
Characterisation of the Complete Chloroplast Genomes of Seven Hyacinthus orientalis L. Cultivars: Insights into Cultivar Phylogeny

 , , ,
, , ,
Abstract
:1. Introduction
1.1. Taxonomy of Hyacinthus orientalis L.
1.1.1. Morphology
1.1.2. Nomenclature
1.2. Cultivation of Hyacinthus orientalis L.
1.2.1. History of Cultivation
1.2.2. Nomenclatural Circumscription of Hyacinthus Cultivars
1.3. Recent Molecular Insight into Hyacinthus Cultivars and the Potentiality of Chloroplast Genomes for Cultivar Phylogeny
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. Chloroplast Genome Sequencing, Assembly and Annotation
2.3. Sequence Repeat Analysis
2.4. Visualisation of the Boundary Variations
2.5. Comparative and Phylogenetic Analysis
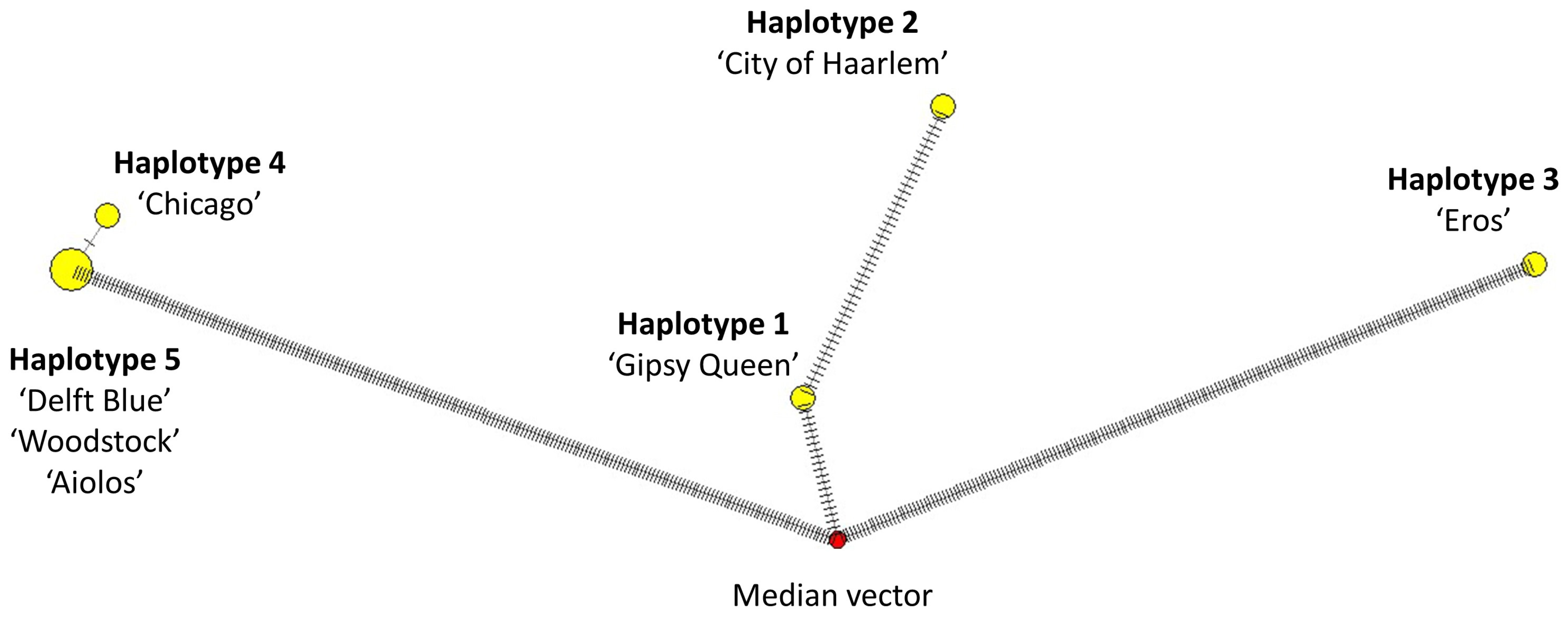
2.6. Haplotype Grouping and Identification of Molecular Diagnostic Characters
3. Results
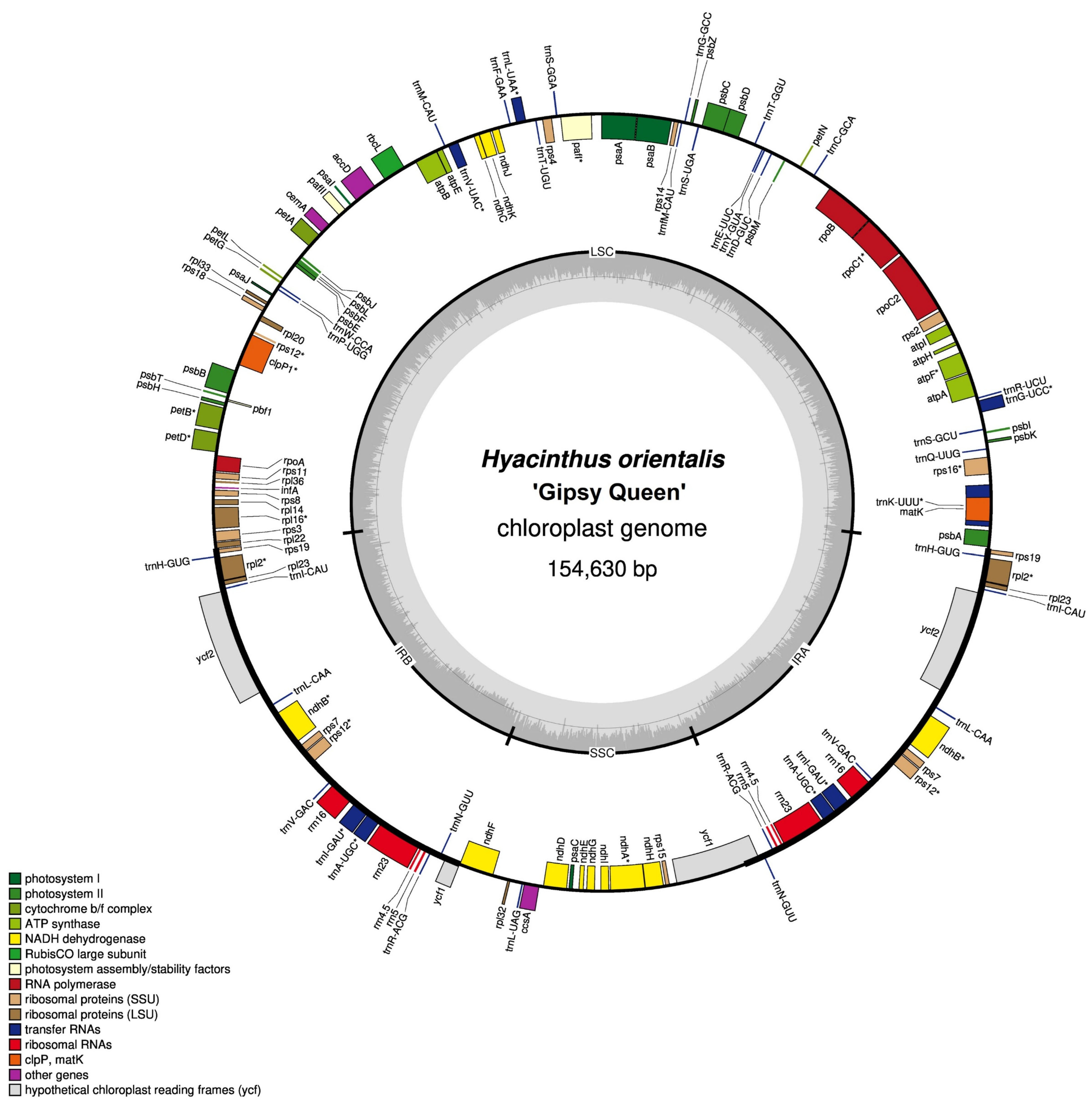
3.1. Features of Hyacinthus Chloroplast Genomes
3.1.1. Genome Size
3.1.2. Gene Number and Content
3.1.3. Guanine–Cytosine (GC) Content
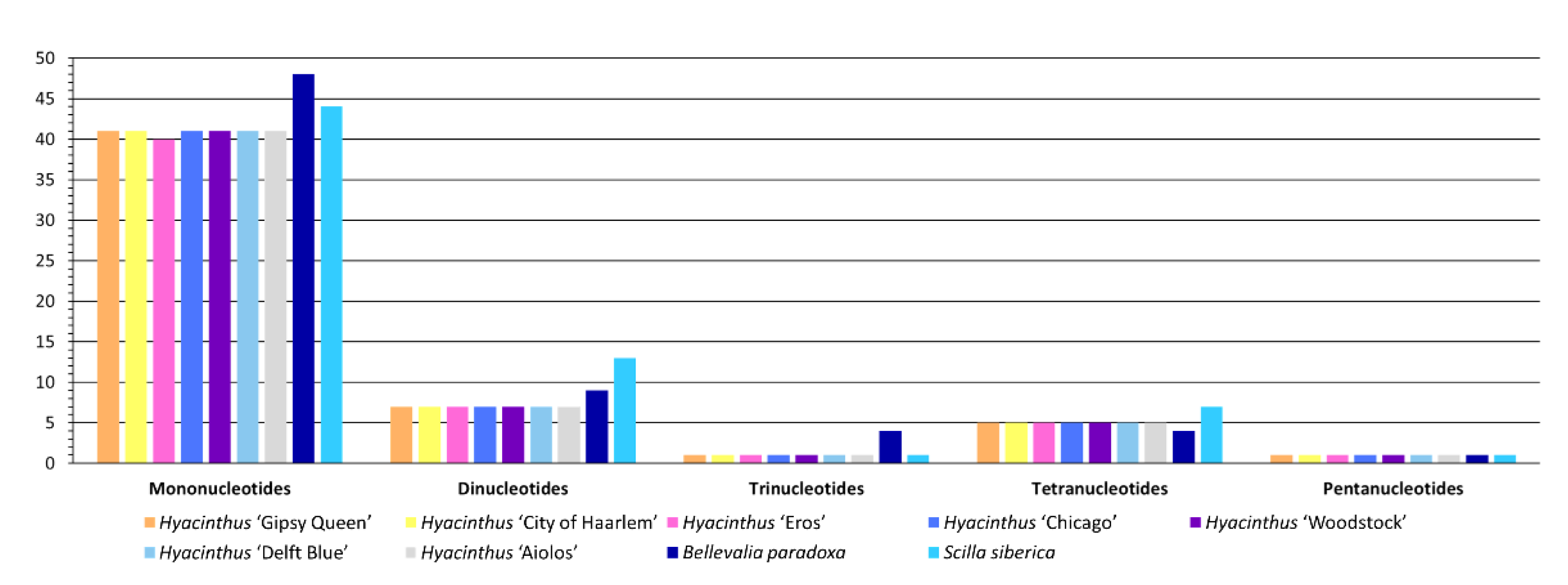
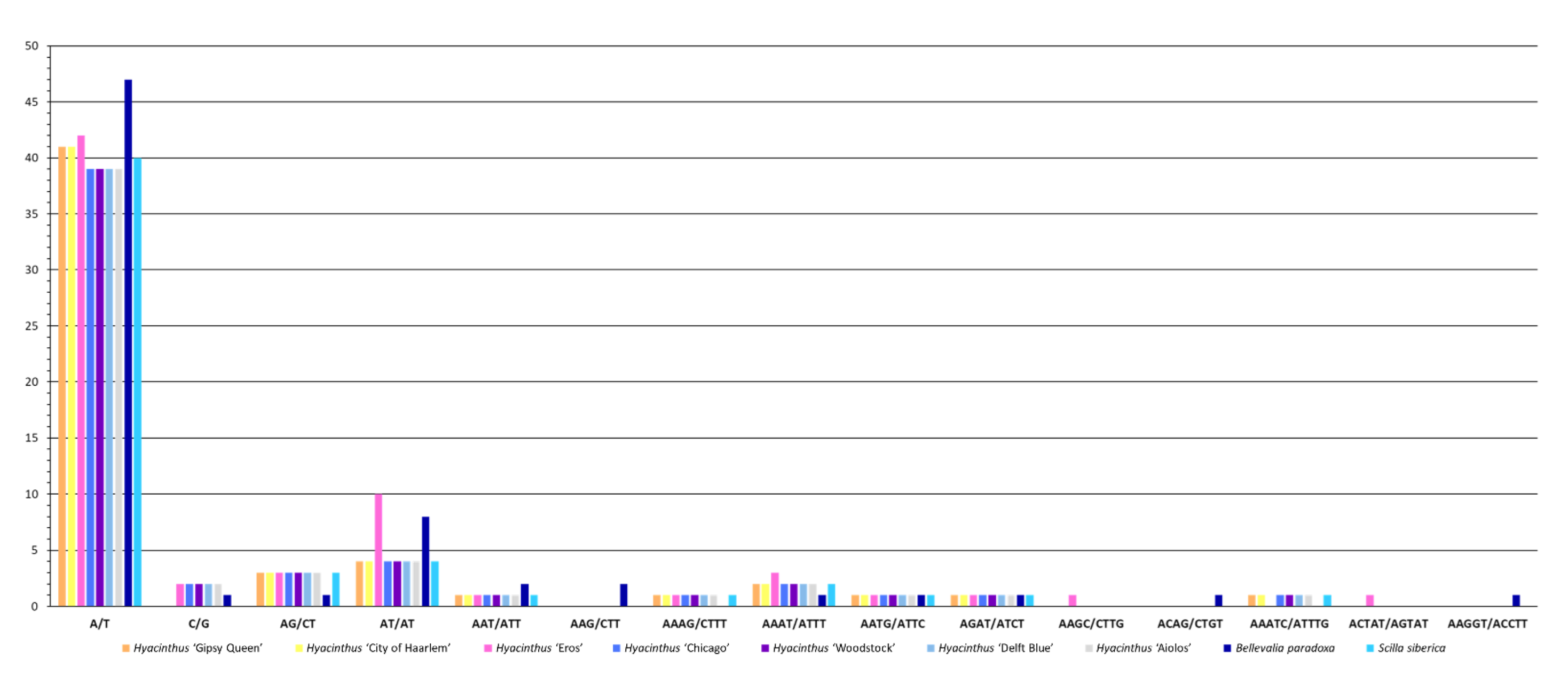
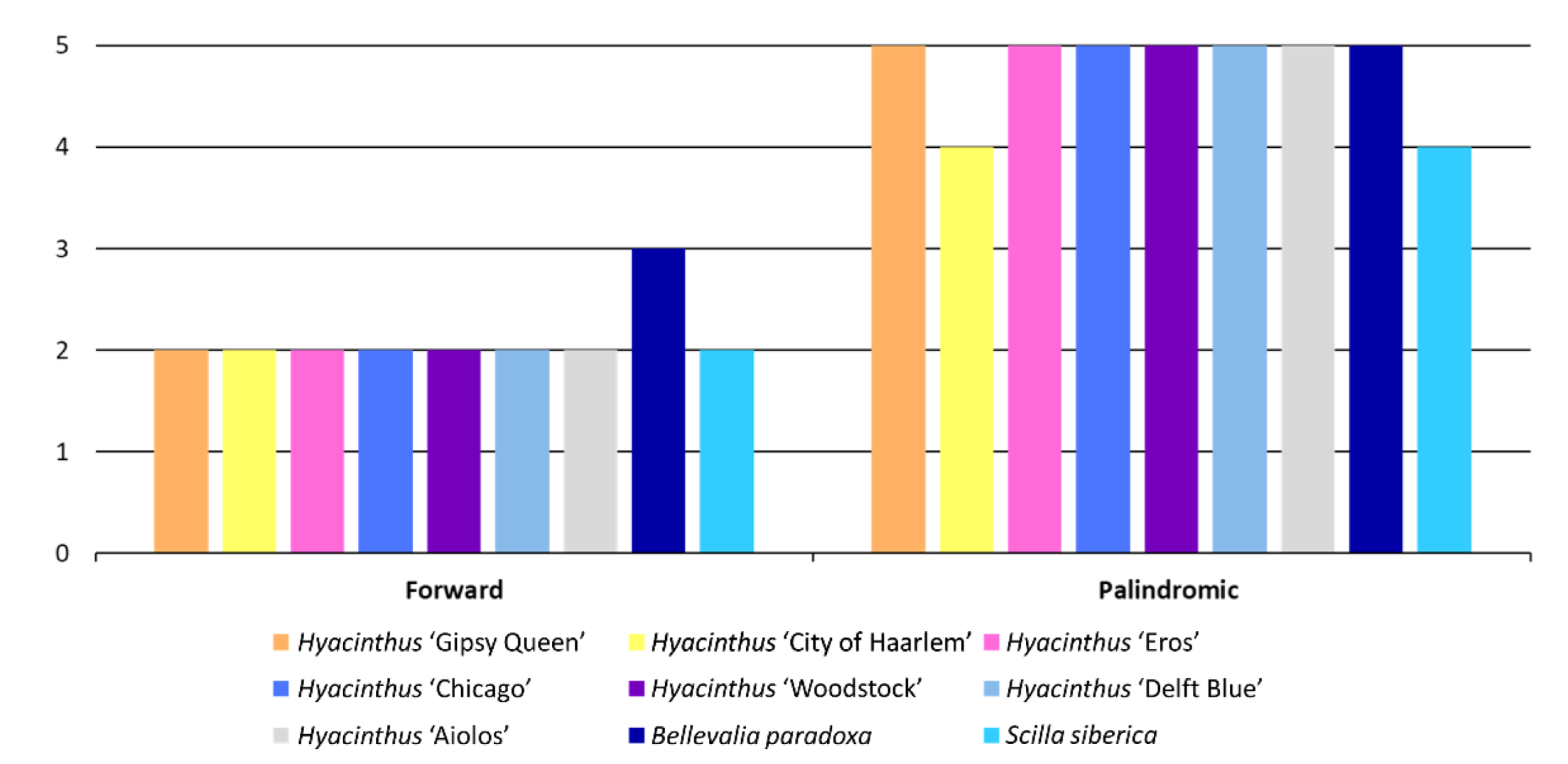
3.2. Sequence Repeats
3.2.1. Simple Sequence Repeats (SSRs)
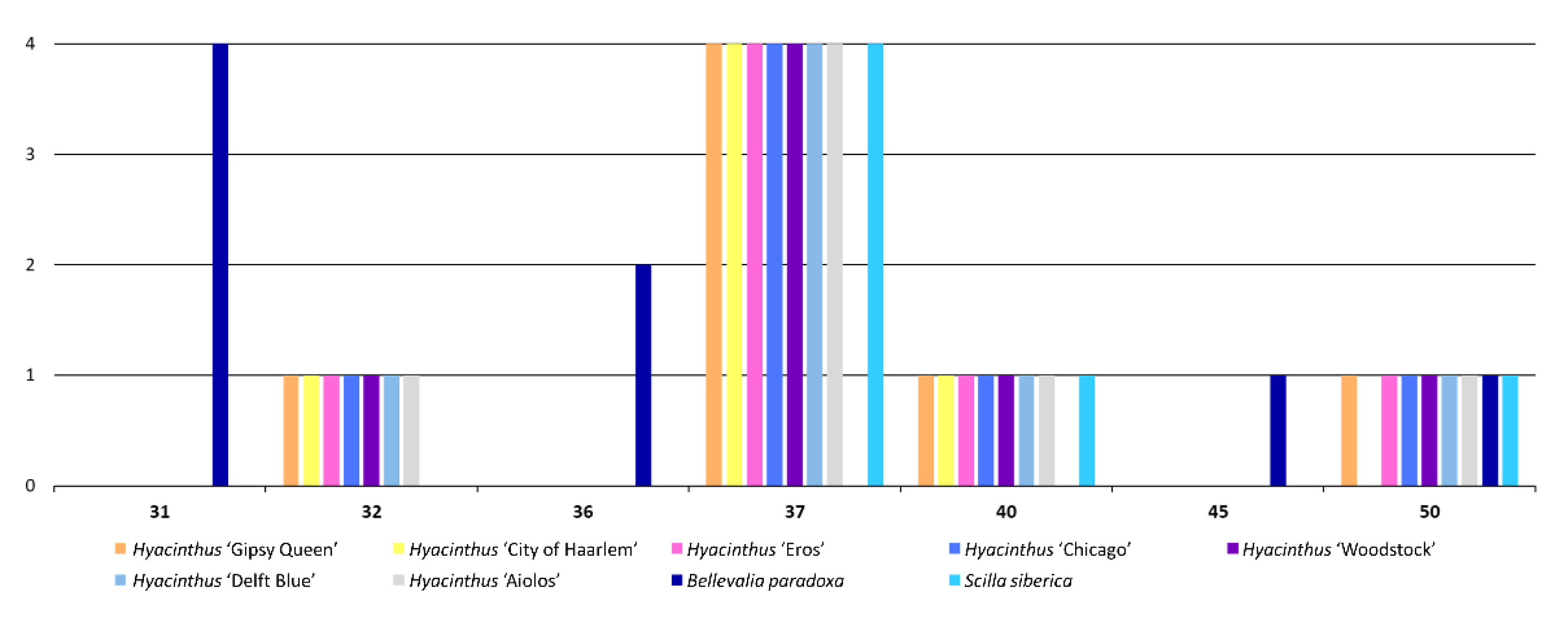
3.2.2. Long Tandem Repeats (LTRs)
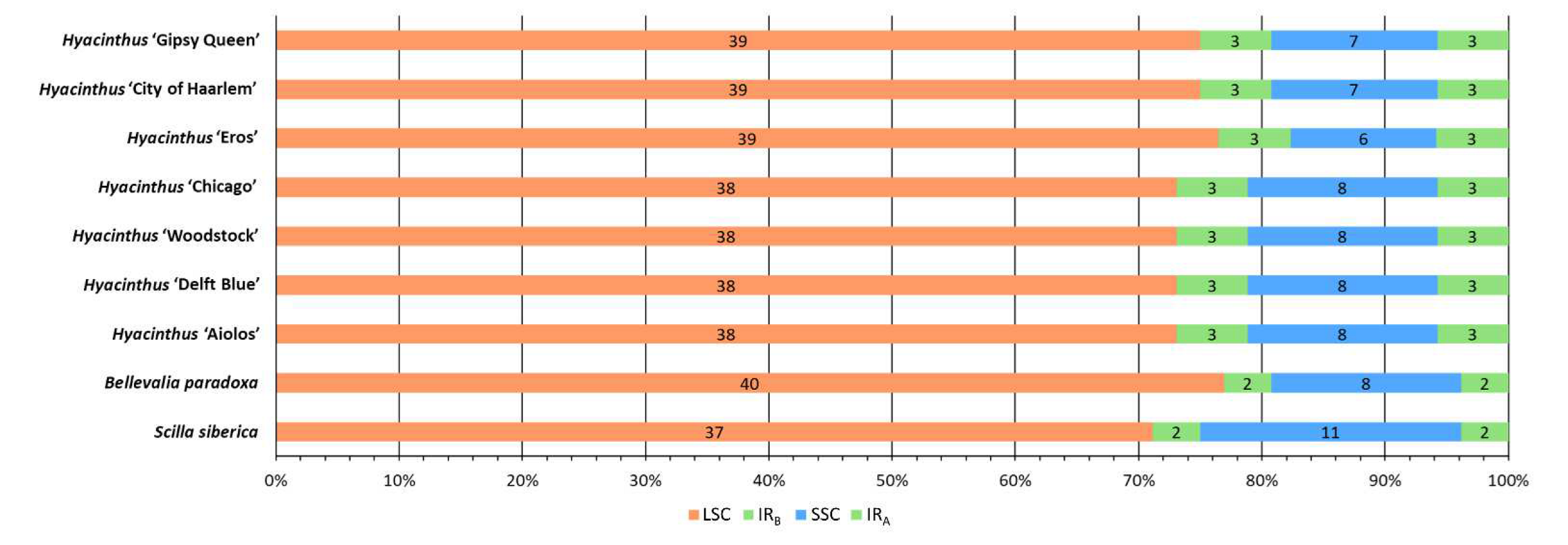
3.3. Boundary Variation
3.4. Divergence Hotspots
3.5. Phylogenetic Analysis
3.6. Haplotype Grouping and Identification of Molecular Diagnostic Characters
4. Discussion
