Downregulation of the Expression of GLUT1 Plays a Role in Apoptosis Induced by Sodium Butyrate in HT-29 Cell Line

Abstract
:1. Introduction
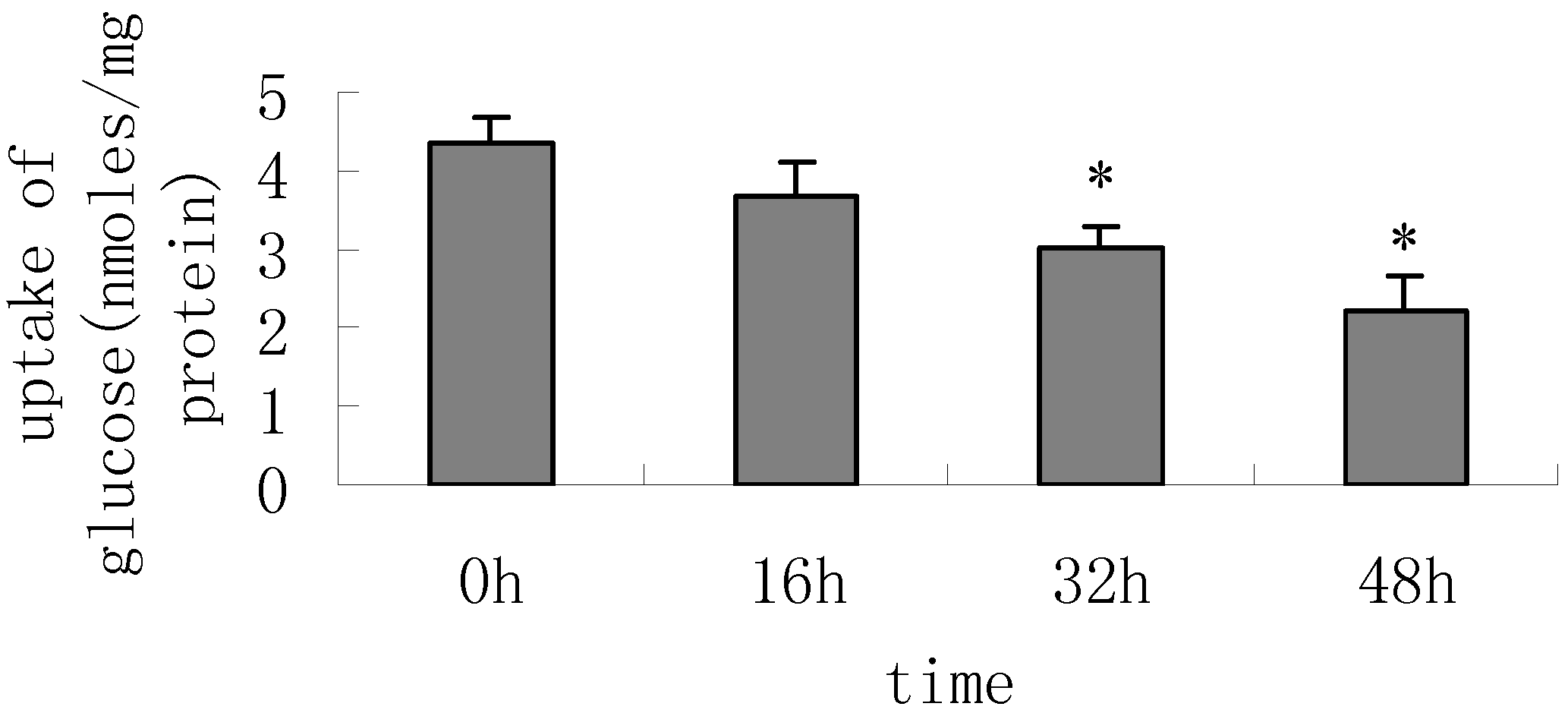
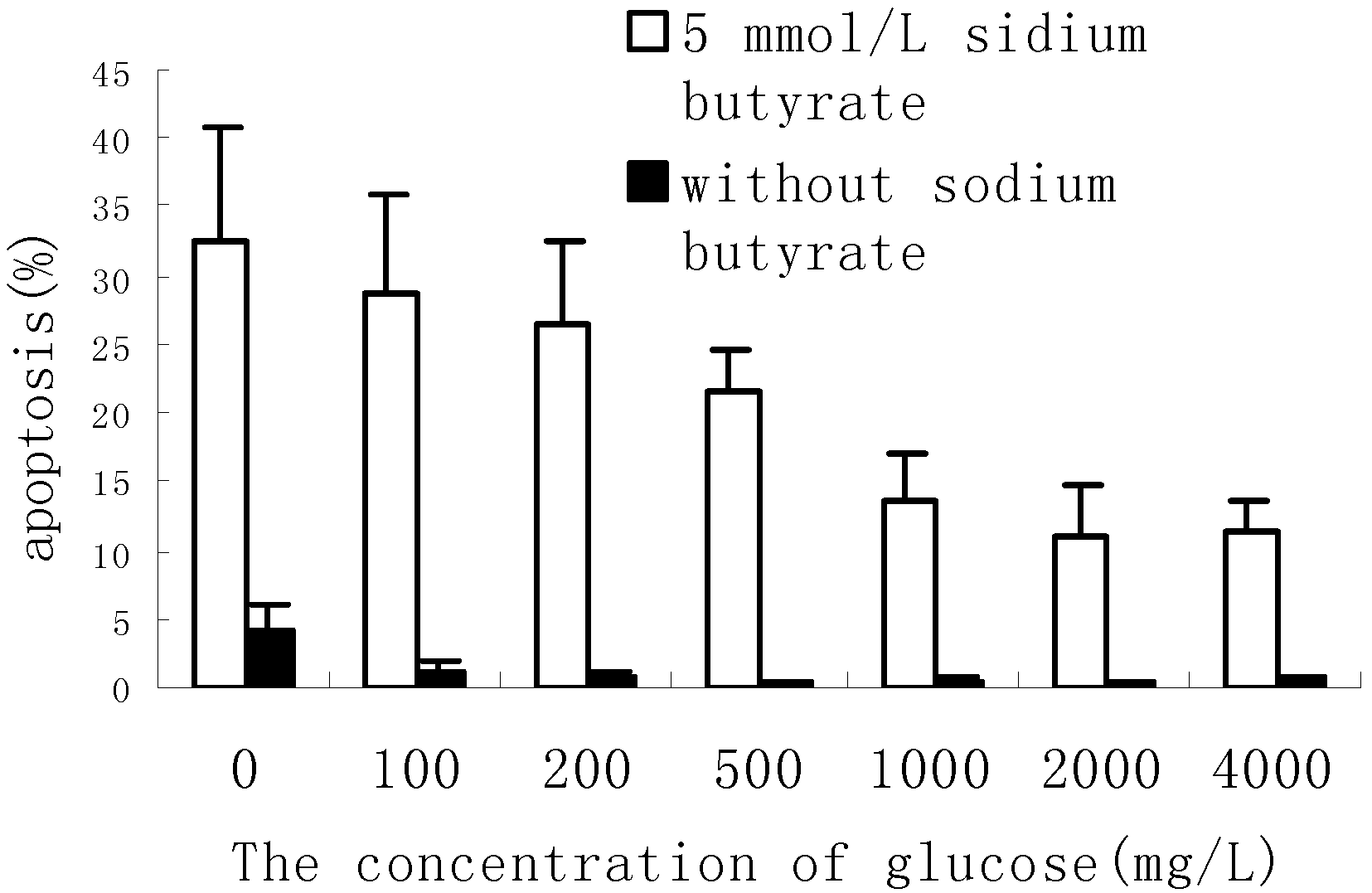
Result and discussion





Conclusion
Experimental
Cells and Cell Culture
RNA Preparation
Reverse Transcriptase-polymerase Chain Reaction
Western Blot

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers (sense and anti-sense) | Size of PCR product (bp) | |
|---|---|---|
| GLUT1 | 5’ CGGGCCAAGAGTGTGCTAAA 3’ | 283bp |
| 5’ TGACGATACCGGAGCCAATG 3’ | ||
| GLUT2 | 5’CGTCTCCTTTGACATTTCCTTC3’ | 221bp |
| 5’GGTGGAGAAAACAGCCTAGAGAT3’ | ||
| GLUT3 | 5’CCAACTTCCTAGTCGGATTG3’ | 250bp |
| 5’AGGAGGCACGACTTAGACAT3’ | ||
| GLUT4 | 5’CCCCCTCAGCAGCGAGTGA3’ | 319bp |
| 5’GCACCGCCAGCACATTGTTG3’ | ||
| GLUT5 | 5’GCAACAGGATCAGAGCATGA3’ | 316bp |
| 5’TCGCAGGCACGATAGAAAAT3’ | ||
| MCT1 | 5’ CACCACCAGCGAAGTGTC 3’ | 158bp |
| 5’ AGAAAGAAGCTGCAATCAAG 3’ | ||
| β-actin | 5’ CGAGCGGGAAATCGTGCGTGACATTAAGGAGA 3’ | 479bp |
| 5’ CGTCATACTCCTGCTTGCTGATCCACATCTGC 3’ | ||
The Uptake of Glucose Assay
Apoptosis Assay
Acknowledgments
References and Notes
- Jacobs, L.R. Fiber and colon cancer. Gastroenterol Clin. North. Am. 1988, 17, 747–60. [Google Scholar]
- Hernandez, A.; Thomas, R.; Smith, F.; Sandberg, J.; Kim, S.; Chung, DH.; Evers, B.M. Butyrate sensitizes human colon cancer cells to TRAIL-mediated apoptosis. Surgery 2001, 130, 265–72. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, X.; Hernandez, A.; Kim, S.; Evers, B.M. Inhibition of the phosphatidylinositol 3-kinase pathway contributes to HT29 andCaco-2 intestinal cell differentiation. Gastroenterology 2001, 120, 1381–1392. [Google Scholar]
- Otaka, M.; Singhal, A.; Hakomori, S. Antibody-mediated targeting of differentiation inducers to tumor cells:inhibition of colonic cancer cell growth in vitro and in vivo. Biochem.Biophys. Res. Commun. 1989, 158, 202–208. [Google Scholar] [CrossRef]
