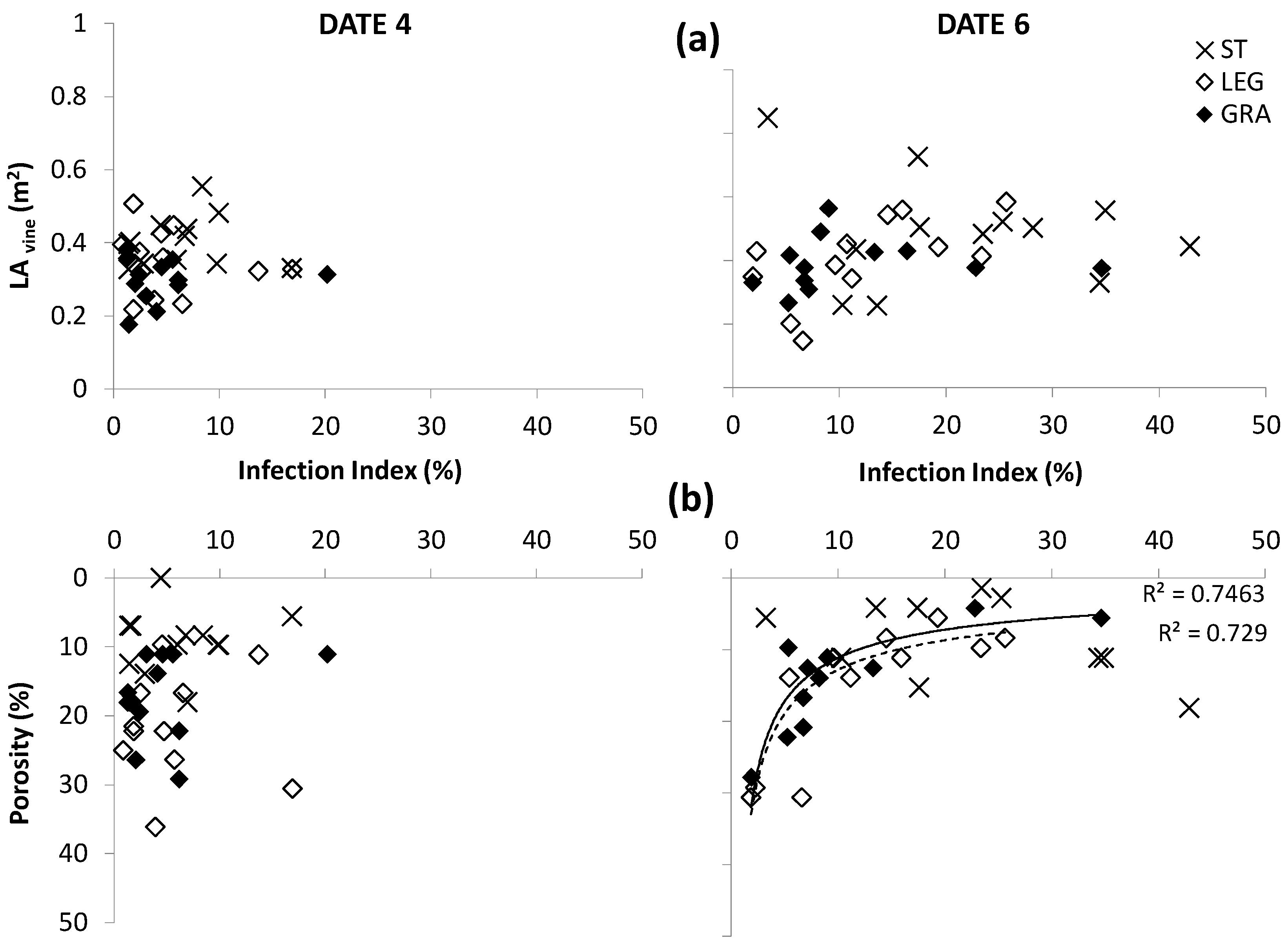
Besides LA growth, also the differences in canopy porosity among treatments had a significant effect on the spread of powdery mildew on the canopy wall after the first observation date but the relation between canopy density and disease spread was not clearly evident for downy mildew (
Figure 7 and
Figure 8). In fact, taking into account the average data of the three-year study period, no simple regression between the total LA and infection index was observed in all treatments (
Figure 7a and
Figure 8a).
