Antimicrobial Susceptibility of Staphylococcus aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa

Abstract
:1. Introduction
2. Materials and Methods
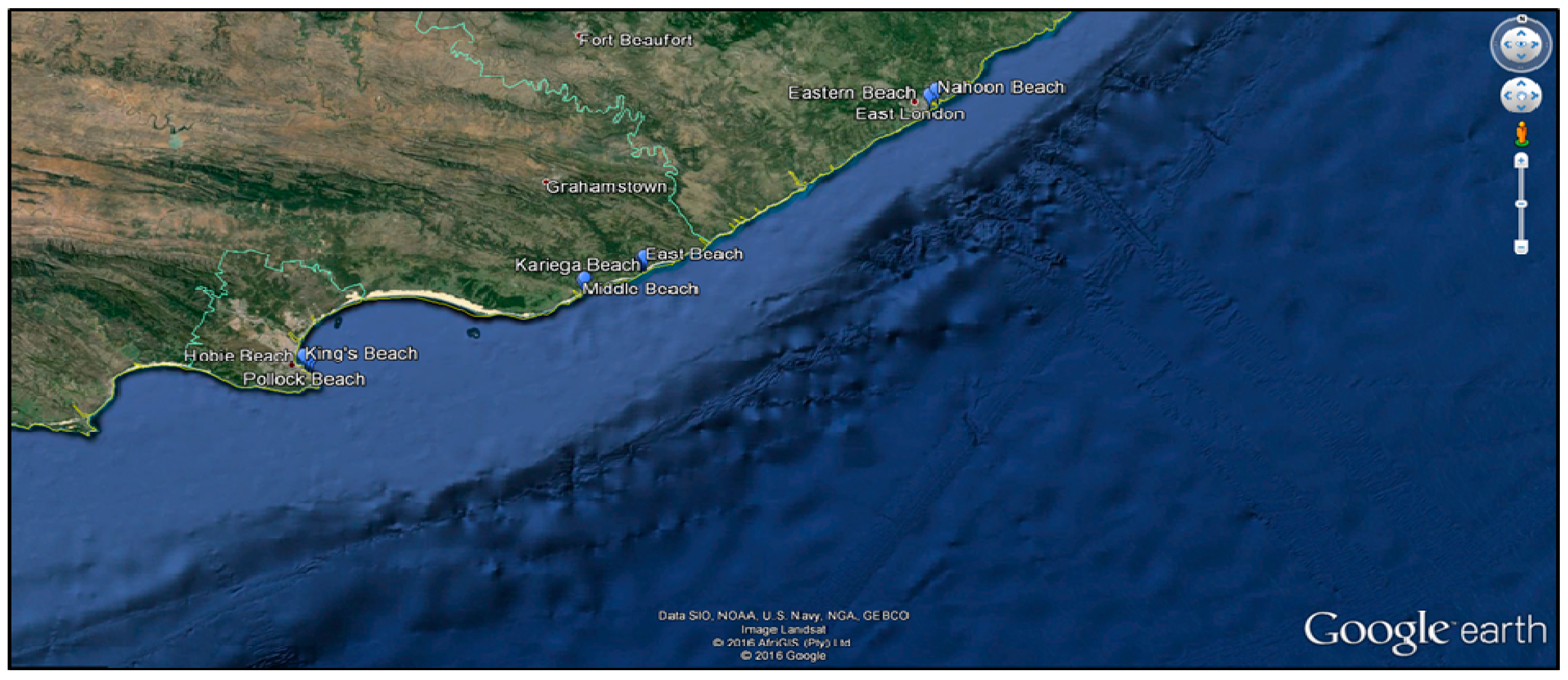
2.1. Study Site
2.2. Sample Collection
Isolation and Molecular Confirmation of S. aureus
2.3. Antimicrobial Susceptibility Testing
2.4. Detection of MRSA
2.4.1. Phenotypic
2.4.2. Molecular Confirmation of MRSA
2.5. PCR Detection of Antibiotic Resistance Genes
3. Results
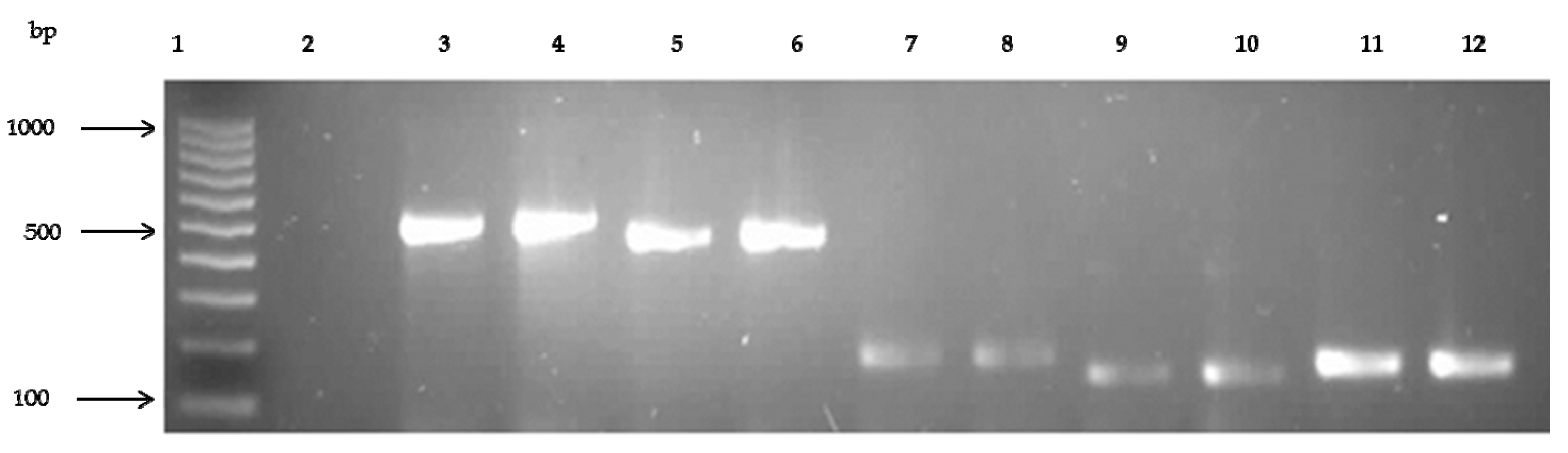
3.1. Molecular Identification of Staphylococcus aureus in Recreational Beach Water and Sand Samples
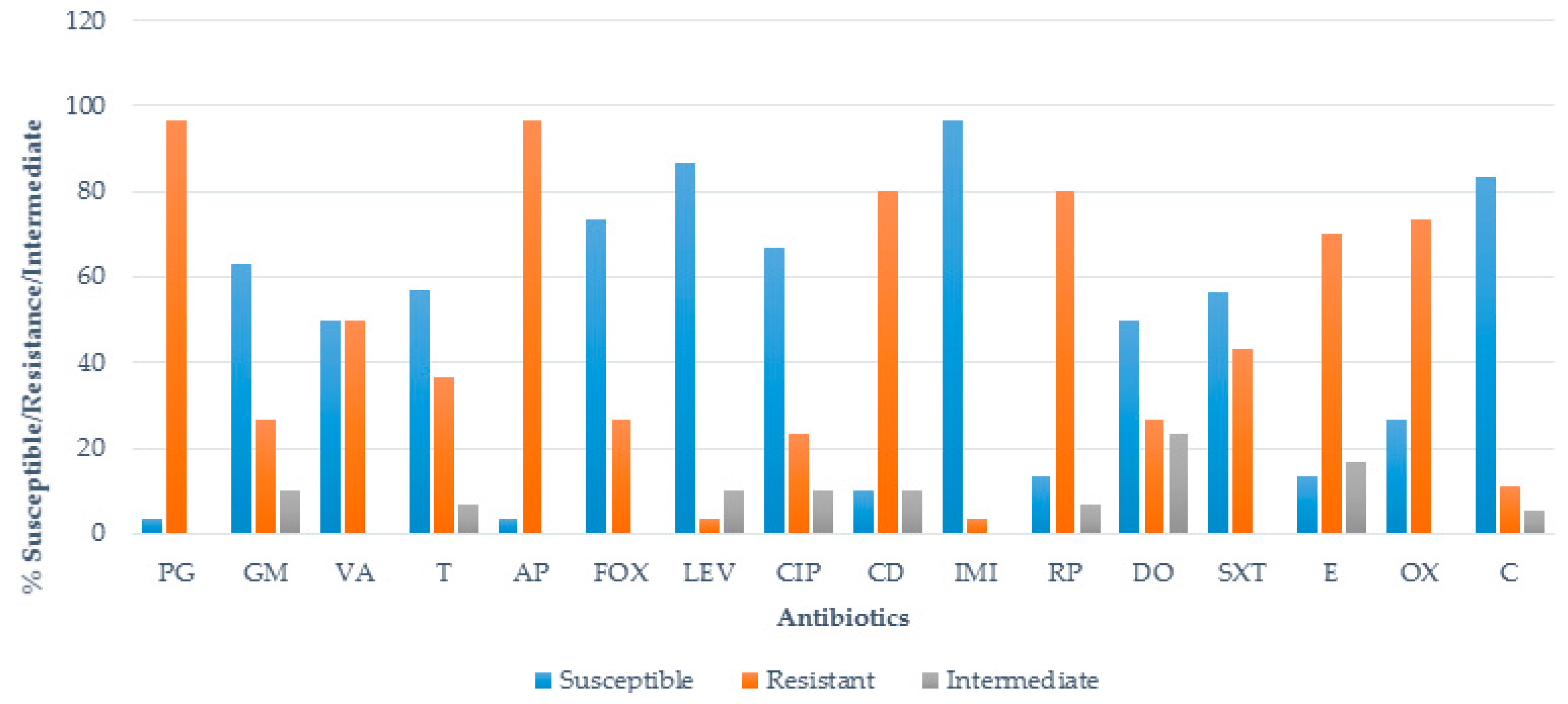
3.2. Antimicrobial Susceptibility Test (AST)
3.3. Phenotypic Detection of MRSA
3.4. Multiple Antibiotic Resistance (MAR)/MAR Phenotypes of S. aureus
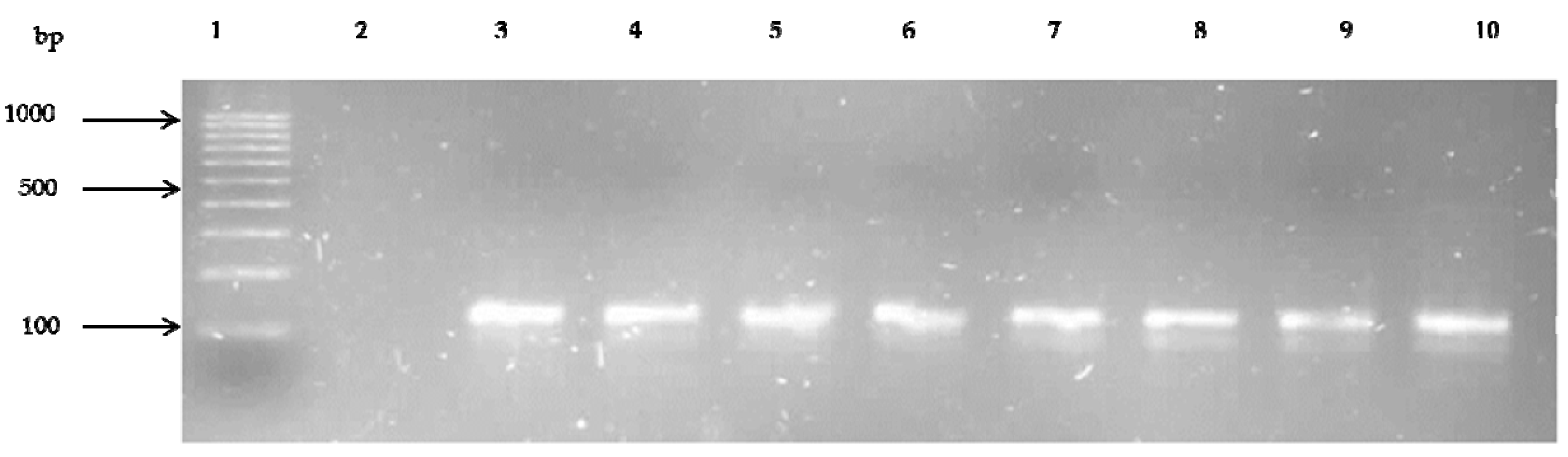
3.5. Prevalence of Antibiotic Resistance Genes
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ho, J.; O’donoghue, M.M.; Boost, M.V. Occupational exposure to raw meat: A newly-recognized risk factor for Staphylococcus aureus nasal colonization amongst food handlers. Int. J. Hyg. Environ. Health 2014, 217, 347–353. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, K.; Lee, J.C. Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev. 2004, 17, 218–234. [Google Scholar] [CrossRef] [PubMed]
- Witte, W.; Strommenger, B.; Klare, I.; Werner, G. Antibiotic-resistant nosocomial pathogens. Part I: Diagnostic and typing methods. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2004, 47, 352–362. [Google Scholar] [PubMed]
- Shuval, H. Estimating the global burden of thalassogenic diseases: Human infectious diseases caused by wastewater pollution of the marine environment. J. Water Health 2003, 1, 53–64. [Google Scholar] [PubMed]
- Yau, V.; Wade, T.J.; de Wilde, C.K.; Colford, J.M., Jr. Skin-related symptoms following exposure to recreational water: A systematic review and meta-analysis. Water Qual. Expos. Health 2009, 1, 79–103. [Google Scholar] [CrossRef]