4.1. Chloroplast Genomes Serve as New Evidence in the Phylogeny of Hyacinthus cultivars
4.2. Timeline of Colour Evolution and Phylogenetic Relationship of Hyacinthus Cultivars
4.3. Differences among the Chloroplast Genomes of Hyacinthus Cultivars
4.4. Understanding the Phylogenetic Relationship Contributes to Cultivar Breeding
4.5. Future Direction of Hyacinthus Cultivar Phylogeny
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yeo, P.F. Hyacinthus Linnaeus. In The European Garden Flora, 1st ed.; Walters, S.M., Brady, A., Brickell, C.D., Cullen, J., Green, P.S., Lewis, J., Matthews, V.A., Webb, D.A., Yeo, P.F., Alexander, J.C.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA; Melbourne, Australia, 1986; Volume I, p. 221. ISBN 0-521-24859-0. (hardback). [Google Scholar]
- Shen, Q.; Wang, H.; Zhu, H.X.; Gu, J.J. Tulips and Hyacinths, 1st ed.; China Forestry Press: Beijing, China, 2004; ISBN 9787503836473. [Google Scholar]
- Çığ, A.; Başdoğan, G. In vitro Propagation Techniques for Some Geophyte Ornamental Plants with High Economic Value. Int. J. Second. Metab. 2015, 2, 27–49. [Google Scholar]
- Bailey, L.H.; Bailey, E.Z. Hyacinthus L. In Hortus Third: A Concise Dictionary of Plants Cultivated in the United States and Canada, 1st ed.; Macmillian Publishing Company: New York, NY, USA; Collier Macmillan Publisher: London, UK, 1976; p. 577. [Google Scholar]
