Optimization of Pulse-Field Gel Electrophoresis for Subtyping of Klebsiella pneumoniae

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. PFGE Protocol
2.3. Computer Analysis of PFGE Patterns
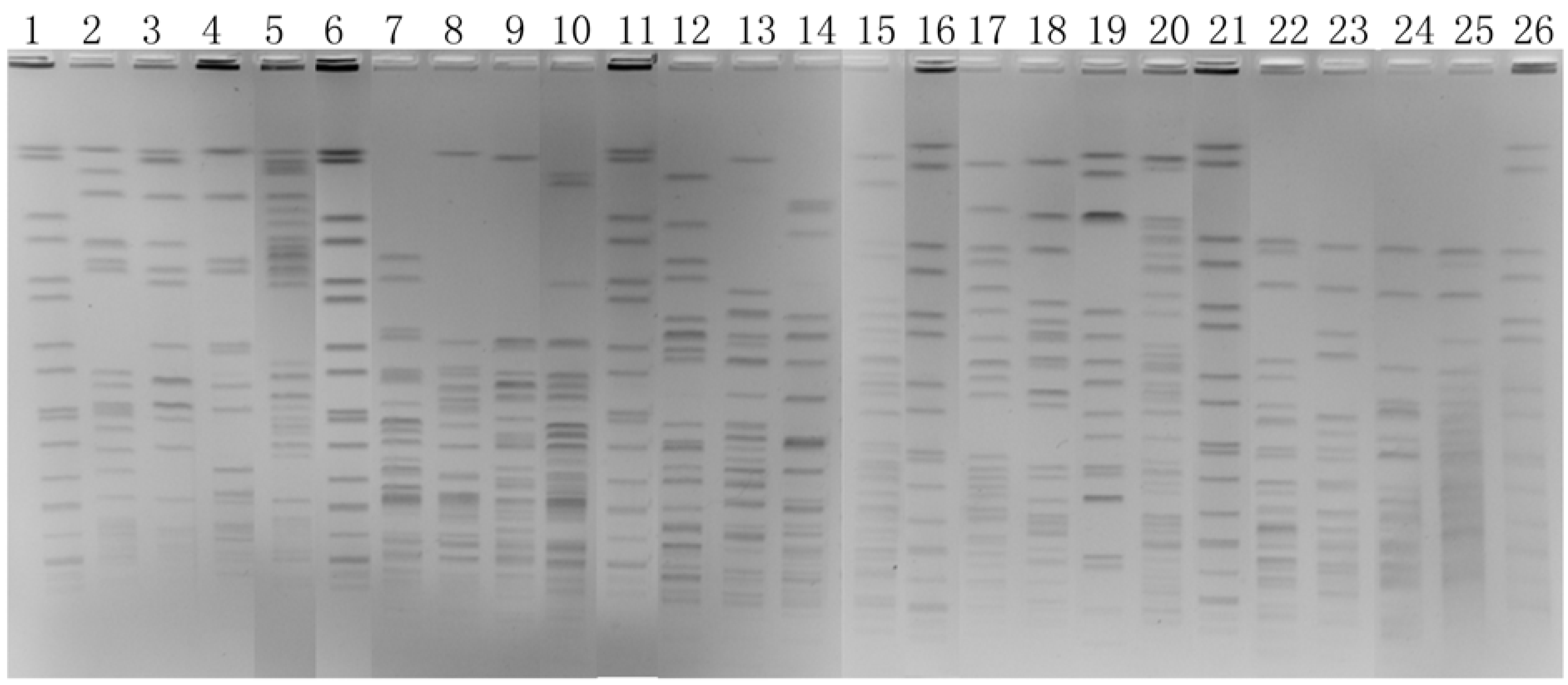
2.4. Enzyme Selection
2.5. Optimization of Electrophoresis Parameters (EP) for Optimal Enzyme Digestion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrophoresis parameters | Switch time (s) | Total run time (h) |
|---|---|---|
| EP-a | 6–20 | 18 |
| EP-b | 6–36 | 18.5 |
| EP-c | 4–40 | 19 |
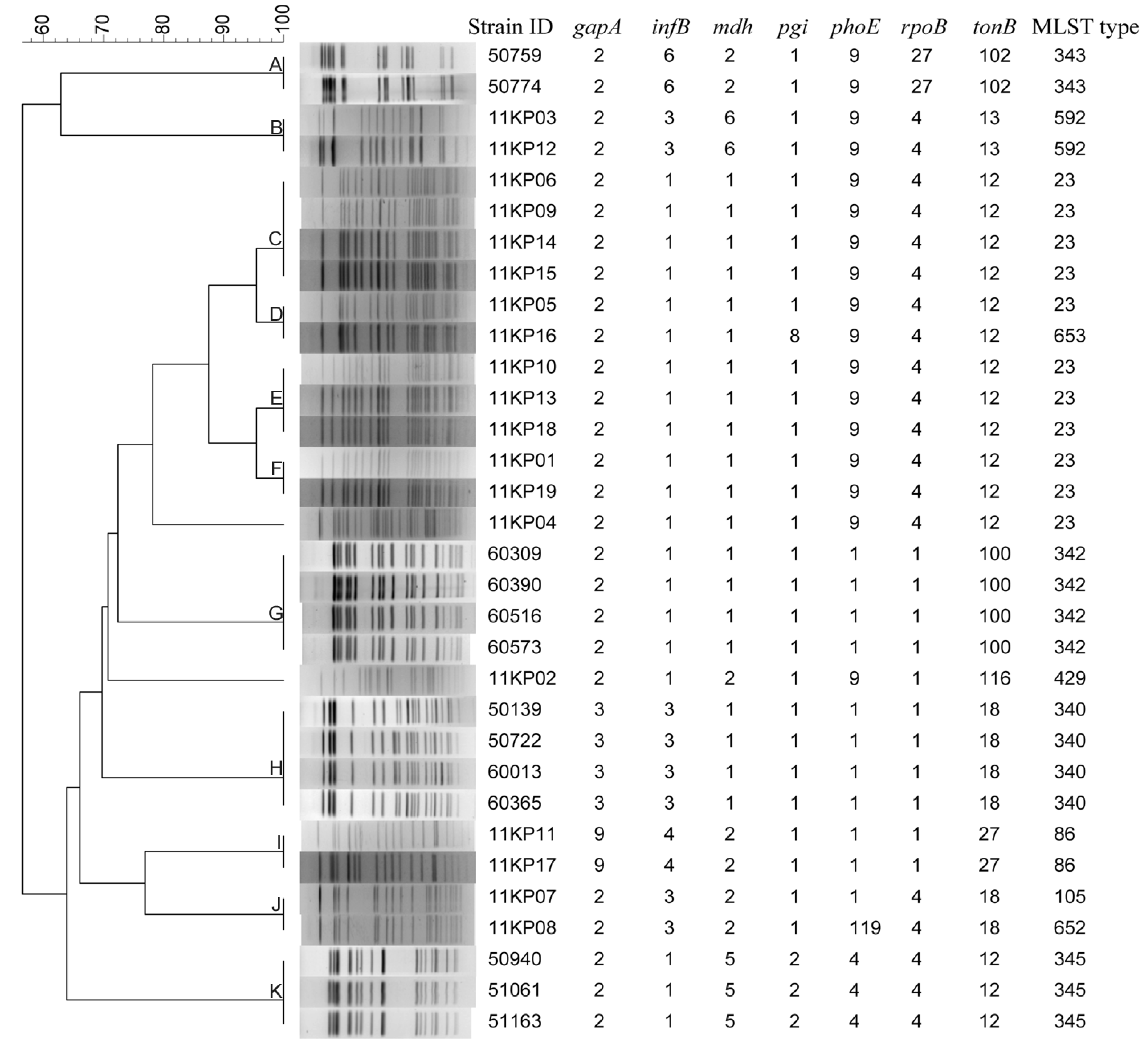
2.6. Multilocus Sequence Typing
3. Results
3.1. Optimization of Electrophoretic Parameters

3.2. Selection of Another Enzyme

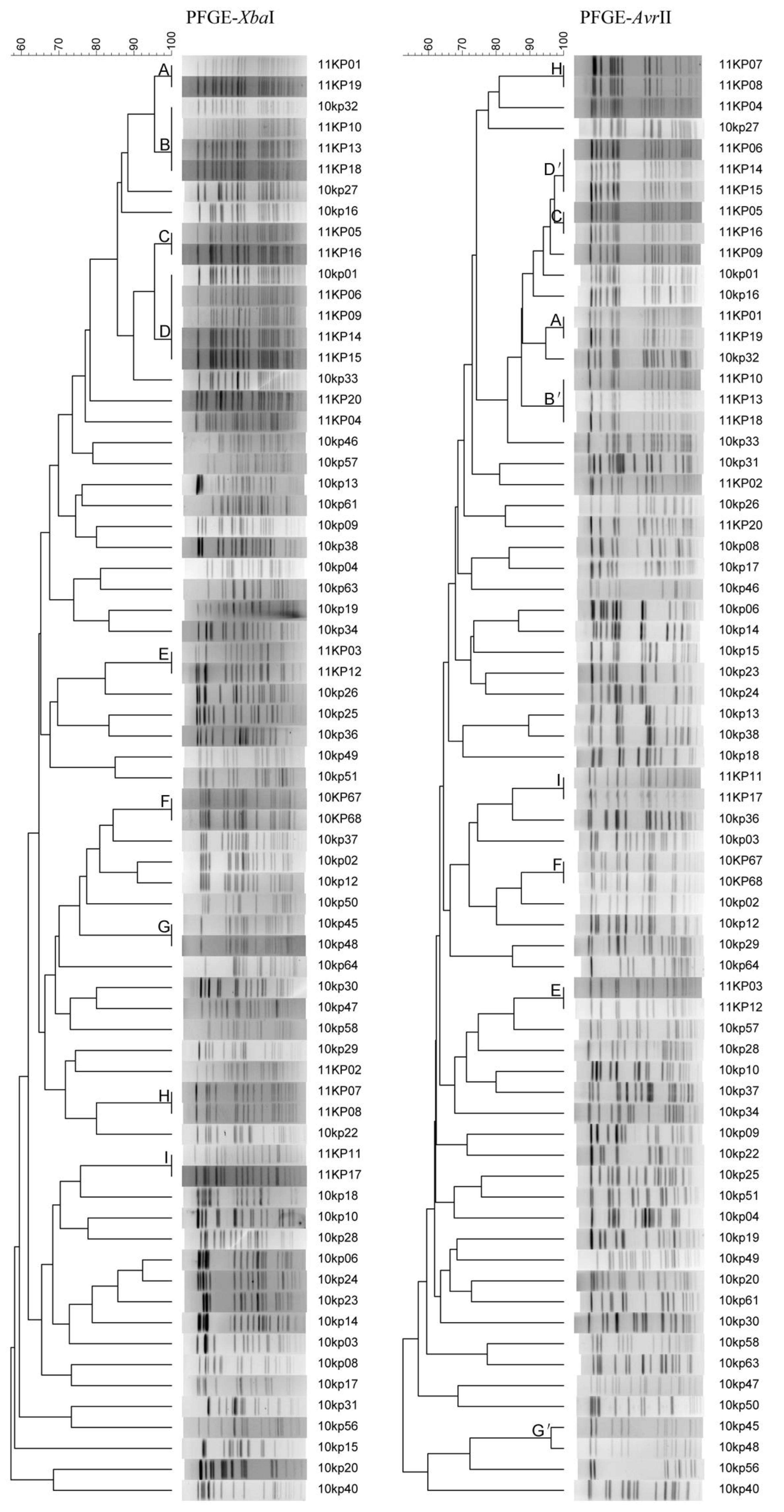
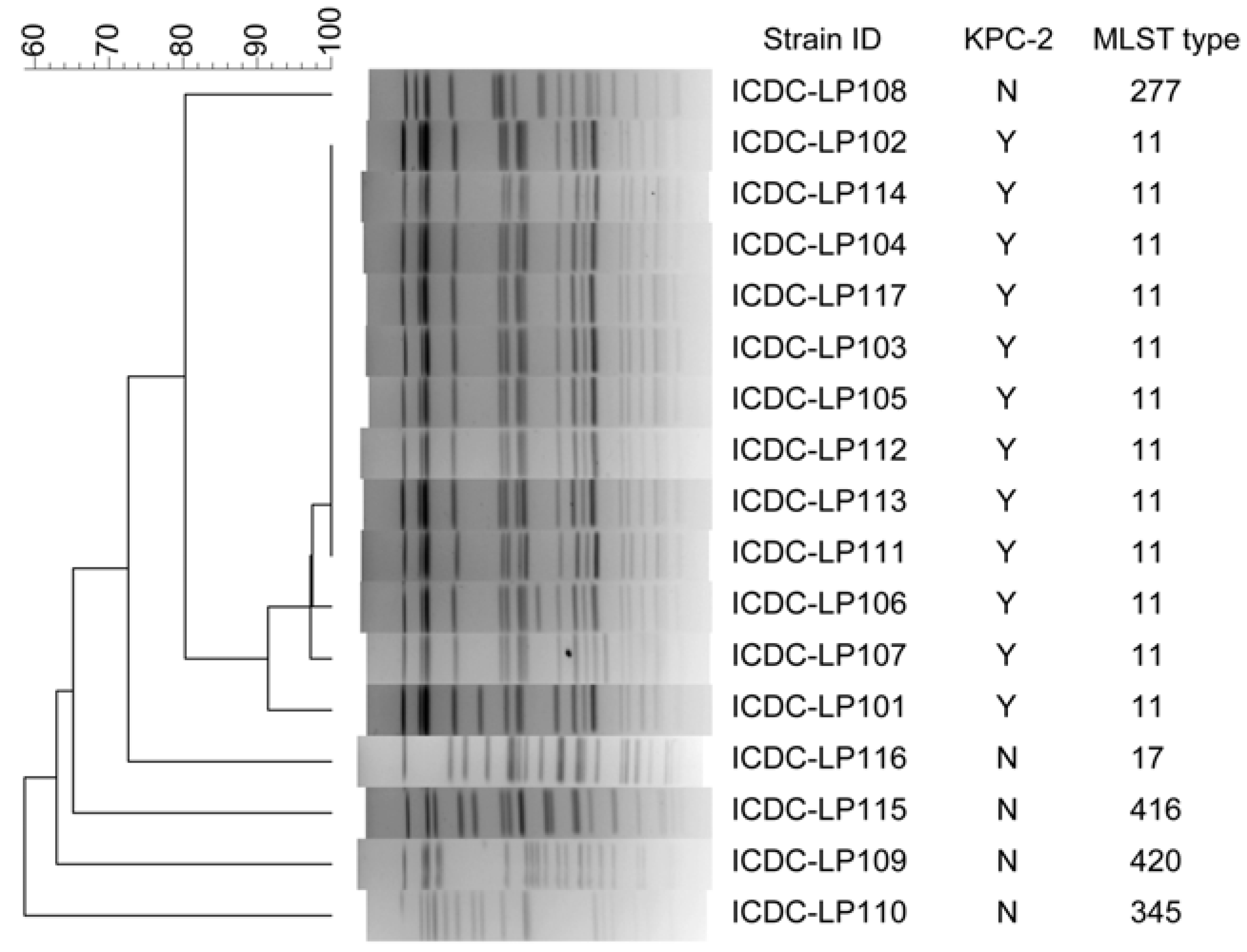
3.3. Typeability, Reproducibility and Discriminatory Power


3.4. Concordance between PFGE and MLST Methods


4. Discussion and Conclusions
Acknowledgments
Conflict of Interest
References