- Moore, P.C.L.; Lindsay, J.A. Genetic variation among hospital isolates of methicillin-sensitive Staphylococcus aureus: Evidence for horizontal transfer of virulence genes. J. Clin. Microbiol. 2001, 39, 2760–2767. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.F.; Brüssow, H. Common themes among bacteriophage-encoded virulence factors and diversity among the bacteriophages involved. Trends Microbiol. 2002, 10, 521–529. [Google Scholar] [CrossRef]
- Robinton, E.D.; Mood, E.W. A quantitative and qualitative appraisal of microbial pollution of water by swimmers: A preliminary report. J. Hyg. 1966, 64, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Charoenca, N.; Fujioka, R. Association of staphylococcal skin infections and swimming. Water Sci. Technol 1995, 31, 11–17. [Google Scholar]
- WHO. Coastal and Fresh Waters. In Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2003; Volume 1. [Google Scholar]
- Papadakis, J.A.; Mavridou, A.; Richardson, S.C.; Lampiri, M.; Marcelou, U. Bather-related microbial and yeast populations in sand and seawater. Water Res. 1997, 31, 799–804. [Google Scholar] [CrossRef]
- Henrickson, S.E.; Wong, T.; Allen, P.; Ford, T.; Epstein, P.R. Marine swimming-related illness: Implications for monitoring and environmental policy. Environ. Health Perspect. 2001, 109, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Gabutti, G.; De Donno, A.; Bagordo, F.; Montagna, M.T. Comparative survival of faecal and human contaminants and use of Staphylococcus aureus as an effective indicator of human pollution. Mar. Pollut. Bull. 2000, 40, 697–700. [Google Scholar] [CrossRef]