- Vidyasagar, S.; Ramakrishna, B.S. Effects of butyrate on active sodium and chloride transport in rat and rabbitdistal colon. J. Physiol. 2002, 539, 163–173. [Google Scholar] [CrossRef]
- Scheppach, W.; Bartram, P.; Richter, A.; Richter, F.; Liepold, H.; Dusel, G.; Hofstetter, G.; Ruthlein, J.; Kasper, H. Effect of short-chain fatty acids on the human colonic mucosa in vitro. JPEN. J. Parenter. Entera. l. Nutr. 1992, 16, 43–48. [Google Scholar] [CrossRef]
- Butzner, J.D.; Meddings, J.B.; Dalal, V. Inhibition of short-chain fatty acid absorption and Na+ absorption during acute colitis in the rabbit. Gastroenterology 1994, 106, 1190–1198. [Google Scholar]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef]
- Stein, J.; Zores, M.; Schroder, O. Short-chain fatty acid (SCFA) uptake into Caco-2 cells by a pH-dependent and carrier mediated transport mechanism. Eur. J. Nutr. 2000, 39, 121–125. [Google Scholar] [CrossRef]
- Younes, M.; Lechago, L.V.; Somoano, J.R.; Mosharaf, M.; Lechago, J. Wide expression of the human erythrocyte glucose transporter Glut1 in human cancers. Cancer Res. 1996, 56, 1164–1167. [Google Scholar]
- Bozzetti, F.; Gavazzi, C.; Mariani, L.; Crippa, F. Glucose-based total parenteral nutrition does not stimulate glucose uptake by humans tumours. Clin. Nutr. 2004, 23, 417–421. [Google Scholar] [CrossRef]
- Noguchi, Y.; Marat, D.; Saito, A.; Yoshikawa, T.; Doi, C.; Fukuzawa, K.; Tsuburaya, A.; Satoh, S.; Ito, T. Expression of facilitative glucose transporters in gastric tumors. Hepatogastroenterology. 1999, 46, 2683–2689. [Google Scholar]
- Mellanen, P.; Minn, H.; Grenman, R.; Harkonen, P. Expression of glucose transporters in head-and-neck tumors. Int. J. Cancer. 1994, 6, 622–629. [Google Scholar]
- Younes, M.; Brown, R.W.; Stephenson, M.; Gondo, M.; Cagle, P.T. Overexpression of Glut1 and Glut3 in stage I nonsmall cell lung carcinoma is associated with poor survival. Cancer 1997, 80, 1046–1051. [Google Scholar] [CrossRef]