- The Royal Horticultural Society. The New Royal Horticultural Society Dictionary of Gardening, 1st ed.; Huxley, A., Griffiths, M., Levy, M., Eds.; The Macmillan Press Limited: London, UK; The Stockton Press: New York, NY, USA, 1992; pp. 604–605. ISBN 1-56159-001-0. [Google Scholar]
- The Royal Horticultural Society. The Royal Horticultural Society Plant Guides: Bulbs, 1st ed.; Dorling Kindersley Publishers Ltd: London, UK, 1997; ISBN 9780751303056. [Google Scholar]
- Nowak, J.; Rudnicki, R.M. Hyacinthus. In The Physiology of Flower Bulbs, 1st ed.; de Hertogh, A., le Nard, M., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 335–347. ISBN 0-444-87498-4. [Google Scholar]
- Hu, F.R.; Liu, H.H.; Wang, F.; Bao, R.L.; Liu, G.X. Root tip chromosome karyotype analysis of hyacinth cultivars. Genet. Mol. Res. 2015, 14, 10863–10876. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.R.; Hu, Y.M.; Wang, F.; Ren, C. Genetic Diversity of 29 Hyacinth Germplasm Resources Revealed by Using ISSR Markers. Mol. Plant Breed. 2015, 13, 379–385. [Google Scholar] [CrossRef]
- Conran, J. Family Hyacinthaceae. In Horticultural Flora of South-Eastern Australia, 1st ed.; Spencer, R., Ed.; University of New South Wales Press Ltd: Sydney, Australia, 2005; Volume 5, pp. 349–367. ISBN 0868408328. [Google Scholar]
- Hu, F.R.; Ren, C.; Bao, R.L.; Liu, G.X. Chromosomes analysis of five diploid garden Hyacinth species. Sci. Hortic. 2011, 13, 82–87. [Google Scholar] [CrossRef]
- Danin, A.; Fragman-Sapir, O. Hyacinthus orientalis L. In Flora of Israel Online. 2016. Available online: https://flora.org.il/en/plants/hyaori/ (accessed on 31 January 2022).