- Fung, C.P.; Chang, F.Y.; Lee, S.C.; Hu, B.S.; Kuo, B.I.; Liu, C.Y.; Ho, M.; Siu, L.K. A global emerging disease of Klebsiella pneumoniae liver abscess: Is serotype K1 an important factor for complicated endophthalmitis? Gut 2002, 50, 420–424. [Google Scholar] [CrossRef]
- Ko, W.C.; Paterson, D.L.; Sagnimeni, A.J.; Hansen, D.S.; Von Gottberg, A.; Mohapatra, S.; Casellas, J.M.; Goossens, H.; Mulazimoglu, L.; Trenholme, G.; et al. Community-acquired Klebsiella pneumoniae bacteremia: Global differences in clinical patterns. Emerg. Infect. Dis. 2002, 8, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Karama, E.M.; Willermain, F.; Janssens, X.; Claus, M.; Van den Wijngaert, S.; Wang, J.T.; Verougstraete, C.; Caspers, L. Endogenous endophthalmitis complicating Klebsiella pneumoniae liver abscess in Europe: Case report. Int. Ophthalmol. 2008, 28, 111–113. [Google Scholar] [CrossRef]
- Cuzon, G.; Naas, T.; Truong, H.; Villegas, M.; Wisell, K.T.; Carmeli, Y.; Gales, A.C.; Navon-Venezia, S.; Quinn, J.P.; Nordmann, P. Worldwide diversity of Klebsiella pneumoniae that produce β-Lactamase blaKPC-2 gene. Emerg. Infect. Dis. 2010, 16, 1349–1356. [Google Scholar]
- Nordmann, P.; Naas, T.; Poirel, L. Global Spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef]