However, the cover crops resulted in a lower canopy density and this played an important role in reducing the spread of
E. necator in the canopy wall (
Figure 8b). By observation date 6, for the vines subjected to cover-crop treatments, it was possible to represent the development of powdery mildew infection on the canopies with a similar logistic curve, which is the typical model of micro-organism population growth whenever it is strongly dependent upon the available biomass [
41].
When the biomass was evaluated as a function of leaf N, expressed by the mean SPAD values (
Table A2), significantly higher N were found on the grapevine leaves of ST and LEG compared to GRA, in seasons 2013 and 2015 [
35]. Besides, for the first period of disease development in all three years of the trial, the GRA leaves showed lower values than the other two treatments. Moreover, in 2014, leaf N was significantly different among all treatments in date 1—lower in GRA and higher in ST—as compared to the data collected in date 2. When the last observations of disease symptoms were performed (date 3), the differences in SPAD values between GRA and LEG had already increased, due to both the increased N values in LEG and decreased N in GRA. These differences in leaf N among grapevines under soil tillage and cover crops were probably fostered by the complete soil cover. In most studies conducted in Mediterranean climate conditions, mineral balance differences are evident in soil samples and slight in grapevine leaves and petioles [
42]. Besides, legume cover tends to induce higher N contents than soil tillage under cover-cropped inter-rows [
43,
44,
45].
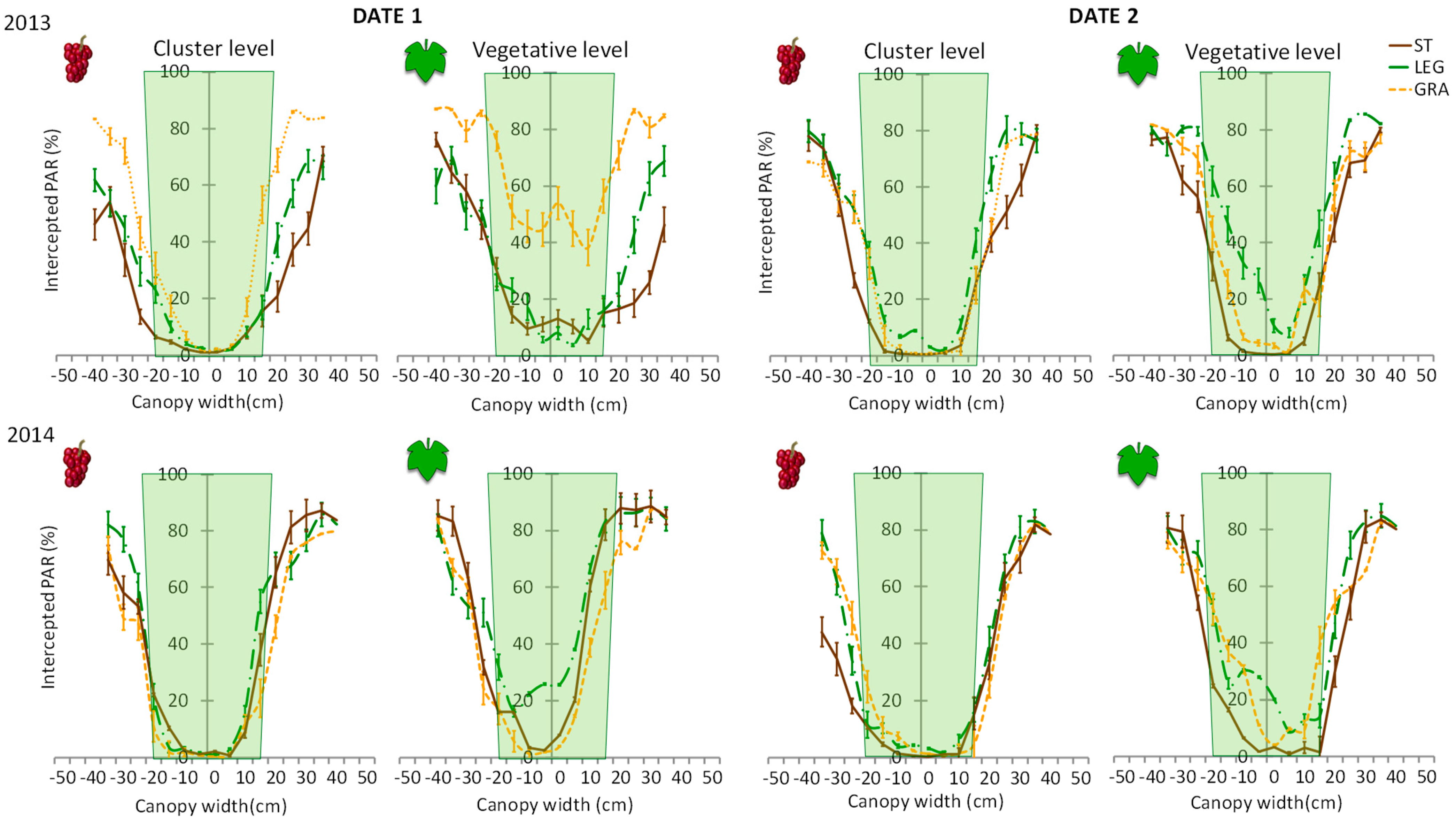
The PAR measurements carried out close to date 1 of the disease observations in shoots showed greater light interception in the inner canopy layers of GRA. Statistically more significant differences were observed at the middle canopy height rather than at the cluster level, and in the tilled plots, significantly less sunlight penetration occurred (
Figure 9). Taking into consideration that the PAR measurements were performed at the solar zenith, the magnitude of the differences among treatments in sunlight penetration into the inner canopy layers was considerably different across the day [
30].
Overall, these results are in accordance with those of Guerra and Steenwerth [
12], who observed a higher canopy efficiency and better growth balance, leading to less favorable conditions for disease development in cover-cropped vineyards. The competition for water and nutrients among grapevine and cover crops, which may lead to yield reductions [
5,