- Hickey, M.; King, C. The Cambridge Illustrated Glossary of Botanical Terms, 1st ed.; Cambridge University Press: Cambridge, UK, 2000; ISBN1 ISBN-13: 978-0-521-79401-5. ISBN2 ISBN-10: 0-521-79401-3. [Google Scholar]
- Saint-Simon, M.H. Des Jacintes de Leur Anatomie Reproduction et Culture, 1st ed.; De Límprimerie de C. Eel: Amsterdam, The Netherlands, 1768. [Google Scholar] [CrossRef] [Green Version]
- Beentjie, H. The Kew Plant Glossary, 2nd ed.; Kew Publishing: Surrey, UK, 2016; ISBN 978-1-84246-604-9. [Google Scholar]
- Kersten, H.J.H. The cultivation of the hyacinth in Holland. J. R. Hortic. Soc. Lond. 1889, 11, 54–63. Available online: https://www.biodiversitylibrary.org/item/164536 (accessed on 31 January 2022).
- Zhao, Z.Z.; Yang, P.P.; Chen, Q.Q.; Yu, J.J.; Wu, C.Z. Study on Phenophase and Growth Characteristics of 12 Cultivars of Hyacinthus orientalis. J. Kashi Univ. 2019, 40, 111–115. [Google Scholar] [CrossRef]
- Pasztor, R.; Bala, M.; Sala, F. Flowers quality in relation to planting period in some hyacinth cultivars. AgroLife Sci. J. 2020, 9, 263–272. Available online: http://agrolifejournal.usamv.ro/pdf/vol.IX_1/Art33.pdf (accessed on 31 January 2022).
- Van Scheepen, J. International Checklist for Hyacinths and Miscellaneous Bulbs; Royal General Bulbgrowers’ Association: Hillegom, The Netherlands, 1991; ISBN 90-73350-01-8. [Google Scholar]
- Darlington, C.; Hair, J.; Hurcombe, R. The history of the garden hyacinths. Heredity 1951, 5, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Wendelbo, P. Hyacinthus L. In Flora of Turkey and the East Aegean Islands, 1st ed.; Davis, P., Mill, R., Tan, K., Eds.; Edinburgh University Press: Edinburgh, UK, 1984; Volume 8, pp. 263–264. ISBN 0-85224-494-0. [Google Scholar]
- Horovitz, A.; Danin, A. Relatives of ornamental plants in the flora of Israel. Isr. J. Plant Sci. 1983, 32, 75–95. Available online: https://flora.org.il/en/articles/relatives-of-ornamental-plants-in-the-flora-of-israel-2/ (accessed on 31 January 2022).
- Linnaei, C. Hyacinthus. In Species Plantarum, 1st ed.; Impensis Laurentii Salvii: Holmiae, Sweden, 1753; Tomus I; pp. 316–318. [Google Scholar] [CrossRef] [Green Version]
- Missouri Botanical Garden. Hyacinthaceae Batsch ex Borkh. 2022. Available online: https://tropicos.org/name/50324572 (accessed on 31 January 2022).
- Missouri Botanical Garden. Hyacinthus L. 2022. Available online: https://www.tropicos.org/name/40028561 (accessed on 31 January 2022).
- Borkhausen, M.B. Hyacinthinae Batsch. In Botanisches Wörterbuch, 1st ed.; G.F. Heyer: Giessen, Germany, 1797; Volume 1, p. 315. [Google Scholar] [CrossRef] [Green Version]
- Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F.; Faden, R.B.; Jacobsen, N.; Jakobsen, K.; Jensen, S.R.; Nielsen, B.J.; Rasmussen, F.N. Hyacinthaceae Batsch (1802). In The Families of the Monocotyledons: Structure, Evolution, and Taxonomy, 1st ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Tokyo, Japan, 1985; pp. 188–193. ISBN 3-540-13655-X. [Google Scholar]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 2003, 141, 399–436. [Google Scholar] [CrossRef] [Green Version]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Chase, M.W.; Reveal, J.L.; Fay, M.F. A subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Bot. J. Linn. Soc. 2009, 161, 132–136. [Google Scholar] [CrossRef] [Green Version]
- McNeill, J.; Barrie, F.R.; Burdet, H.M.; Demoulin, V.; Hawksworth, D.L.; Marhold, K.; Nicolson, D.H.; Prado, J.; Silva, P.C.; Skog, J.E.; et al. International code of botanical nomenclature—Vienna Code. Regnum Veg. 2006, 146, 29–32. Available online: https://www.iapt-taxon.org/historic/2006.htm (accessed on 31 January 2022).
- Link, J.H.F. Subordo 2. Hyacinthinae. Hyacinthartige. In Handbuch zur Erkennung der Nutzbarsten und am Häufigsten Vorkommenden Gewächse, 1st ed.; Spenerschen Buchhandlung: Berlin, Germany, 1829; p. 160. [Google Scholar] [CrossRef] [Green Version]
- Burnett, G.T. Scillidae. In Outlines of Botany, including a General History of the Vegetable Kingdom. In which Plants Are Arranged According to the System of Natural Affinities; John Churchill: London, UK, 1835; Volume 1, p. 428. Available online: https://archive.org/details/b28742631_0001/page/428/mode/2up (accessed on 31 January 2022).
- Douglas, J. The Hyacinth from an English Point of View. J. R. Hortic. Soc. Lond. 1889, 11, 63–70. Available online: https://www.biodiversitylibrary.org/item/164536 (accessed on 31 January 2022).
- International Society for Horticultural Science. ICRA Report Sheet. Royal General Bulbgrowers’ Association (K.A.V.B.). Available online: https://www.ishs.org/sci/icralist/49.htm (accessed on 31 January 2022).
- International Society for Horticultural Science. International Society for Horticultural Science. International Code of Nomenclature for Cultivated Plants (ICNCP). In Scripta Horticulturae, 9th ed.; International Society for Horticultural Science: Leuven, Belgium, 2016; Volume 18, ISBN1 978-94-6261-116-0. Available online: https://www.ishs.org/sites/default/files/static/ScriptaHorticulturae_18.pdf (accessed on 31 January 2022)ISBN2 978-94-6261-116-0.
- Koninklijke Algemeene Vereeniging voor Bloembollencultuur; Search/Zoekopdracht: Hyacinthus, Zoekresultaten. Available online: https://www.kavb.nl/zoekresultaten (accessed on 31 January 2022).
- Darlington, C.D.; Mather, K. Chromosome balance and interaction in Hyacinthus. J. Genet. 1944, 46, 52–61. [Google Scholar] [CrossRef]
- Brat, S. Fertility and selection in hyacinth I. Gametic selection. Heredity 1967, 22, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Brat, S. Fertility and selection in garden hyacinth II. Zygotic selection. Heredity 1969, 24, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.R.; He, G.R.; Wang, F.; Ren, C. The Identification of the Hyacinth Hybrid Progeny by the Method of ISSR Molecular Marker. Mol. Plant Breed. 2015, 13, 1336–1342. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Muller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Leonl, S.; Gernandt, D.S.; Perez de la Rosa, J.A.; Jardon-Barbolla, L. Phylogenetic relationships and species delimitation in Pinus section Trifoliae inferred from plastid DNA. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Li, X.W.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.T.; Chen, S.L. Plant DNA barcoding: From gene to genome. Biol. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Williams, A.V.; Miller, J.T.; Small, I.; Nevill, P.G.; Boykin, L.M. Integration of complete chloroplast genome sequences with small amplicon datasets improves phylogenetic resolution in Acacia. Mol. Phylogenet. Evol. 2016, 96, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.P.; Bi, Y.; Yang, F.P.; Zhang, M.F.; Chen, X.Q.; Xue, J.; Zhang, X.H. Complete chloroplast genome sequences of Lilium: Insights into evolutionary dynamics and phylogenetic analyses. Sci. Rep. 2017, 7, 5751. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, S.I.; Kim, B.R.; Choi, I.Y.; Ryser, P.; Kim, N.S. Chloroplast genomes of Lilium lancifolium, L. amabile, L. callosum, and L. philadelphicum: Molecular characterization and their use in phylogenetic analysis in the genus Lilium and other allied genera in the order Liliales. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef]
- Li, W.; Zhang, C.; Guo, X.; Liu, Q.; Wang, K. Complete chloroplast genome of Camellia japonica genome structures, comparative and phylogenetic analysis. PLoS ONE 2019, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Dong, W.; Li, W.; Lu, Y.; Xie, X.; Jin, X.; Shi, J.; He, K.; Suo, Z. Comparative Analysis of Six Lagerstroemia Complete Chloroplast Genomes. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tan, W.; Sun, J.; Du, J.H.; Zheng, C.G.; Tian, X.X.; Zheng, M.; Xiang, B.B.; Wang, Y. Comparison of Four Complete Chloroplast Genomes of Medicinal and Ornamental Meconopsis Species: Genome Organization and Species Discrimination. Sci. Rep. 2019, 9, 10567. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.J.; Zhang, K.; Peng, Z.F.; Sun, S.S.; Ya, H.Y.; Zhang, Y.Z.; Cheng, Y.W. Comparative Analysis of Chloroplast Genome Characteristics between Paeonia jishanensis and Other Five Species of Paeonia. Lin Ye Ke Xue 2020, 56, 82–88. Available online: http://www.linyekexue.net/EN/10.11707/j.1001-7488.20200409 (accessed on 31 January 2022).