- Bratu, S.; Landman, D.; Haag, R.; Recco, R.; Eramo, A.; Alam, M.; Quale, J. Rapid spread of carbapenem-resistant Klebsiella pneumoniae in New York City: A new threat to our antibiotic armamentarium. Arch Intern. Med. 2005, 165, 1430–1435. [Google Scholar] [CrossRef]
- Jonas, D.; Spitzmüller, B.; Daschner, F.D.; Verhoef, J.; Brisse, S. Discrimination of Klebsiella pneumoniae and Klebsiella oxytoca phylogenetic groups and other Klebsiella species by use of amplified fragment length polymorphism. Res. Microbiol. 2004, 155, 17–23. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef]
- Lau, Y.J.; Hu, B.S.; Wu, W.L.; Lin, Y.H.; Chang, H.Y.; Shi, Z.Y. Identification of a major cluster of Klebsiella pneumoniae isolates from patients with liver abscess in Taiwan. J. Clin. Microbiol. 2000, 38, 412–414. [Google Scholar]
- Mamlouk, K.; Boutiba-Ben Boubaker, I.; Gautier, V.; Vimont, S.; Picard, B.; Ben Redjeb, S.; Arlet, G. Emergence and outbreaks of CTX-M beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae strains in a Tunisian hospital. J. Clin. Microbiol. 2006, 44, 4049–4056. [Google Scholar] [CrossRef]