- Brown, R.S.; Wahl, R.L. Overexpression of Glut-1 glucose transporter in human breast cancer. Cancer 1993, 72, 2979–2985. [Google Scholar] [CrossRef]
- Rivenzon-Segal, D.; Rushkin, E.; Polak-Charcon, S.; Degani, H. Glucose transporters and transport kinetics in retinoic acid-differentiated T47D human breast cancer cells. Am. J. Physiol. Endocrinol. Metab. 2000, 279, 508–519. [Google Scholar]
- Kang, S.S.; Chun, Y.K.; Hur, M.H.; Lee, H.K.; Kim, Y.J.; Hong, S.R.; Lee, J.H.; Lee, S.G.; Park, Y.K. Clinical significance of glucose transporter 1 (GLUT1) expression in human breast carcinoma . Jpn. J. Cancer Res. 2002, 93, 1123–1128. [Google Scholar] [CrossRef]
- Lambert, D.W.; Wood, I.S.; Ellis, A.; Shirazi-Beechey, S.P. Molecular changes in the expression of human colonic nutrient transporters during the transition from normality to malignancy. Br. J. Cancer 2002, 86, 1262–1269. [Google Scholar] [CrossRef]
- Hadjiagapiou, C.; Schmidt, L.; Dudeja, P.K.; Layden, T.J. Mechanism(s) of butyrate transport in Caco-2 cells: role of monocarboxylatetransporter 1. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, 775–780. [Google Scholar]
- Ritzhaupt, A.; Wood, I.S.; Ellis, A.; Hosie, K.B.; Shirazi-Beechey, S.P. Identification and characterization of a monocarboxylate transporter (MCT1) inpig and human colon: its potential to transport L-lactate as well as butyrate. J. Physiol. 1998, 513, 719–732. [Google Scholar] [CrossRef]
- Cuff, M.A.; Lambert, D.W.; Shirazi-Beechey, S.P. Substrate-induced regulation of the human colonic monocarboxylate transporter,MCT1. J. Physiol. 2002, 539, 361–371. [Google Scholar] [CrossRef]
- Pellizzaro, C.; Coradini, D.; Daidone, M.G. Modulation of angiogenesis-related proteins synthesis by sodium butyrate in colon cancer cell line HT29. Carcinogenesis 2002, 23, 735–740. [Google Scholar] [CrossRef]