- Chen, J.L.; Zhou, Y.Z.; Zhao, K. Chloroplast genome of Prunus campanulata ‘Fei han’ (Rosaceae), a new cultivar in the modern cherry breeding. Mitochondrial DNA Part B Resour. 2020, 5, 1369–1371. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Xu, Y.; Liao, B.; Huang, Z.; Zhou, H.; Wang, F. Complete chloroplast genome sequence of a Chinese traditional cultivar in Chrysanthemum, Chrysanthemum morifolium ’Anhuishiliuye’. Mitochondrial DNA B Resour. 2021, 6, 1281–1282. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.H.; Gao, J.; Li, M.; Wu, X.; Shen, C.; Wu, J.; Qi, Z.C.; Li, P. The complete chloroplast genome sequences of Barnardia japonica (Thunb.) Schult. and Schult.f. Mitochondrial DNA B Resour. 2018, 3, 697–698. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, Y.; Chen, J.; Lin, N.; Xia, M. The complete plastome and phylogeny of Barnardia japonica (Asparagaceae). Mitochondrial DNA B Resour. 2020, 5, 2640–2641. [Google Scholar] [CrossRef] [PubMed]
- Garnett, G.J.L.; Könyves, K.; Bilsborrow, J.; David, J.; Culham, A. The complete plastome of Hyacinthoides non-scripta (L.) Chouard ex Rothm. (Asparagaceae). Mitochondrial DNA B Resour. 2020, 5, 1003–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKain, M.R.; McNeal, J.R.; Kellar, P.R.; Eguiarte, L.E.; Pires, J.C.; Leebens-Mack, J. Timing of rapid diversification and convergent origins of active pollination within Agavoideae (Asparagaceae). Am. J. Bot. 2016, 103, 1717–1729. [Google Scholar] [CrossRef] [PubMed]
- Wendelbo, P. Bellevalia Lapeyr. In Flora of Turkey and the East Aegean Islands, 1st ed.; Davis, P., Mill, R., Tan, K., Eds.; Edinburgh University Press: Edinburgh, UK, 1984; Volume 8, pp. 264–274. ISBN 0-85224-494-0. [Google Scholar]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The Manifold Applications of Repeat Analysis on a Genomic Scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32 (Suppl. 2), W273–W279. [Google Scholar] [CrossRef]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal Alignment: Finding Rearrangements During Alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sánchez-Gracia, A. DNA Sequence Polymorphism (DnaSP). Version: 6.12.03 (26 February 2019) [software]. Universitat de Barcelona. Available online: http://www.ub.edu/dnasp/ (accessed on 31 January 2022).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Free Phylogenetic Network Software. Available online: https://www.fluxus-engineering.com/sharenet.htm (accessed on 26 April 2022).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Barnaart, H.A.E. Historical notes on Dutch hyacinths. J. R. Hortic. Soc. Lond. 1889, 11, 49–54. Available online: https://www.biodiversitylibrary.org/item/164536 (accessed on 31 January 2022).
- Gerard, J. Hyacinthus orientalis. In The Herball, or, Generall Historie of Plantes, 1st ed.; John Norton: London, UK, 1597; p. 100. Available online: https://archive.org/details/mobot31753000817749/page/100/mode/2up (accessed on 31 January 2022).
- Palmer, J.D.; Jansen, R.K.; Michaels, H.J.; Chase, M.W.; Manhart, J.R. Chloroplast DNA Variation and Plant Phylogeny. Ann. Mo. Bot. Gard. 1988, 75, 1180–1206. [Google Scholar] [CrossRef]
- Wu, L.L.; Li, J.A.; Gu, Y.Y.; Zhang, F.H.; Li, Z.; Tan, X.F. Complete Chloroplast Genome Sequences and Phylogenetic Analysis of Three Camellia oleifera Cultivars. Int. J. Agric. Biol. 2020, 24, 743–750. Available online: https://www.fspublishers.org/html_issue.php?i_id=41338 (accessed on 31 January 2022).
- Kim, K.J.; Lee, H.L. Complete chloroplast genome sequences from Korean ginseng (Panax schinseng Nees) and comparative analysis of sequence evolution among 17 vascular plants. DNA Res. 2004, 11, 247–261. [Google Scholar] [CrossRef]
- Asaf, S.; Waqas, M.; Khan, A.L.; Khan, M.A.; Kang, S.M.; Imran, Q.M.; Shahzad, R.; Bilal, S.; Yun, B.W.; Lee, I.J. The complete chloroplast genome of wild rice (Oryza minuta) and its comparison to related species. Front. Plant Sci. 2017, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.L.; Cheng, L.L.; Huang, W.G.; Cao, Q.C.; Zhou, L.; Jia, W.S.; Lan, Y.P. Chloroplast genomes of seven species of Coryloideae (Betulaceae): Structures and comparative analysis. Genome 2020, 63, 337–348. [Google Scholar] [CrossRef]
- Hu, G.; Wang, Y.; Wang, Y.; Zheng, S.; Dong, W.; Dong, N. New Insight into the Phylogeny and Taxonomy of Cultivated and Related Species of Crataegus in China, Based on Complete Chloroplast Genome Sequencing. Horticulturae 2021, 7, 301. [Google Scholar] [CrossRef]
- Hong, S.Y.; Cheon, K.S.; Yoo, K.O.; Lee, H.O.; Cho, K.S.; Suh, J.T.; Kim, S.J.; Nam, J.H.; Sohn, H.B.; Kim, Y.H. Complete Chloroplast Genome Sequences and Comparative Analysis of Chenopodium quinoa and C. album. Front Plant Sci 2017, 8, 1696. [Google Scholar] [CrossRef]
- Xu, W.B.; Xia, B.S.; Li, X.W. The complete chloroplast genome sequences of five pinnate-leaved Primula species and phylogenetic Analyses. Sci. Rep. 2020, 10, 20782. [Google Scholar] [CrossRef]
- Silva, S.R.; Pinheiro, D.G.; Penha, H.A.; Płachno, B.J.; Michael, T.P.; Meer, E.J.; Miranda, V.F.O.; Varani, A.M. Intraspecific Variation within the Utricularia amethystine Species Morphotypes Based on Chloroplast Genomes. Int. J. Mol. Sci. 2019, 20, 6130. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Shiokai, S.; Yamaguchi, M.; Kishitani, S.; Nishio, T. Dot-blot-SNP analysis for practical plant breeding and cultivar identification in rice. Theor. Appl. Genet. 2006, 13, 147–155. [Google Scholar] [CrossRef]
- Sato, H.; Endo, T.; Shiokai, S.; Nishio, T.; Yamaguchi, M. Identification of 205 current rice cultivars in Japan by dot-blot-SNP analysis. Breed Sci. 2010, 60, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.K.; Park, S.W.; Liu, W.Y.; Kang, B.C. Discovery of single nucleotide polymorphism in Capsicum and SNP markers for cultivar identification. Euphytica 2010, 175, 91–107. [Google Scholar] [CrossRef]
- Liu, W.; Xiao, Z.; Bao, X.; Yang, X.; Fang, J.; Xiang, X. Identifying Litchi (Litchi chinensis Sonn.) Cultivars and Their Genetic Relationships Using Single Nucleotide Polymorphism (SNP) Markers. PLoS ONE 2015, 10, e0135390. [Google Scholar] [CrossRef] [Green Version]
- Jakse, J.; Martin, W.; McCallum, J.; Havey, M.J. Single Nucleotide Polymorphisms, Indels, and Simple Sequence Repeats for Onion Cultivar Identification. J. Am. Soc. Hortic. Sci. 2005, 130, 912–917. [Google Scholar] [CrossRef]
- Wu, K.; Yang, M.; Liu, H.; Tao, Y.; Mei, J.; Zhao, Y. Genetic analysis and molecular characterization of Chinese sesame (Sesamum indicum L.) cultivars using insertion-deletion (InDel) and simple sequence repeat (SSR) markers. BMC Genet. 2014, 19, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, H.B.; Kim, S.J.; Hwang, T.Y.; Park, H.M.; Lee, Y.Y.; Markkandan, K.; Lee, D.; Lee, S.; Hong, S.Y.; Song, Y.H.; et al. Barcode System for Genetic Identification of Soybean [Glycine max (L.) Merrill] Cultivars Using InDel Markers Specific to Dense Variation Blocks. Front. Plant Sci. 2017, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, T.; Daiou, K.; Mihara, T.; Nagano, Y. Potential application of simple easy-to-use insertion-deletion (InDel) markers in citrus cultivar identification. Breed Sci. 2021, 71, 601–608. [Google Scholar] [CrossRef]
- Jafari, A.; Vaezi, J.; Forghanifard, M.M.; Forest, F.; Manning, J.C. A revised infrageneric classification of Bellevalia Lapeyr. (Asparagaceae: Scilloideae) based on molecular analysis. Phytotaxa 2021, 525, 70–84. [Google Scholar] [CrossRef]
- Wainwright, M. After 16 years of seed testing, the new breed of black hyacinth reveals appeal of dark side of the bloom. In Gardens, UK News, The Guardian, 22 March 2005. Available online: https://www.theguardian.com/uk/2005/mar/22/homesandgardens.shopping (accessed on 31 January 2022).