- Cooper, K.L.; Luey, C.K.; Bird, M.; Terajima, J.; Nair, G.B.; Kam, K.M.; Arakawa, E.; Safa, A.; Cheung, D.T.; Law, C.P.; et al. Development and validation of a PulseNet standardized pulsed-field gel electrophoresis protocol for subtyping of Vibrio cholerae. Foodborne Pathog. Dis. 2006, 3, 51–58. [Google Scholar] [CrossRef]
- Kam, K.M.; Luey, C.K.; Parsons, M.B.; Cooper, K.L.; Nair, G.B.; Alam, M.; Islam, M.A.; Cheung, D.T.; Chu, Y.W.; Ramamurthy, T.; et al. Evaluation and validation of a PulseNet standardized pulsed-field gel electrophoresis protocol for subtyping Vibrio parahaemolyticus: An international multicenter collaborative study. J. Clin. Microbiol. 2008, 46, 2766–2773. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef]
- Deshpande, L.M.; Jones, R.N.; Fritsche, T.R.; Sader, H.S. Occurrence and characterization of carbapenemase-producing Enterobacteriaceae: Report from the SENTRY Antimicrobial Surveillance Program (2000–2004). Microb. Drug Resist. 2006, 2, 223–230. [Google Scholar]
- Moustaoui, N.; Soukri, A.; Elmdaghri, N.; Boudouma, M.; Benbachir, M. Molecular biology of extended-spectrum beta-lactamase-producing Enterobacteriaceae responsible for digestive tract colonization. J. Hosp. Infect. 2004, 57, 202–208. [Google Scholar] [CrossRef]
- Peirano, G.; Hung King Sang, J.; Pitondo-Silva, A.; Laupland, K.B.; Pitout, J.D. Molecular epidemiology of extended-spectrum-β-lactamase-producing Klebsiella pneumoniae over a 10 year period in Calgary, Canada. J. Antimicrob. Chemother. 2012, 67, 1114–1120. [Google Scholar] [CrossRef]
- Hunter, S.B.; Vauterin, P.; Lambert-Fair, M.A.; Duyne, M.S.; Kubota, K.; Graves, L.; Wrigley, D.; Barrett, T.; Ribot, E. Establishment of a universal size standard strain for use with the PulseNet standardized pulsedfield gel electrophoresis protocols: Converting the national databases to the new size standard. J. Clin. Microbiol. 2005, 43, 1045–1050. [Google Scholar] [CrossRef]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar]
- Institut Pasteur. Klebsiella Pneumoniae MLST Database. Available online: www.pasteur.fr/recherche/genopole/PF8/mlst/Kpneumoniae.html (accessed on 2 February 2012).