- Choi, J.W.; Yoon, D.J.; Lee, H.W.; Han, D.P.; Ahn, Y.H. Antisense GLUT1 RNA suppresses the transforming phenotypes of NIH 3T3 cells transformed by N-Ras. Yonsei Med. J. 1995, 36, 480–486. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Noguchi, Y.; Okamoto, T.; Marat, D.; Yoshikawa, T.; Saitoh, A.; Doi, C.; Fukuzawa, K.; Tsuburaya, A.; Satoh, S.; Ito, T. Expression of facilitative glucose transporter 1 mRNA in colon cancer was not regulated by k-ras. Cancer Lett. 2000, 154, 137–142. [Google Scholar] [CrossRef]
- Younes, M.; Lechago, L.V. Lechago, Overexpression of the human erythrocyte glucose transporter occurs as a late event in human colorectal carcinogenesis and is associated with an increased incidence of lymph node metastases. J. Clin.Cancer Res. 1996, 2, 1151–1154. [Google Scholar]
- Loberg, R.D.; Vesely, E.; Brosius, F.C. Enhanced glycogen synthase kinase-3beta activity mediates hypoxia-induced apoptosis of vascular smooth muscle cells and is prevented by glucose transport and metabolism. J. Biol. Chem. 2002, 277, 41667–41673. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Plas, D.R.; Rathmell, J.C.; Fox, C.J.; Harris, M.H.; Thompson, C.B. Growth factors can influence cell growth and survival through effects on glucose metabolism. Mol. Cell. Biol. 2001, 21, 5899–5912. [Google Scholar] [CrossRef]
- Moley, K.H.; Mueckler, M.M. Glucose transport and apoptosis. Apoptosis 2000, 5, 99–105. [Google Scholar] [CrossRef]
- Takata, K.; Hirano, H. Mechanism of glucose transport across the human and rat placental barrier. Microsc. Res. Tech. 1997, 38, 145–152. [Google Scholar] [CrossRef]
- Sato, Y.; Ito, T.; Udaka, N.; Kanisawa, M.; Noguchi, Y.; Cushman, S.W.; Satoh, S. Immunohistochemical localization of facilitated-diffusion glucose transportersin rat pancreatic islets. Tissue Cell 1996, 28, 637–643. [Google Scholar] [CrossRef]
- Kovarikova, M.; Hofmanova, J.; Soucek, K.; Kozubik, A. The effects of TNF-alpha and inhibitors of arachidonic acid metabolism on human colon HT-29 cells depend on differentiation status. Differentiation 2004, 72, 23–31. [Google Scholar] [CrossRef]