- Shoub, J.; de Hertogh, A.A. Floral stalk topple: A disorder of Hyacinthus orientalis L. and its control. HortScience 1975, 10, 26–28. [Google Scholar]
- Statistics Netherlands (CBS). Agriculture; crops, livestock and land use by general farm type, region. In StatLine—Electronic Database of Statistics Netherlands. Available online: https://opendata.cbs.nl/#/CBS/en/dataset/80783eng/barv?ts=1635259886355 (accessed on 31 January 2022).
- Steele, P.R.; Pires, J.C. Biodiversity assessment: State-of-the-art techniques in phylogenomics and species identification. Am. J. Bot. 2011, 98, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Malé, P.G.; Bardon, L.; Besnard, G.; Coissac, E.; Delsuc, F.; Engel, J.; Lhuillier, E.; Scotti-Saintagne, C.; Tinaut, A.; Chave, J. Genome skimming by shotgun sequencing helps resolve the phylogeny of a pantropical tree family. Mol. Ecol. Resour. 2014, 14, 966–975. [Google Scholar] [CrossRef]
- Gardner, E.M.; Johnson, M.G.; Ragone, D.; Wickett, N.J.; Zerega, N.J. Low-coverage, whole-genome sequencing of Artocarpus camansi (Moraceae) for phylogenetic marker development and gene discovery. Appl. Plant Sci. 2016, 4, 1600017. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Kobayashi, N.; Nakatsuka, A.; Ohta, H.; Isobe, S. Whole-genome sequencing and analysis of two azaleas, Rhododendron ripense and Rhododendron kiyosumense. DNA Res. 2021, 28, dsab010. [Google Scholar] [CrossRef]
- Straub, S.C.; Parks, M.; Weitemier, K.; Fishbein, M.; Cronn, R.C.; Liston, A. Navigating the tip of the genomic iceberg: Next-generation sequencing for plant systematics. Am. J. Bot. 2012, 99, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Blischak, P.D.; Wenzel, A.J.; Wolfe, A.D. Gene prediction and annotation in Penstemon (Plantaginaceae): A workflow for marker development from extremely low-coverage genome sequencing. Appl. Plant Sci. 2014, 2, 1400044. [Google Scholar] [CrossRef]
- Pryer, K.M.; Schneider, H.; Zimmer, E.A.; Banks, J.A. Deciding among green plants for whole genome studies. Trends Plant Sci 2002, 7, 550–554. [Google Scholar] [CrossRef]
- Kury, L.T.A.; Taha, Z.; Talib, W.H. Immunomodulatory and Anticancer Activities of Hyacinthus orientalis L.: An In Vitro and In Vivo Study. Plants 2021, 10, 617. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collector No. | Inventory No. | Sheet No. | Date of Collection | NCBI Accession No. | Raw Data (GB) | Coverage |
|---|---|---|---|---|---|---|---|
| Hyacinthus orientalis L. ‘Gipsy Queen’ | K. H. Wong 127 | CUSLSH2896 | CUHK05312 | 27 January 2021 | OM320803 | 3.2 | 308× |
| Hyacinthus orientalis L. ‘City of Haarlem’ | K. H. Wong 139 | CUSLSH2915 | CUHK05574 | 11 February 2021 | OM320804 | 3.5 | 203× |
| Hyacinthus orientalis L. ‘Eros’ | K. H. Wong 144 | CUSLSH2922 | CUHK05309 | 15 February 2021 | OM320805 | 4.1 | 194× |
| Hyacinthus orientalis L. ‘Chicago’ | K. H. Wong 047 | CUSLSH2460 | CUHK05305 | 22 January 2020 | OM320806 | 3.7 | 138× |
| Hyacinthus orientalis L. ‘Woodstock’ | K. H. Wong 114 | CUSLSH2881 | CUHK05317 | 22 January 2021 | OM320807 | 3.5 | 279× |
| Hyacinthus orientalis L. ‘Delft Blue’ | K. H. Wong 118 | CUSLSH2885 | CUHK05307 | 22 January 2021 | OM320809 | 4.1 | 221× |
| Hyacinthus orientalis L. ‘Aiolos’ | K. H. Wong 120 | CUSLSH2887 | CUHK05303 | 22 January 2021 | OM320808 | 3.5 | 255× |
| Bellevalia paradoxa (Fisch. et C.A.Mey.) Boiss. | K. H. Wong 126 | CUSLSH2893 | CUHK05320 | 24 January 2021 | OM320811 | 3.8 | 338× |
| Scilla siberica Haw. | K. H. Wong 141 | CUSLSH2917 | CUHK05324 | 11 February 2021 | OM320810 | 3.0 | 231× |
| Hyacinthus orientalis ‘Gipsy Queen’ | Hyacinthus orientalis ‘City of Haarlem’ | Hyacinthus orientalis ‘Eros’ | Hyacinthus orientalis ‘Chicago’ | Hyacinthus orientalis ‘Woodstock’ | Hyacinthus orientalis ‘Delft Blue’ | Hyacinthus orientalis ‘Aiolos’ | Bellevalia paradoxa | Scilla siberica | |
|---|---|---|---|---|---|---|---|---|---|
| Total length (bp) | 154,630 | 154,599 | 154,458 | 154,641 | 154,640 | 154,640 | 154,640 | 154,020 | 154,943 |
| LSC (bp) | 83,180 | 83,149 | 83,168 | 83,159 | 83,159 | 83,159 | 83,159 | 83,028 | 83,384 |
| SSC (bp) | 18,462 | 18,462 | 18,302 | 18,476 | 18,475 | 18,475 | 18,475 | 18,570 | 18,449 |
| IR (bp) | 26,494 | 26,494 | 26,494 | 26,503 | 26,503 | 26,503 | 26,503 | 26,211 | 26,555 |
| Gene no. | 133 | 133 | 133 | 133 | 133 | 133 | 133 | 133 | 133 |
| mRNA | 87 | 87 | 87 | 87 | 87 | 87 | 87 | 87 | 87 |
| tRNA | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 |
| rRNA | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| Pseudogene (Ψ) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 1-intron gene | 21 | 21 | 21 | 21 | 21 | 21 | 21 | 21 | 21 |
| 2-introns gene | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| GC content (%) | 37.58 | 37.58 | 37.60 | 37.58 | 37.58 | 37.58 | 37.58 | 37.66 | 37.60 |
| A content (%) | 30.86 | 30.87 | 30.84 | 30.86 | 30.86 | 30.86 | 30.86 | 30.83 | 30.85 |
| C content (%) | 19.13 | 19.13 | 19.14 | 19.13 | 19.13 | 19.13 | 19.13 | 19.16 | 19.14 |
| G content (%) | 18.45 | 18.45 | 18.46 | 18.45 | 18.45 | 18.45 | 18.45 | 18.50 | 18.46 |
| T content (%) | 31.55 | 31.55 | 31.55 | 31.56 | 31.56 | 31.56 | 31.56 | 31.51 | 31.55 |
| Gene Category | Gene Functions | Gene Names |
|---|---|---|
| Photosynthesis-related genes | Rubisco | rbcL |
| Photosystem I | psaA, psaB, psaC, psaI, psaJ | |
| Assembly/ stability of photosystem I | pafI **, pafII, pbf1 | |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, psbZ | |
| ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | |
| Cytochrome b/f complex | petA, petB *, petD *, petG, petL, petN | |
| Cytochrome c synthesis | ccsA | |
| NADPH dehydrogenase | ndhA *, ndhB * (×2), ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Transcription- and translation-related genes | Transcription | rpoA, rpoB, rpoC1 *, rpoC2 |
| Ribosomal protein | rpl2 * (×2), rpl14, rpl16 *, rpl20, rpl22, rpl23 (×2), rpl32, rpl33, rpl36, rps2, rps3, rps4, rps7 (×2), rps8, rps11, rps12 ** (×2, tran-spliced), rps14, rps15, rps16 *, rps18, rps19 (×2) | |
| Translation initiation factor | infA | |
| RNA genes | Ribosomal RNA | rrn4.5 (×2), rrn5 (×2), rrn16 (×2), rrn23 (×2) |
| Transfer RNA | trnA-UGC * (×2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnG-UCC *, trnH-GUG (×2), trnI-CAU (×2), trnI-GAU * (×2), trnK-UUU *, trnL-CAA (×2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU (×2), trnP-UGG, trnQ-UUG, trnR-ACG (×2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC (×2), trnV-UAC *, trnW-CCA, trnY-GUA | |
| Miscellaneous group | Maturase | matK |
| Inner membrane protein | cemA | |
| ATP-dependent protease | clpP1 ** | |
| Acetyl-CoA carboxylase | accD | |
| Unknown functions | ycf1 (×2), ycf2 (×2) |
| Haplotype | Locus | psbA-trnK-UUU | trnK-UUU | matK-rps16 | rps16 | trnQ-UUG-psbK | psbK-psbI | trnS-GCU-trnG-UCC | trnG-UCC | trnG-UCC-trnR-UCU | trnR-UCU | atpA | atpF | atpF-atpH | atpI-rps2 | rps2-rpoC2 | rpoC2 | rpoC1 | rpoB | ||||||||||||||||||||||||||||