- Mulvey, M.R.; Chui, L.; Ismail, J.; Louie, L.; Murphy, C.; Chang, N.; Alfa, M.; the Canadian Committee for the Standardization of Molecular Methods. Development of a Canadian standardized protocol for subtyping methicillin-resistant Staphylococcus aureus using pulsed-field gel electrophoresis. J. Clin. Microbiol. 2001, 39, 3481–3485. [Google Scholar] [CrossRef]
- Murchan, S.; Kaufmann, M.E.; Deplano, A.; Ryck, R.; Struelens, M.; Zinn, C.E.; Fussing, V.; Salmenlinna, S.; Vuopio-Varkila, J.; Solh, N.E.; et al. Harmonization of pulsed-field gel electrophoresis protocols for epidemiological typing of strains of methicillin-resistant Staphylococcus aureus: A single approach developed by consensus in 10 European laboratories and its application for tracing the spread of related strains. J. Clin. Microbiol. 2003, 41, 1574–1585. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fitzgerald, C.; Kubota, K.; Swaminathan, B.; Barrett, T.J. rapid pulsed-field gel electrophoresis protocol for subtyping of Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 1889–1894. [Google Scholar] [CrossRef]
- Hillevi, L.; Alex, M.; Dirk, R. Characterization of the genome composition of Bartonella koehlerae by microarray comparative genomic hybridization profiling. J. Bacteriol. 2005, 187, 6155–6165. [Google Scholar] [CrossRef]
- Swaminathan, B.; Barrett, T.J.; Hunter, S.B.; Tauxe , R.V.; the CDC PulseNet Task Force. PulseNet: The molecular subtyping network for foodborne bacterial disease surveillance, United States. Emerg. Infec. Dis. 2001, 7, 382–389. [Google Scholar]
- Struelens, M.J. Molecular epidemiologic typing systems of bacterial pathogens: Current issues and perspectives. Mem. Inst. Oswaldo Cruz 1998, 93, 581–585. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, H.; Zhou, H.; Li, H.; Gao, Y.; Lu, Z.; Hu, K.; Xu, B. Optimization of Pulse-Field Gel Electrophoresis for Subtyping of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2013, 10, 2720-2731. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph10072720
Han H, Zhou H, Li H, Gao Y, Lu Z, Hu K, Xu B. Optimization of Pulse-Field Gel Electrophoresis for Subtyping of Klebsiella pneumoniae. International Journal of Environmental Research and Public Health. 2013; 10(7):2720-2731. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph10072720
Chicago/Turabian StyleHan, Hui, Haijian Zhou, Haishan Li, Yuan Gao, Zhi Lu, Kongxin Hu, and Baoliang Xu. 2013. "Optimization of Pulse-Field Gel Electrophoresis for Subtyping of Klebsiella pneumoniae" International Journal of Environmental Research and Public Health 10, no. 7: 2720-2731. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph10072720