- Calderon, R.L.; Mood, E.W.; Dufour, A.P. Health effects of swimmers and nonpoint sources of contaminated waters. Int. J. Environ. Health Res. 1991, 1, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, P.L.; Tobin, R.S.; Brown, N.E.; Ness, P.F. A prospective study of swimming-related illness. II. Morbidity and the microbiological quality of water. Am. J. Public Health 1985, 75, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Stotts, S.N.; Nigro, O.D.; Fowler, T.L.; Fujioka, R.S.; Steward, G.F. Virulence and antibiotic resistance gene combinations among Staphylococcus aureus isolated from coastal waters of Oahu, Hawaii. J. Young Investig. 2005, 12, 1–8. [Google Scholar]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef] [PubMed]
- Plano, L.R.W.; Garza, A.C.; Shibata, T.; Elmir, S.M.; Kish, J.; Sinigalliano, C.D.; Gidley, M.L.; Miller, G.; Withum, K.; Fleming, L.E.; et al. Shedding of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus from adult and pediatric bathers in marine waters. BMC Microbiol. 2011, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Elmir, S.M.; Wright, M.E.; Abdelzaher, A.; Solo-Gabriele, H.N.; Fleming, L.E.; Miller, G.; Rybolowik, M.; Shih, M.T.; Pillai, P.; Copoer, J.E.; et al. Quantitative evaluation of bacteria released by bathers in marine water. Water Res. 2007, 41, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, K.D.; Pobuda, M. Performance of CHROMagar Staph aureus and CHROMagar MRSA for detection of Staphylococcus aureus in beach water and sand-comparison of culture, agglutination, and molecular analyses. Water Res. 2009, 43, 4802–4811. [Google Scholar] [CrossRef] [PubMed]
- Soge, O.O.; Meschke, J.S.; No, D.B.; Roberts, M.C. Characterization of methicillin-resistant and methicillin-resistant coagulase-negative Staphylococcus spp. isolated from US West Coast public marine beaches. J. Antimicrob. Chemother. 2009, 64, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Tice, A.D.; Susan, J.R. Meeting the challenges of methicillin-resistant Staphylococcus aureus with outpatient parenteral antimicrobial therapy. Clin. Infect. Dis. 2010, 51, S171–S175. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.H.; Abdelzaher, A.M.; Phillips, M.; Hernandez, R.; Solo-Gabriele, H.M.; Kish, J.; Scorzetti, G.; Fell, J.W.; Diaz, M.R.; Scott, T.M.; et al. Indicator microbes correlate with pathogenic bacteria, yeasts and helminthes in sand at a subtropical recreational beach site. J. Appl. Microbiol. 2011, 110, 1571–1583. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, K.D.; Melody, M.; Yiping, C.; Darcy, E.; Melissa, M.; John, F.G. A multi-beach study of Staphylococcus aureus, MRSA, and enterococci in seawater and beach sand. Water Res. 2012, 46, 4195–4207. [Google Scholar] [CrossRef] [PubMed]
- Deurenberg, R.H.; Stobberingh, E.E. The evolution of Staphylococcus aureus. Infect. Genet. Evol. 2008, 8, 747–763. [Google Scholar] [CrossRef] [PubMed]
- Hartman, B.J.; Tomasz, A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J. Bacteriol. 1984, 158, 513–516. [Google Scholar] [PubMed]
- Tavares, A.L. Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in Portugal: Origin, Epidemiology and Virulence. Ph.D. Thesis, Universidade Nova de Lisboa, Lisbon, Portugal, December 2014. [Google Scholar]
- Werckenthin, C.; Cardoso, M.; Martel, J.L.; Schwarz, S. Antimicrobial resistance in staphylococci from animals with particular reference to bovine Staphylococcus aureus, porcine Staphylococcus hyicus and canine Staphylococcus intermedius. Vet. Res. 2001, 32, 341–362. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; McDougal, L.K.; Goering, R.V.; Killgore, G.; Projan, S.J.; Patel, J.B.; Dunman, P.M. Characterization of a strain of community-associated methicillin-resistant Staphylococcus aureus widely disseminated in the United States. J. Clin. Microbiol. 2006, 44, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- GOOGLE MAPS. Map of Eastern Cape Beaches. [Online]. Google, 2016. Available online: https://www.google.co.za/maps/search/eastern+cape+beaches+map+from+east+london+to+port+elizabeth/@33.9988012,22.5881698,2619m/data = !3m2!1e3!4b1?hl = en&authuser = 0 (accessed on 18 June 2016).