| Position | 1245 | 1282 | 1795 | 2458 | 2664 | 2742 | 3890 | 4395 | 4562 | 4596 | 4689 | 4690 | 4691 | 4939 | 6515 | 7067 | 7739 | 8158 | 8907-8908 | 9531-9533 | 9651 | 10,072 | 10,891 | 11,704 | 12,882 | 12,902 | 14,720 | 15,659 | 16,366 | 17,343 | 19,119 | 19,193 | 21,729 | 23,421 | 23,531 | 23,694 | 24,081 | 25,322 | |||||||||
| 1 | GQ | c | c | g | c | t | g | a | t | c | a | - | - | - | a | c | g | g | a | - t | g g a | t | t | a | a | t | g | - | - | a | t | t | g | t | c | c | g | c | a | ||||||||
| 2 | CH | c | c | g | c | t | g | a | t | c | a | - | - | - | a | c | g | g | a | - t | g g a | t | t | a | a | t | g | - | - | a | t | t | g | t | c | c | g | c | a | ||||||||
| 3 | ER | c | c | g | c | t | g | a | t | c | a | - | - | - | a | c | g | g | a | t t | g g a | t | t | a | a | t | g | - | - | g | t | t | g | t | c | c | g | c | a | ||||||||
| 4 | CO | t | t | a | t | c | a | c | g | t | c | t | t | a | g | t | a | t | g | - - | t c c | c | c | g | g | - | a | a | t | g | c | c | a | c | t | t | t | t | g | ||||||||
| 5 | WS | t | t | a | t | c | a | c | g | t | c | t | t | a | g | t | a | t | g | - - | t c c | c | c | g | g | - | a | a | t | g | c | c | a | c | t | t | t | t | g | ||||||||
| DB | t | t | a | t | c | a | c | g | t | c | t | t | a | g | t | a | t | g | - - | t c c | c | c | g | g | - | a | a | t | g | c | c | a | c | t | t | t | t | g | |||||||||
| AO | t | t | a | t | c | a | c | g | t | c | t | t | a | g | t | a | t | g | - - | t c c | c | c | g | g | - | a | a | t | g | c | c | a | c | t | t | t | t | g | |||||||||
| Haplotype | Gene | rpoB-trnC-GCA | trnC-GCA-petN | petN-psbM | psbM | trnD-GUC-trnY-GUA | trnY-GUA | trnY-GUA-trnE-UUC | trnE-UUC-trnT-GGU | trnT-GGU-psbD | psbC-trnS-UGA | trnS-UGA-psbZ | psbZ-trnG-GCC | psaB | pafI | trnF-GAA-ndhJ | |||||||||||||||||||||||||||||||
| Position | 26,541 | 26,625 | 27,382-27,383 | 27,792 | 28,087 | 28,313 | 28,637 | 28,746 | 29,334-29,339 | 30,065 | 30,688-30,694 | 30,899-30,908 | 30,982 | 31,081 | 31,168 | 31,593 | 31,749 | 35,249 | 35,298-35,304 | 35,516 | 35,596 | 36,142-36,145 | 38,441 | 43,049 | 43,124 | 43,829 | 47,709 | ||||||||||||||||||||
| 1 | GQ | g | - | - - | c | a | t | c | t | - - - - - - | a | a t t g t t a | a a t t g g t t a t | t | c | t | c | t | - | a g t g g g a | g | c | a t a t | c | t | a | g | c | |||||||||||||||||||
| 2 | CH | g | - | - - | c | a | t | c | t | a a a t a a | a | a t t g t t a | a a t t g g t t a t | t | c | t | c | t | - | a g t g g g a | g | c | a t a t | c | t | a | g | t | |||||||||||||||||||
| 3 | ER | g | - | - - | c | a | a | c | t | - - - - - - | a | t a a c a a t | a a t t g g t t a t | c | c | t | c | t | - | a g t g g g a | g | c | a t a t | c | t | a | g | c | |||||||||||||||||||
| 4 | CO | a | t | a g | t | - | t | t | c | - - - - - - | - | t a a c a a t | - - - - - - - - - - | c | t | c | a | c | t | t c c c a c t | a | g | - - - - | t | c | g | a | c | |||||||||||||||||||
| 5 | WS | a | t | a g | t | - | t | t | c | - - - - - - | - | t a a c a a t | - - - - - - - - - - | c | t | c | a | c | t | t c c c a c t | a | g | - - - - | t | c | g | a | c | |||||||||||||||||||
| DB | a | t | a g | t | - | t | t | c | - - - - - - | - | t a a c a a t | - - - - - - - - - - | c | t | c | a | c | t | t t c c c a c t | a | g | - - - - | t | c | g | a | c | ||||||||||||||||||||
| AO | a | t | a g | t | - | t | t | c | - - - - - - | - | t a a c a a t | - - - - - - - - - - | c | t | c | a | c | t | t c c c a c t | a | g | - - - - | t | c | g | a | c | ||||||||||||||||||||
| Haplotype | Locus | trnF-GAA- ndhJ | ndhK | ndhC- trnV-UAC | atpB-rbcL | rbcL-accD | accD | accD-psaI | pafII-cemA | petA | petA-psbJ | psbE-petL | |||||||||||||||||||||||||||||||||||
| Position | 47,711-47,747 | 47,788-47,789 | 47,962 | 49,041 | 50,274 | 50,292 | 50,366 | 54,305-54,306 | 54,337 | 56,269 | 56,271 | 56,273 | 56,703-56,708 | 56,787 | 57,943 | 58,550 | 60,521 | 62,324 | 62,723 | 62,868-62,869 | 62,939 | 64,219-64,223 | |||||||||||||||||||||||||
| 1 | GQ | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | - - | g | g | t | a | g | - - | c | c | a | a | t a t t t a | t | t | a | - | t | c | t c | t | t t t g t | ||||||||||||||||||||||||
| 2 | CH | - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - | - - | g | g | t | a | g | - - | c | c | a | a | t a t t t a | t | t | a | - | t | c | t c | t | t t t g t | ||||||||||||||||||||||||
| 3 | ER | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | - - | g | g | - | t | g | - - | c | t | t | g | t a t t t a | c | t | c | t | t | c | g a | t | t t t g t | ||||||||||||||||||||||||
| 4 | CO | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | a t | a | a | - | a | t | t a | a | t | t | g | - - - - - - | c | c | c | t | c | - | t c | c | - - - - - | ||||||||||||||||||||||||
| 5 | WS | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | a t | a | a | - | a | t | t a | a | t | t | g | - - - - - | c | c | c | t | c | - | t c | c | - - - - - | ||||||||||||||||||||||||
| DB | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | a t | a | a | - | a | t | t a | a | t | t | g | - - - - - - | c | c | c | t | c | - | t c | c | - - - - - | |||||||||||||||||||||||||
| AO | t t t c t g g t t t t t t c a t a c t g t g c t t t c t c t c a c t c a a | a t | a | a | - | a | t | t a | a | t | t | g | - - - - - - | c | c | c | t | c | - | t c | c | - - - - - | |||||||||||||||||||||||||
| Haplotype | Locus | psbE-petL | rpl33-rps18 | rps18-rpl20 | rpl20-rps12 | clpP1 | clpP1-psbB | psbB-psbT | petB | petD | petD-rpoA | rpoA | rps11 | rpl36 | rps8-rpl14 | rpl14-rpl16 | rpl16 | rpl22-rps19 | |||||||||||||||||||||||||||||
| Position | 64734 | 64962 | 65097 | 66897 | 67194-67199 | 67809 | 68912 | 69694 | 69883 | 69975 | 70147 | 71437 | 73343-73344 | 73373 | 74314 | 76275 | 77090 | 77746 | 78243 | 78556 | 78967 | 79305-79306 | 79930 | 80483 | 80905 | 81004-81008 | 81468 | 81662 | 81738 | 81847 | 83198-83212 | ||||||||||||||||
| 1 | GQ | g | c | g | - | a t g t g t | a | - | a | t | t | - | - | - t | a | a | t | g | g | a | c | g | c a | t | g | g | - - - - - | c | t | - | c | g a t t t c t t t a t c a t a | |||||||||||||||
| 2 | CH | g | c | g | - | a t g t g t | a | - | a | t | t | - | - | - t | a | a | t | g | g | a | c | g | c a | t | g | g | - - - - - | c | t | - | c | g a t t t c t t t a t c a t a | |||||||||||||||
| 3 | ER | g | c | g | - | a t g t g t | a | - | a | t | t | t | t | t t | a | g | t | g | g | a | c | g | c a | - | g | g | - - - - - | c | c | - | c | - - - - - - - - - - - - - - - | |||||||||||||||
| 4 | CO | a | a | t | t | - - - - - - | - | t | - | - | c | - | - | - - | t | a | g | a | a | g | t | a | - - | t | a | a | t t a a t | t | t | a | t | g a t t t c t t t a t c a t a | |||||||||||||||
| 5 | WS | a | a | t | t | - - - - - - | - | t | - | - | c | - | - | - - | t | a | g | a | a | g | t | a | - - | t | a | a | t t a a t | t | t | a | t | g a t t t c t t t a t c a t a | |||||||||||||||