35], also determines a better light microclimate and canopy balance for improving berry composition [
46,
47].
The greater development of powdery mildew on the plots managed with soil tillage is related to the effects of the higher vigor and shade on the canopy light microclimate. In fact, throughout its life cycle,
E. necator develops on the outer surfaces of the leaves and berries and thus can be exposed to the negative effect of direct sunlight incidence on the external canopy layer and/or of a sun-fleck on the inner canopy layers of a sparse canopy. Similarly, Austin et al. [
48] reported the important role played by both canopy density and sunlight levels in the canopy on reducing the development of powdery mildew at the cluster zone.
3.4. Weight and Interaction among the Epidemiology Factors for Disease Development
In the three-year period, cover crops induced a lower canopy development and density than soil tillage. The differences in canopy structure and density induced in the ST grapevines a lower radiation interception at the cluster zone and in the inner canopy during the stages of intense canopy growth. However, the development of lateral shoots after fruit set allowed to reach, in the cover-crop plots, LA values similar to those of the tilled plots, especially in the LEG treatment. Nevertheless, light penetration into the canopy of the GRA plants was greater. For this reason, under high grapevine vigor conditions, the use of grass cover crops may also be an effective means to guarantee good canopy microclimate conditions and therefore reduce the need for leaf or lateral shoot removal operations during berry growth and ripening stages.
In addition, to assess the influence of soil management on plant diseases, other than the infection index, it is important to consider the infected LA and the differences in canopy density among treatments, in order to evaluate the actual spread of diseases in the vineyard. As observed in this study, differences in average leaf size, LA, or leaf age can exert significant effects on both tissue attractiveness and the overall extent of the attack, which may foster the differences in infection among treatments to become significant [
44].
Taking into account the strong inter-annual variability that can be found in the vineyard, due to both the duration of the weather conditions favorable for the development of diseases and the evolution of the cover-crop composition and settlement, it is essential to identify and weight the relationships between the set of explanatory variables that show correlations with disease incidence under the different soil management regimes.