- Ruemmele, F.M.; Schwartz, S.; Seidman, E.G.; Dionne, S.; Levy, E.; Lentze, M. J. Butyrate induced Caco-2 cell apoptosis is mediated via the mitochondrial pathway. Gut. 2003, 52, 94–100. [Google Scholar] [CrossRef]
- Munoz-Pinedo, C.; Ruiz-Ruiz, C.; Ruiz de Almodovar, C.; Palacios, C.; Lopez-Rivas, A. Inhibition of glucose metabolism sensitizes tumor cells to death receptor-triggered apoptosis through enhancement of death-inducing signalingcomplex formation and apical procaspase-8 processing. J. Biol. Chem. 2003, 278, 12759–12768. [Google Scholar] [CrossRef]
- Wacheck, V.; Selzer, E.; Gunsberg, P.; Lucas, T.; Meyer, H.; Thallinger, C.; Monia, B.P.; Jansen, B. Bcl-x(L) antisense oligonucleotidesradiosensitise colon cancer cells. Br. J. Cancer 2003, 89, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.Y.; Li, D.C.; Zhang, Z.D.; Zhao, J.; Ge, J.F. Adenovirus-mediated FasL gene transfer into human gastric carcinoma. World. J. Gastroenterol. 2005, 11, 3446–3450. [Google Scholar]
© 2006 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Li, X.; Luo, H.-S.; Paul, S.C.; Tang, T.; Yuan, G.-J. Downregulation of the Expression of GLUT1 Plays a Role in Apoptosis Induced by Sodium Butyrate in HT-29 Cell Line. Int. J. Mol. Sci. 2006, 7, 59-70. https://0-doi-org.brum.beds.ac.uk/10.3390/i7020059
Li X, Luo H-S, Paul SC, Tang T, Yuan G-J. Downregulation of the Expression of GLUT1 Plays a Role in Apoptosis Induced by Sodium Butyrate in HT-29 Cell Line. International Journal of Molecular Sciences. 2006; 7(2):59-70. https://0-doi-org.brum.beds.ac.uk/10.3390/i7020059
Chicago/Turabian StyleLi, Xi, He-Sheng Luo, Shelley C. Paul, Tao Tang, and Guang-Jin Yuan. 2006. "Downregulation of the Expression of GLUT1 Plays a Role in Apoptosis Induced by Sodium Butyrate in HT-29 Cell Line" International Journal of Molecular Sciences 7, no. 2: 59-70. https://0-doi-org.brum.beds.ac.uk/10.3390/i7020059