| DB | a | a | t | t | - - - - - - | - | t | - | - | c | - | - | - - | t | a | g | a | a | g | t | a | - - | t | a | a | t t a a t | t | t | a | t | g a t t t c t t t a t c a t a | ||||||||||||||||
| AO | a | a | t | t | - - - - - - | - | t | - | - | c | - | - | - - | t | a | g | a | a | g | t | a | - - | t | a | a | t t a a t | t | t | a | t | g a t t t c t t t a t c a t a | ||||||||||||||||
| Haplotype | Locus | rps19 | ycf2 | ndhB-rps7 | trnR-ACG-trnN-GUU | ycf1 | ndhF | ndhF-rpl32 | |||||||||||||||||||||||||||||||||||||||
| Position | 83248 | 86579 | 90294 | 96775 | 107482- 107490 | 107915 | 107924 | 107931 | 109716 | 110054 | 110285 | 112295-112398 | |||||||||||||||||||||||||||||||||||
| 1 | GQ | t | t | t | t | - - - - - - - - - | - | t | g | - | c | a | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | ||||||||||||||||||||||||||||||||||
| 2 | CH | t | t | t | t | - - - - - - - - - | - | t | g | - | c | a | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | ||||||||||||||||||||||||||||||||||
| 3 | ER | t | t | t | t | - - - - - - - - - | - | t | g | - | c | a | - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - | ||||||||||||||||||||||||||||||||||
| 4 | CO | - | c | c | c | c a g a a a a a a g | c | c | t | t | t | g | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | ||||||||||||||||||||||||||||||||||
| 5 | WS | - | c | c | c | a g a a a a a a g | c | c | t | - | t | g | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | ||||||||||||||||||||||||||||||||||
| DB | - | c | c | c | a g a a a a a a g | c | c | t | - | t | g | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | |||||||||||||||||||||||||||||||||||
| AO | - | c | c | c | a g a a a a a a g | c | c | t | - | t | g | t a a a a a a g t a t a g t a a g t a a a a a c a a t t a t a t c a a a a c a g t a g a a t a t g g a t c a t a a t a t g c a t t g a t a a a t c g a c t a a a a a a a a a a g a c c t a a t t a t t c t a a t | |||||||||||||||||||||||||||||||||||
| Haplotype | Locus | ndhF-rpl32 | ndhF-rpl32 | rpl32- trnL-UAG | ccsA-ndhD | ndhD | psaC-ndhE | ndhE-ndhG | ndhI | ndhI-ndhA | ndhA | ndhH | rps15 | rps15-ycf1 | |||||||||||||||||||||||||||||||||
| Position | 112399-112455 | 112621 | 113173 | 113555 | 115044-115048 | 115050 | 115345 | 117286-117290 | 117432-117438 | 117922 | 119067 | 119459 | 120327 | 120328 | 120555 | 120815 | 120971 | 122856 | 123081 | 123607 | 123669 | 123670 | |||||||||||||||||||||||||
| 1 | GQ | g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | c | - | a | a g g a a | c | g | - - - - - | - - - - - - - | t | g | g | - | - | - | a | c | a | g | c | c | a | ||||||||||||||||||||||||
| 2 | CH | g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | c | - | a | a g g a a | c | g | - - - - - | - - - - - - - | t | g | g | - | - | - | a | c | a | g | c | c | a | ||||||||||||||||||||||||
| 3 | ER | - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - | c | - | a | t t c c t | c | g | - - - - - | - - - - - - - | t | g | g | - | - | a | a | c | a | g | c | c | a | ||||||||||||||||||||||||
| 4 | CO | g g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | t | t | g | t t c c t | g | t | a t g t g | a t t t a t a | a | a | t | a | a | - | g | t | g | t | g | - | - | ||||||||||||||||||||||||
| 5 | WS | g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | t | t | g | t t c c t | g | t | a t g t g | a t t t a t a | a | a | t | a | a | - | g | t | g | t | g | - | - | ||||||||||||||||||||||||
| DB | g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | t | t | g | t t c c t | g | t | a t g t g | a t t t a t a | a | a | t | a | a | - | g | t | g | t | g | - | - | |||||||||||||||||||||||||
| AO | g a a t t c a t t t g t a g t a a t t a c c t a a t t c a t t g c a g a a t t t a t t g g a t t c c a a t c c a a | t | t | g | t t c c t | g | t | a t g t g | a t t t a t a | a | a | t | a | a | - | g | t | g | t | g | - | - | |||||||||||||||||||||||||
| Haplotype | Locus | ycf1 | trnN-GUU- trnR-ACG | trnR-ACG- rrn5 | rps7-ndhB | ycf2 | rps19-psbA | ||||||||||||||||||||||||||||||||||||||||
| Position | 125016 | 125184 | 125908 | 126456 | 127892 | 129981 | 129988- 129989 | 130415- 130423 | 141137 | 147618 | 151333 | 154655 | |||||||||||||||||||||||||||||||||||
| 1 | GQ | a | c | g | c | a | c | - a | - - - - - - - - - | a | a | a | a | ||||||||||||||||||||||||||||||||||
| 2 | CH | a | c | g | c | a | c | - a | - - - - - - - - - | a | a | a | a | ||||||||||||||||||||||||||||||||||
| 3 | ER | a | c | g | c | a | c | - a | - - - - - - - - - | a | a | a | a | ||||||||||||||||||||||||||||||||||
| 4 | CO | g | t | a | a | c | a | g g | t t t t t c t c t | g | g | g | - | ||||||||||||||||||||||||||||||||||
| 5 | WS | g | t | a | a | c | a | g g | t t t t t c t c t | g | g | g | - | ||||||||||||||||||||||||||||||||||
| DB | g | t | a | a | c | a | g g | t t t t t c t c t | g | g | g | - | |||||||||||||||||||||||||||||||||||
| AO | g | t | a | a | c | a | g g | t t t t t c t c t | g | g | g | - | |||||||||||||||||||||||||||||||||||
| Cultivar | Date of Registration | Registrant | Description | Perianth | Forcing Time | Reference |
|---|---|---|---|---|---|---|
| ‘City of Haarlem’ | 1893/12/31 | J.H. Kersten | Soft primrose-yellow | Single | Late forcing | [19,39] |
| ‘Gipsy Queen’ | 1927/dd/mm * | G. van der Meij | Dark salmon (421/1) and apricot (608/1) | Single | Mid early forcing | [19,39] |
| ‘Eros’ | 1935/12/31 | M. Veldhuyzen-van Zanten | Deep pink | Double | Early forcing | [19,39] |
| ‘Delft Blue’ | 1944/12/31 | J.W.A. Lefeber | Soft lilac-blue | Single | Early forcing | [19,39] |
| ‘Aiolos’ | 1985/12/31 | I.V.T. | Ivory white, outside tube creamy white, grey-yellow blotch at top | Single | / | [19,39] |
| ‘Woodstock’ | 1992/9/17 | Jac. Prins et Zn. B.V. | / | Single | / | [39] |
| ‘Chicago’ | 2009/6/17 | Fa. M.C. Zonneveld et Zn. | / | Single | / | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, K.-H.; Wu, H.-Y.; Kong, B.L.-H.; But, G.W.-C.; Siu, T.-Y.; Hui, J.H.-L.; Shaw, P.-C.; Lau, D.T.-W. Characterisation of the Complete Chloroplast Genomes of Seven Hyacinthus orientalis L. Cultivars: Insights into Cultivar Phylogeny. Horticulturae 2022, 8, 453. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050453
Wong K-H, Wu H-Y, Kong BL-H, But GW-C, Siu T-Y, Hui JH-L, Shaw P-C, Lau DT-W. Characterisation of the Complete Chloroplast Genomes of Seven Hyacinthus orientalis L. Cultivars: Insights into Cultivar Phylogeny. Horticulturae. 2022; 8(5):453. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050453
Chicago/Turabian StyleWong, Kwan-Ho, Hoi-Yan Wu, Bobby Lim-Ho Kong, Grace Wing-Chiu But, Tin-Yan Siu, Jerome Ho-Lam Hui, Pang-Chui Shaw, and David Tai-Wai Lau. 2022. "Characterisation of the Complete Chloroplast Genomes of Seven Hyacinthus orientalis L. Cultivars: Insights into Cultivar Phylogeny" Horticulturae 8, no. 5: 453. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050453