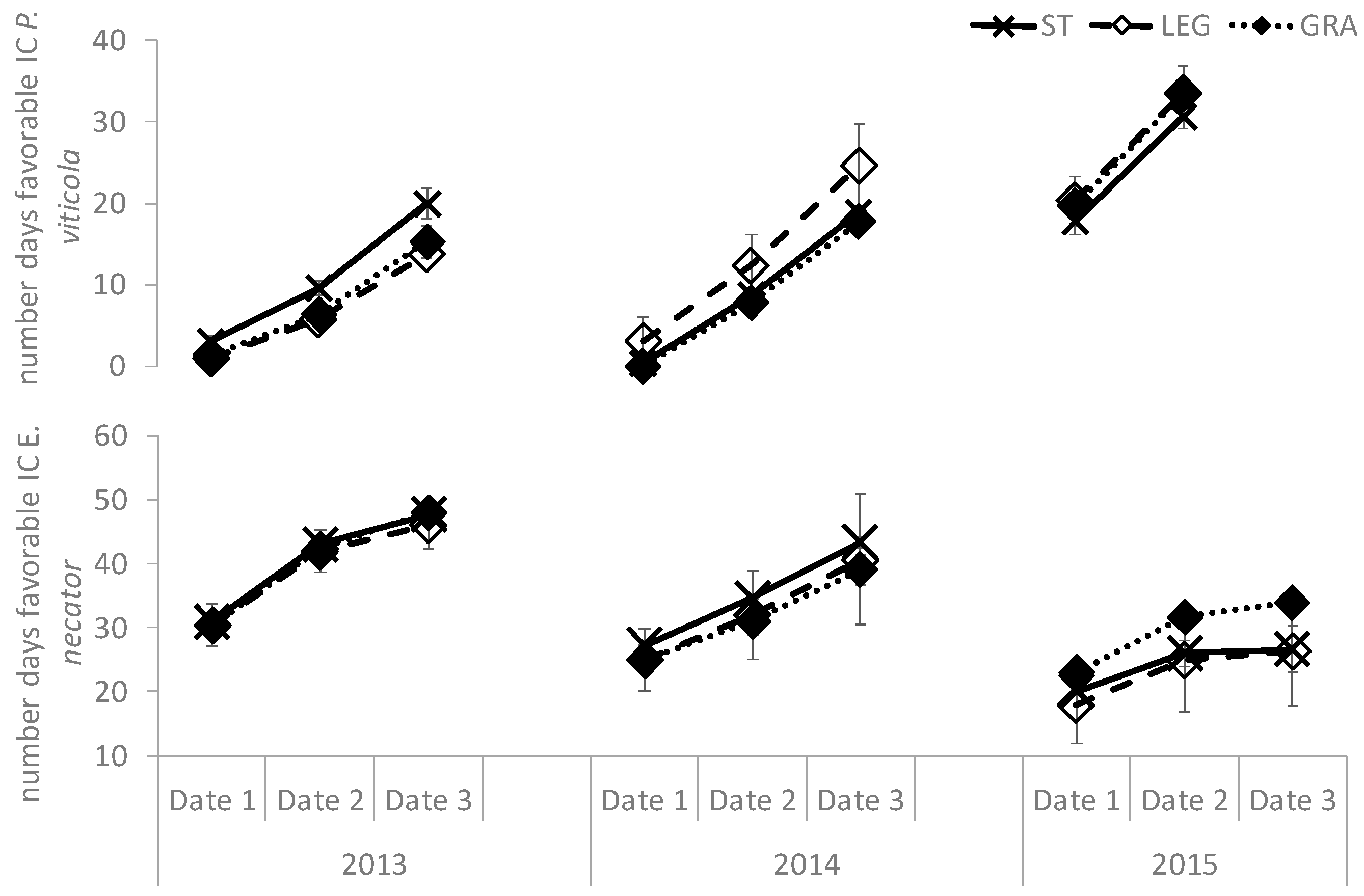
As far as downy mildew is concerned, it is likely that the main effect of the meteorological pattern was the most influential factor regulating the development of
P. viticola, since no significant differences among treatments concerning disease development were observed in the fields during the whole study period. A similar result, as well as the major dependence of disease development upon weather conditions, have been previously reported [
28,
49]. Furthermore, the role played by direct sunlight exposure is probably less influential on the development of this disease, in contrast to
E. necator, as the downy mildew infections take place in the dark and develop inside the leaf and berry tissues without any exposure to direct sunlight, as is the case for powdery mildew. To better elucidate this issue and in order to weight the relative importance of the variables affecting disease development and the relation among them in each soil management treatment, a multifactor analysis was performed for dates 1 and 3 of both diseases development in shoots for the whole study period (
Figure 12 and
Figure 13).