- Baums, I.B.; Goodwin, K.D.; Kiesling, T.; Wanless, D.; Fell, J.W. Luminex detection of faecal indicators in river samples, marine recreational water, and beach sand. Mar. Pollut. Bull. 2007, 54, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Soumet, C.; Ermel, G.; Fach, P.; Colin, P. Evaluation of different DNA extraction procedures for the detection of Salmonella from chicken products by polymerase chain reaction. Lett. Appl. Microbiol. 1994, 19, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Maes, N.; Magdalena, J.; Rottiers, S.; De Gheldre, Y.; Struelens, M. Evaluation of a triplex PCR assay to discriminate Staphylococcus aureus from coagulase-negative staphylococci and determine methicillin resistance from blood cultures. J. Clin. Microbiol. 2002, 40, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [PubMed]
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Eighteenth Informational Supplement; CLSI Document M100-S18; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Veenemans, J.; Verhulst, C.; Punselie, R.; Van Keulen, P.H.J.; Kluytmans, J.A.J.W. Evaluation of brilliance MRSA 2 agar for detection of methicillin-resistant Staphylococcus aureus in clinical samples. J. Clin. Microbiol. 2013, 51, 1026–1027. [Google Scholar] [CrossRef] [PubMed]
- Mason, W.J.; Blevins, J.S.; Beenken, K.; Wibowo, N.; Ojha, N.; Smeltzer, M.S. Multiplex PCR protocol for the diagnosis of Staphylococcal infection. J. Clin. Microbiol. 2001, 39, 3332–3338. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for Detection of Genes for Staphylococcus aureus Enterotoxins, Exfoliative Toxins, Toxic Shock Syndrome Toxin 1, and Methicillin Resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [PubMed]
- Schmitz, F.J.; Jones, M.E. Antibiotics for treatment of infections caused by MRSA and elimination of MRSA carriage. What are the choices? Int. J. Antimicrob. Agents 1997, 9, 1–19. [Google Scholar] [CrossRef]
- Martineau, F.; Picard, F.J.; Lansac, N.; Menard, C.; Roy, P.H.; Ouellette, M.; Bergeron, M.G. Correlation between the resistance genotype determined by multiplex PCR assays and the antibiotic susceptibility patterns of Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2000, 44, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Aboshkiwa, M.; Rowland, G.; Coleman, G. Nucleotide sequence of the Staphylococcus aureus RNA polymerase rpoB gene and comparison of its predicted amino acid sequence with those of other bacteria. Biochim. Biophys. Acta BBA Gene Struct. Express. 1995, 1262, 73–78. [Google Scholar] [CrossRef]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Kim, M.J.; Park, C.; Park, J.G.; Lee, G.C. Detection and genotyping of vancomycin-resistant Enterococcus spp. by multiplex polymerase chain reaction in Korean aquatic environmental samples. Int. J. Hyg. Environ. Health 2012, 216, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Kuehnert, M.J.; Hill, H.A.; Kupronis, B.A.; Tokars, J.I.; Solomon, S.L.; Jernigan, D.B. Methicillin-resistant-Staphylococcus aureus hospitalizations, United States. Emerg. Infect. Dis. 2005, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.J.M.; Rafael, C. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.J.C.; de Franco, P.T.R.; Pinto, A.B. Antimicrobial resistance of heterotrophic marine bacteria isolated from seawater and sands of recreational beaches with different organic pollution levels in southeastern Brazil: evidences of resistance dissemination. Environ. Monit. Assess. 2010, 169, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Rajaram, G.; Ramesh, S.; Ashokkumar, S.; Damotharan, P. Occurrence and seasonal distribution of antibiotic resistance heterotrophic bacteria and physico-chemical characteristics of Muthupettai mangrove environment, southeast coast of India. J. Environ. Sci. Technol. 2011, 4, 139–149. [Google Scholar]
- Mudryk, Z.; Perliński, P.; Skórczewski, P. Detection of antibiotic resistant bacteria inhabiting the sand of non-recreational marine beach. Mar. Pollut. Bull. 2010, 60, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Mudryk, Z. Occurrence and distribution antibiotic resistance of heterotrophic bacteria isolated from a marine beach. Mar. Pollut. Bull. 2005, 50, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.W.; Kohenen, W.; Jansen, B.; Obst, U. Detection of antibiotic resistant bacteria and their resistance genes in wastewater, surface water and drinking water biofilms. FEMS Microbiol Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef] [PubMed]
- David, M.Z.; Daum, R.S. Community-Associated Methicillin-Resistant Staphylococcus aureus: Epidemiology and Clinical Consequences of an Emerging Epidemic. Clin. Microbiol. Rev. 2010, 23, 616–687. [Google Scholar] [CrossRef] [PubMed]
- Zetola, N.; Francis, J.S.; Nuermberger, E.L.; Bishai, W.R. Community-acquired meticillin-resistant Staphylococcus aureus: An emerging threat. Lancet Infect. Dis. 2005, 5, 275–286. [Google Scholar] [CrossRef]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Boyle-Vavra, S.; Robert, S.D. Community-acquired methicillin-resistant Staphylococcus aureus: The role of Panton-Valentine leukocidin. Lab. Investig. 2007, 87, 3. [Google Scholar] [CrossRef] [PubMed]
- Abdelzaher, A.M.; Wright, M.E.; Ortega, C.; Solo-Gabriele, H.M.; Miller, G.; Elmir, S.; Newman, X.; Shih, P.; Bonilla, J.A.; Bonilla, T.D.; et al. Presence of pathogens and indicator microbes at a non-point source subtropical recreational marine beach. Appl. Environ. Microbiol. 2010, 76, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Levin-Edens, E.; Bonilla, N.; Meschke, J.S.; Roberts, M.C. Survival of environmental and clinical strains of methicillin-resistant Staphylococcus aureus (MRSA) in marine and fresh waters. Water Res. 2011, 45, 5681–5686. [Google Scholar] [CrossRef] [PubMed]
- Yamahara, K.M.; Sassoubre, L.M.; Goodwin, K.D.; Boehm, A.B. Occurrence and persistence of human pathogens and indicator organisms in beach sands along the California coast. Appl. Environ. Microbiol. 2011, 78, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Gundogan, N.; Citak, S.; Yucel, N.; Devren, A. A note on the incidence and the antibiotic resistance of Staphylococcus aureus isolated from meat and chicken samples. Meat Sci. 2005, 69, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Akindolire, M.A. Detection and Molecular Characterization of Virulence Genes in Antibiotic Resistant Staphylococcus aureus from Milk in the North West Province. Ph.D. Thesis, North West University, Mafikeng, North West Province, South Africa, 2013. [Google Scholar]
- Bartley, J. First case of VRSA identified in Michigan. Infect. Control Hosp. Epidemiol. 2002, 23, 480. [Google Scholar] [PubMed]
- Boerlin, P.; Burnens, A.P.; Frey, J.; Kuhnert, P.; Nicolet, J. Molecular epidemiology and genetic linkage of macrolide and aminoglycoside resistance in Staphylococcus intermedius of canine origin. Vet. Microbiol. 2001, 79, 155–169. [Google Scholar] [CrossRef]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug resistance: An emerging crisis. Interdiscip. Perspect. Infect. Dis. 2014. [Google Scholar] [CrossRef] [PubMed]
- Skórczewski, P.; Jan Mudryk, Z.; Miranowicz, J.; Perlinski, P.; Zdanowicz, M. Antibiotic resistance of Staphylococcus-like organisms isolated from a recreational sea beach on the southern coast of the Baltic Sea as one of the consequences of anthropogenic pressure. Int. J. Oceanogr. Hydrobiol. 2014, 43, 41–48. [Google Scholar] [CrossRef]
- Ateba, C.N.; Mbewe, M.; Moneoang, M.S.; Bezuidenhout, C.C. Antibiotic-resistant Staphylococcus aureus isolated from milk in the Mafikeng Area, North West province, South Africa. S. Afr. J. Sci. 2010, 106, 1–6. [Google Scholar] [CrossRef]
- Pesavento, G.; Ducci, B.; Comodo, N. Antimicrobial resistance profile of Staphylococcus aureus isolated from raw meat: A research for methicillin resistant Staphylococcus aureus (MRSA). Food Control 2007, 18, 196–200. [Google Scholar] [CrossRef]
- Normanno, G.; La Salandra, G.; Dambrosio, A.; Quaglia, N.; Corrente, M.; Parisi, A.; Santagada, G.; Firinu, A.; Crisetti, E.; Celano, G. Occurrence, characterization and antimicrobial resistance of entero-toxigenic Staphylococcus aureus isolated from meat and dairy products. Int. J. Food Microbiol. 2007, 115, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Okurna, K.; Ma, X.X.; Yuzawa, H.; Hiramatsu, K. Insights on antibiotic resistance of Staphylococcus aureus from its whole genome: Genomic island SCC. Drug Resist. Updates 2003, 6, 41–52. [Google Scholar] [CrossRef]