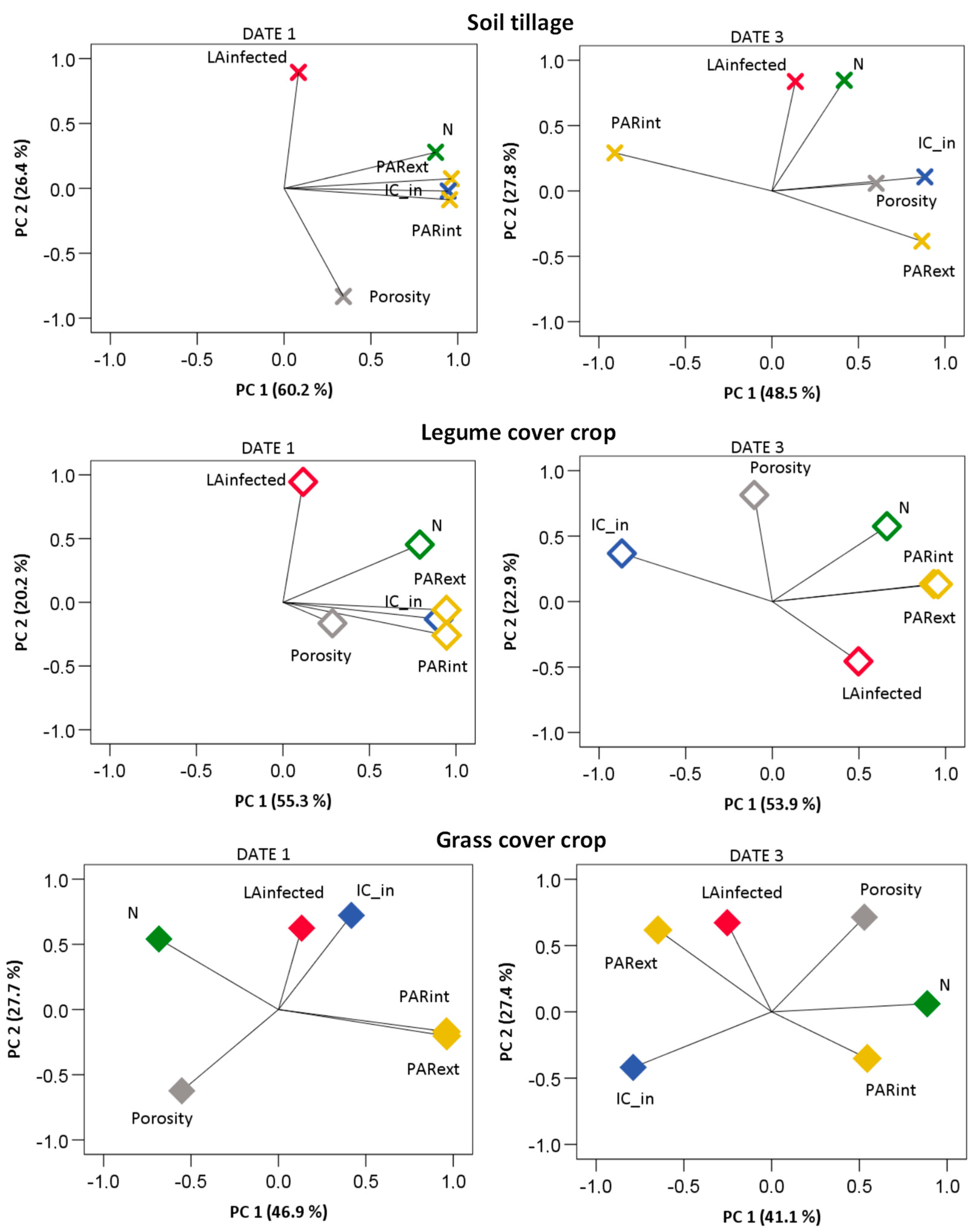
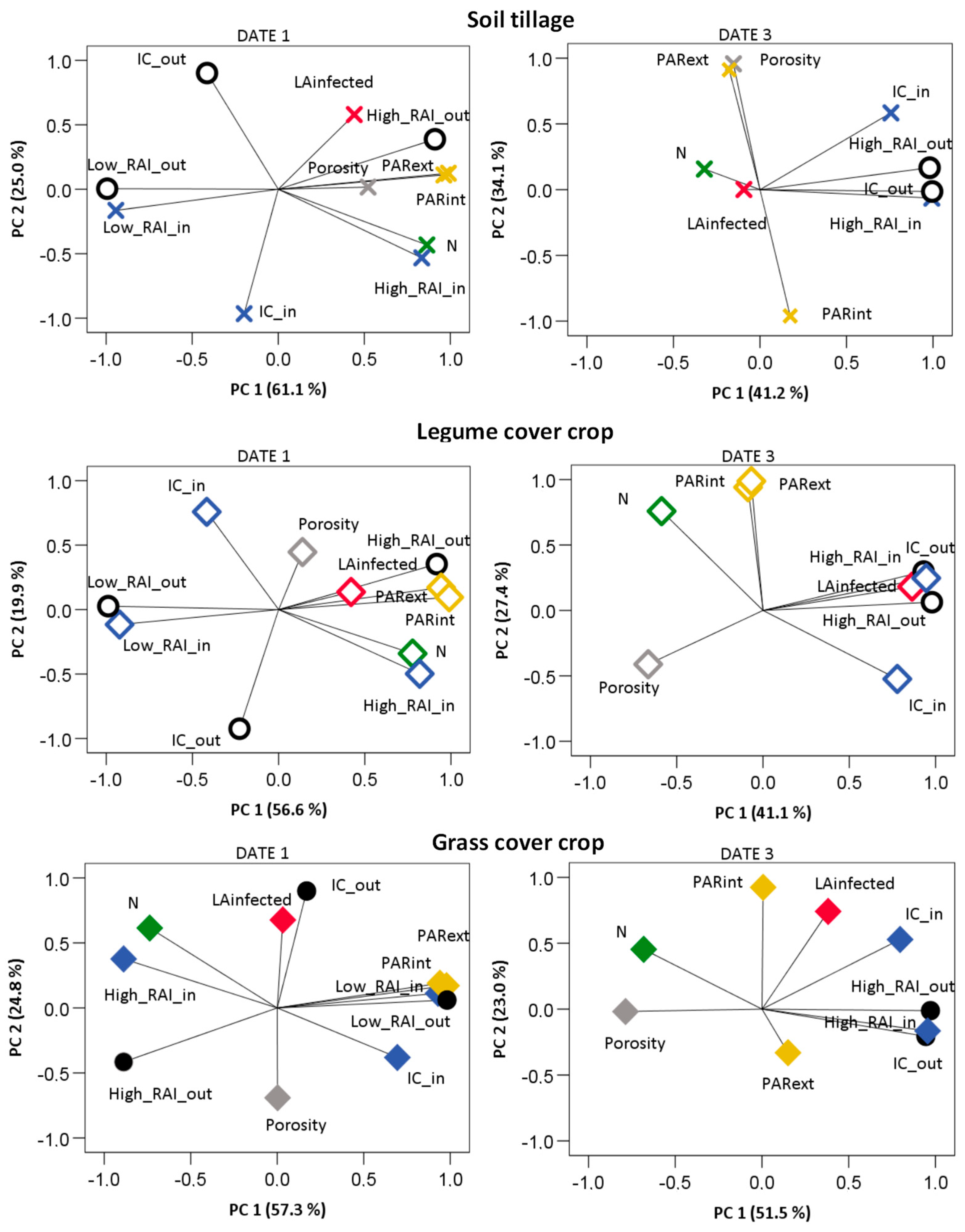
The multifactor analysis confirmed a weaker influence of the hygrothermal conditions inside the canopy compared to the growth, N, and canopy density factors on downy mildew development than that observed for powdery mildew. The variation in these variables was mostly explained by two principal components (PC), which represented, respectively, about 87, 80, and 75% of the whole variance in ST, LEG, and GRA in date 1 of the disease observations, and about 76, 77, and 69% by the last date of disease monitoring in grapevine shoots (
Figure 12,
Table A4). During the first observations of disease symptoms in ST, infected LA and porosity evolved along PC 2, while a favorable IC, canopy PAR levels, and N variations were explained by PC 1. In LEG plants, the infected LA variation was mostly related to N, along PC 2, while the other variables varied along PC 1. The infected LA of GRA varied with N along PC 2. Yet, also porosity and the weather conditions for infection inside the canopy were related with the infected LA in PC 2.
By the time the last disease symptoms were observed in the shoots, N variation had already assumed a strong relation with infected LA along PC 2 in ST, while the other factors varied along PC 1. Meanwhile, in LEG plants, the variation in LA infected by downy mildew was partially related to the light and hygrothermal conditions for infection inside the canopy, in PC 1, and with porosity and N along PC2. During last disease observations, the infected LA of GRA varied along PC 2 with porosity and high PAR levels at the external leaf layers, and also with the PAR levels inside the canopy along PC 3.