- Guardabassi, L.; Petersen, A.; Olsen, J.E.; Dalsgaard, A. Antibiotic resistance in Acinetobacter sp. isolated from sewers receiving waste effluent from a hospital and a pharmaceutical plant. Appl. Environ. Microbiol. 1998, 64, 3499–3502. [Google Scholar] [PubMed]
- Kümmerer, K. Drugs in the environment: Emission of drugs, diagnostic aids and disinfectants into wastewater by hospitals in relation to other sources—A review. Chemosphere 2001, 45, 957–969. [Google Scholar] [CrossRef]
- Zscheck, K.K.; Murray, B.E. Genes involved in the regulation of β-lactamase production in enterococci and staphylococci, Antimicrob. Agents Chemoth. 1993, 37, 1966–1970. [Google Scholar] [CrossRef]
- Lee, J.H. Methicillin (oxacillin)-resistant Staphylococcus aureus strains isolated from major food animals and their potential transmission to humans. Appl. Environ. Microbiol. 2003, 69, 6489–6494. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.M.; Bayourni, M.A.; Abd El Aal, S.F.A. MRSA detection in raw milk, some dairy products and hands of dairy workers in Egypt, a mini-survey. Food Control 2013, 33, 49–53. [Google Scholar] [CrossRef]
- Mirzaei, H.; Tofighi, A.; Sarabi, H.K.; Faraj1i, M. Prevalence of Methicillin-Resistant Staphylococcus aureus in Raw Milk and Dairy Products in Sarab by Culture and PCR Techniques. J. Anim. Vet. Adv. 2011, 10, 3107–3111. [Google Scholar]
- Börjesson, S.; Melin, S.; Matussek, A.; Lindgren, P.E. A seasonal study of the mecA gene and Staphylococcus aureus including methicillin-resistant S. aureus in a municipal wastewater treatment plant. Water Res. 2009, 43, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Swenson, J.M.; Lonsway, D.; McAllister, S.; Thompson, A.; Jevitt, L.; Zhu, W.; Patel, J.B. Detection of mecA-mediated resistance using reference and commercial testing methods in a collection of Staphylococcus aureus expressing borderline oxacillin MICs. Diagn. Microbiol. Infect. Dis. 2007, 58, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for rapid identification of structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F. Methicillin resistance in staphylococci: Molecular and biochemical basis and clinical implications. Clin. Microbiol. Rev. 1997, 10, 781–791. [Google Scholar] [PubMed]
- Wehrli, W. Rifampin: Mechanisms of action and resistance. Rev. Infect. Dis. 1983, 5, 407–411. [Google Scholar] [CrossRef]
- Aubry-Damon, H.; Soussy, C.J.; Courvalin, P. Characterization of mutants in the rpoB gene that confer rifampin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1998, 42, 2590–2594. [Google Scholar] [PubMed]
- Severinov, K.; Mustaev, A.; Severinova, E.; Kozlov, M.; Darst, S.A.; Goldfarb, A. The b-subunit rif-cluster I is only angstroms away from the active center of Escherichia coli RNA polymerase. J. Biol. Chem. 1995, 270, 29428–29432. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.J.; Gross, C.A. Mapping and sequencing of mutations in the Escherichia coli rpoB gene that lead to rifampicin resistance. J. Mol. Biol. 1988, 202, 45–58. [Google Scholar] [CrossRef]
- Weisblum, B. Erythromycin resistance by ribosomes modification. Antimicrob. Agents Chemother. 1995, 39, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Nicola, F.G.; McDougal, L.K.; Biddle, J.W.; Tenover, F.C. Characterization of erythromycin- resistant isolates of Staphylococcus aureus recovered in the United States from 1958 through 1969. Antimicrob. Agents Chemother. 1998, 42, 3024–3027. [Google Scholar] [PubMed]
- Simeoni, D.; Rizzotti, L.; Cocconcelli, P.; Gazzola, S.; Dellaglio, F.; Torriani, S. Antibiotic resistance genes and identification of staphylococci collected from the production chain of swine meat commodities. Food Microbiol. 2008, 25, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Maidhof, H.; Reinicke, B.; Blumel, P.; Berger-Bachi, B.; Labischinski, H. femA, Which Encodes a Factor Essential for Expression of Methicillin Resistance, Affects Glycine Content of Peptidoglycan in Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Strains. J. Bacterial. 1991, 173, 3507–3513. [Google Scholar] [CrossRef]
- Kobayashi, N.; Wu, H.; Kojima, K.; Taniguchi, K.; Urasawa, S.; Uehara, N.; Omizu, Y.; Kishi, Y.; Yagihashi, A.; Kurokawa, I. Detection of mecA, femA, and femB genes in clinical strains of staphylococci using polymerase chain reaction. Epidemiol. Infect. 1994, 113, 259–266. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Product Size (bp) | Cycling Conditions | Reference |