With regard to powdery mildew development, the canopy growth, leaf N, and microclimate variables showed a different relative importance on disease development in the three treatments, on the first and third monitoring dates (
Figure 13,
Table A5).
Overall, for the first observations of powdery mildew, about 86, 77, and 82 % of the variation in the treatments ST, LEG, and GRA were explained by two PCs. For ST, PC1 correlated the variation of infected LA with N, PAR, and the RAI classes both inside and outside the canopy, while PC 2 mostly grouped infected LA with IC inside and outside the canopy. Similar relationships were observed in LEG, but the infected LA also correlated with IC_out along the PC 2 and a third component related the infected LA with porosity and N. The importance of PAR interception in the disease pressure for ST canopies is consistent with an increase in disease severity in shaded canopies, as reported by Austin and Wilcox [
50], thus confirming the important role of light microclimate on the development of powdery mildew. In GRA plants, the infected LA correlated well with IC_out and porosity along PC2, but IC_in and all the outer weather and microclimate variables correlated poorly with the infected LA in the first stages of disease development. These results highlight the major role of the canopy temperature and humidity in the first steps of disease development in ST and LEG, while in GRA canopies the air atmospheric condition and low canopy density were the main factors affecting infections and disease pressure at its first stages of development, along the three years of the trial. As the disease evolved, the weight of the two principal components decreased slightly to 75% in ST, 69% in LEG, and 75% in GRA. The decreases were not caused by a weaker influence of canopy light, weather, or hygrothermal conditions for disease development, but were rather due to an increase in the relationship among the infected LA and N in ST (along a PC 3;
Table A5) and, conversely, in LEG, with a lower correlation of the PAR levels and N with disease development. Date 3 corresponds to an advanced stage of the cover-crop growth cycle, and it is probable that the nitrogen fixation activity had already decayed in the LEG cover crop. This result is consistent with those of Patrick et al. [
13], who observed the higher potential of a legume cover crop to supply N to the grapevine during the stages of intense canopy growth and until fruit set. Celette et al. [
51] also observed a stronger linkage between the cover crop and mild nitrogen stress in the vineyard during the early canopy growth stages. Regarding GRA, the infected LA correlated the most with the high levels of PAR at the inner canopy layers and also correlated with PAR in the outer canopy along PC3. Still, porosity and N correlated with infected LA along PC1, as well as with the hygrothermal conditions for disease development and pressure. It is likely that a decrease in pathogen activity occurred due to the higher exposure of the canopy to high solar radiation intensity, both in the inner and outer canopy layers of GRA, as observed also by Austin et al. [
48] in cluster layers. Our results also suggest that in GRA grapevines, both the leaf N decrease and higher canopy exposure may have led to higher dryness and hardness of leaf tissues (typical of older leaves), which are less attractive to the pathogen. In fact, in date 3, PC3 of the GRA treatment concerned mostly the PAR levels, evolving in a different direction from both canopy growth and disease development. Conversely, in ST at this stage, a strong relationship between the high leaf infection and N resulted in a third discriminant component, which brought the explained variability to 91%. Meanwhile, the affected LA of GRA was still highly correlated with the conditions for infections, to RAI and porosity along PC 1, and inversely correlated with the internal levels of PAR in PC 2.
Therefore, the low density of the GRA canopies allowed for the positive effect of higher sunlight levels, reducing both diseases’ incidence and pressure in the vines managed with a grass cover crop, while the impact of the legume’s nitrogen fixation on disease development in grapevine leaves of LEG was less important than the effect of nitrogen availability in the ST on grapevine susceptibility to both downy and powdery mildew.
Among the benefits of a grass cover crop in vineyard observed in this trial, the better canopy growth balance and microclimate had a positive effect on limiting downy and powdery mildew diseases. However, under arid environments and during hot and dry seasons, the use of permanent or total grass cover crops for long periods may lead to greater grapevine vigor reductions and an increased frequency of severe water-stress conditions, especially in sandy and/or low-fertility soils. In similar conditions, an efficient irrigation system that allows for appropriate irrigation supply, to counterbalance the competition for water and nutrients among the herbaceous cover and the vines, must be used [
11,
52].


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