|---|---|---|---|---|
| nuc-F | GCGATTGATGGTGGATACGGT | 279 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 45 s, 58 °C for 45 s and 72 °C for 90 s. Final extension at 72 °C for 10 min | [35] |
| nuc-R | AGCCAAGCCTTGACGAACTAAAGC | |||
| mecA-F | TCCAGGAATGCAGAAAGACCAAAGC | 499 | Initial denaturation at 94 °C for 3 min, followed by 40 cycles of 94 °C for 30 s, 59 °C for 30 s and 72 °C for 1 min. Final extension at 72 °C for 8 min. | [41] |
| mecA-R | GACACGATAGCCATCTTCATGTTGG | |||
| ermA-F | TATCTTATCGTTGAGAAGGGATT | 139 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 40 s, 48 °C for 40 s and 72 °C for 90 s. Final extension at 72 °C for 8 min. | [42] |
| ermA-R | CTACACTTGGCTTAGGATGAAA | |||
| ermB-F | CTATCTGATTGTTGAAGAAGGATT | 142 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 40 s, 47 °C for 40 s and 72 °C for 90 s. Final extension at 72 °C for 8 min. | [42] |
| ermB-R | GTTTACTCTTGGTTTAGGATGAAA | |||
| ermC-F | CTTGTTGATCACGATAATTTCC | 190 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 40 s, 49 °C for 40 s and 72 °C for 90 s. Final extension at 72 °C for 8 min. | [42] |
| ermC-R | ATCTTTTAGCAAACCCGTATTC | |||
| blaZ-F | ACTTCAACACCTGCTGCTTTC | 173 | Initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 49 °C for 30 s and 72 °C for 1 min. Final extension at 72 °C for 8 min. | [42] |
| blaZ-R | TGACCACTTTTATCAGCAACC | |||
| rpoB1-F | ACCGTCGTTTACGTTCTGTA | 460 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 40 s, 45.5 °C for 40 s and 72 °C for 90 s. Final extension at 72 °C for 8 min. | [43] |
| rpoB2-R | TCAGTGATAGCATGTGTATC | |||
| tetM-F | AGTGGAGCGATTACAGAA | 158 | Initial denaturation at 94 °C for 3 min, followed by 40 cycles of 94 °C for 30 s, 45 °C for 30 s and 72 °C for 1 min. Final extension at 72 °C for 8 min. | [44] |
| tetM-R | CATATGTCCTGGCGTGTCTA | |||
| tetK-F | GTAGCGACAATAGGTAATAGT | 360 | Initial denaturation at 94 °C for 3 min, followed by 40 cycles of 94 °C for 30 s, 47 °C for 30 s and 72 °C for 1 min. Final extension at 72 °C for 8 min. | [44] |
| tetK-R | GTAGTGACAATAAACCTCCTA | |||
| vanA | GCGCGGTCCACTTGTAGATA | 314 | Initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 1 min, 56.5 °C for 1 min and 72 °C for 1 min. Final extension at 72 °C for 10 min. | [45] |
| vanA | TGAGCAACCCCCAAACAGTA | |||
| vanB | AGACATTCCGGTCGAGGAAC | 220 | Initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 1 min, 56.5 °C for 1 min and 72 °C for 1 min. Final extension at 72 °C for 10 min. | [45] |
| vanB | GCTGTCAATTAGTGCGGGAA | |||
| femA-F | AAAAAAGCACATAACAAGCG | 132 | Initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 40 s, 45.5 °C for 40 s and 72 °C for 90 s. Final extension at 72 °C for 8 min. | [40] |
| femA-R | GATAAAGAAGAAACCAGCAG |
| No. Resistant by Disc Diffusion | Associated ARG Tested | ARG Detected | ||
|---|---|---|---|---|
| Sand (%) | Water (%) | Total (%) | ||
| Ampicillin & Penicillin (n = 29) | blaZ | 4 (25%) | 12 (75%) | 16 (55.2%) |
| Methicillin (n = 22) | mecA | 1 (20%) | 4 (80%) | 5 (22.7%) |
| Rifampicin (n = 24) | rpoB | 2 (18.2%) | 9 (81.8%) | 11 (45.8%) |
| Erythromycin (n = 21) | ermB | 3 (20%) | 12 (80%) | 15 (71.4%) |
| Tetracycline (n = 11) | tetM | 1 (12.5%) | 7 (87.5%) | 8 (72.7%) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akanbi, O.E.; Njom, H.A.; Fri, J.; Otigbu, A.C.; Clarke, A.M. Antimicrobial Susceptibility of Staphylococcus aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2017, 14, 1001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph14091001
Akanbi OE, Njom HA, Fri J, Otigbu AC, Clarke AM. Antimicrobial Susceptibility of Staphylococcus aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa. International Journal of Environmental Research and Public Health. 2017; 14(9):1001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph14091001
Chicago/Turabian StyleAkanbi, Olufemi Emmanuel, Henry Akum Njom, Justine Fri, Anthony C. Otigbu, and Anna M. Clarke. 2017. "Antimicrobial Susceptibility of Staphylococcus aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa" International Journal of Environmental Research and Public Health 14, no. 9: 1001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph14091001